Correlation between vein density and water use efficiency of Salix matsudana in Zhangye Wetland, China
XUTing, ZHAOCheng-Zhang*,, HANLing, FENGWei, DUANBei-Bei, ZHENGHui-Ling College of Geography and Environmental Science, Northwest Normal University, Research Center of Wetland Resources Protection and Industrial Development Engineering of Gansu Province, Lanzhou 730070, China 收稿日期:2016-05-31 接受日期:2017-04-6 网络出版日期:2017-07-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家自然科学基金(41461013和91125014) 作者简介: -->* 通信作者Author for correspondence (E-mail:zhaocz601@163.com)
关键词:旱柳;叶脉密度;水分利用效率;关联性;土壤水分梯度;张掖湿地 Abstract Aims The correlation between vein density and water use efficiency (WUE) affects the balance between water supply and demand of plant leaves, which is significant for comprehending the ecological adaptation strategies of plants. The objective of this study was to study how Salix matsudana modulated vein density and WUE along a soil moisture gradient in Zhangye Wetland, China. Methods The study was conducted in floodplain wetland near Heihe River in Zhangye City, Gansu Province, China. Three sample plots, at a spatial interval of 70 m, were set up along a soil moisture gradient ordinally from the area near the water body to the wetland edge, plot I (69.23%), spot II (48.38%) and spot III (35.27%). Community traits were investigated by using diagonal method, and all individuals of S. matsudana were used for measurements of height and canopy. At each plot, 5 individuals of S. matsudana at 4 vertices and diagonal intersection were selected for measurements of vein density, WUE, net photosynthetic rate (Pn), transpiration rate (Tr), photosynthetically active radiation (PAR), saturated vapor pressure differences (VPD), specific leaf area, stomatal conductance (Gs) and intercellular CO2 concentration (Ci). We used mathematical methods of correlation analysis and standardized major axis to investigate relationships between vein density and WUE. Important findings With decreasing soil moisture, the height, canopy, specific leaf area, Gs and Ci of S. matsudana decreased gradually, while the vein density, WUE, Pn, Tr, PAR and VPD increased gradually. The correlation between vein density and WUE was positive in all the three plots, but the relationship varied along the soil moisture plots gradient. There was a highly significant positive correlation (p < 0.01) between the vein density and WUE at plot I and III, whereas the correlation only reached a significant level (p < 0.05) at plot II; The correlation coefficient between vein density and WUE is significantly smaller than 1 at plot I (p < 0.05), while the correlation coefficient is significant greater than 1 at plot II and III (p < 0.05). We can conclude that varied relationships between vein density and WUE of S. matsudana along a soil moisture gradient could reflect plant acclimation.
不同样地旱柳林群落的平均高度、郁闭度均存在显著差异(p < 0.05, 表1)。随着样地I向样地II、III的逐渐过渡, 土壤含水量减少, 旱柳林样地群落的平均高度、郁闭度均呈现逐渐减小趋势, 平均土壤含水量减少49.05%, 群落平均高度减少21.13 %, 郁闭度降低了59.15%。表明在3个样地中, 样地I生境条件(土壤含水量65%-75%)更适宜旱柳植株的生长。 Table 1 表1 表1不同水分样地的主要特征(平均值±标准误差, n = 60) Table 1Main characteristics of plots in different water plots (mean ± SE, n = 60)
样地 Plot
土壤含水量 Soil moisture (%)
群落特征 Community characteristics
高度 Height (cm)
郁闭度 Crown density (%)
I
69.23 ± 3.55a
636.96 ± 27.63a
91.21 ± 4.56a
II
48.38 ± 2.46b
589.25 ± 17.28b
75.69 ± 3.78b
III
35.27 ± 1.76c
502.34 ± 26.76c
37.26 ± 1.86c
Different lowercase letters in the same column indicate significant differences among plots (p < 0.05).同列不同小写字母表示样地间差异显著(p < 0.05)。 新窗口打开
2.2 不同样地旱柳叶性状特征及WUE
不同土壤含水量条件下旱柳种群叶性状特征及WUE的变化如图1所示。由单因素方差分析得出, 旱柳单叶面积、比叶面积、叶脉密度及WUE在不同水分梯度存在显著差异(p < 0.05)。随着土壤含水量的减少, 旱柳单叶面积和比叶面积均呈逐渐减小趋势, 从样地I到样地III分别减少了71.67%和29.14%; 叶脉密度和WUE则均呈增大趋势, 从样地I到样地III分别增加了43.00%和54.10%。 显示原图|下载原图ZIP|生成PPT 图1不同样地旱柳叶性状特征及水分利用效率(WUE)的变化(平均值±标准误差, n = 60)。不同小写字母表示相同构件在不同样地差异显著(p < 0.05)。 -->Fig. 1Change among leaf traits and water use efficiency (WUE) of Salix matsudana in different plots (mean ± SE, n = 60). Different lowercase letters indicate significant differences of different plots for identical module (p < 0.05). -->
2.3 不同样地旱柳叶片光合特性
不同土壤含水量条件下旱柳叶片光合特性的变化如表2所示。由单因素方差分析得出, 旱柳气孔导度(Gs)、Pn、Tr、光合有效辐射(PAR)和VPD在不同水分梯度存在显著差异(p < 0.05), 而胞间CO2浓度(Ci)在不同梯度差异不显著(p > 0.05)。随着土壤含水量的减少, 旱柳Gs和Ci呈减小趋势, 从样地I到样地III分别减少了81.92%和12.90%; 旱柳Pn、Tr、PAR和VPD均呈增大趋势, 从样地I到样地III分别增加了77.81%、55.65%、49.92%及21.88%。 Table 2 表2 表2不同样地旱柳叶片光合特性(数值为平均值±标准误差, n = 60) Table 2Photosynthetic characteristics of Salix matsudana in different plots (mean ± SE, n = 60)
样地 Plot
I
II
III
饱和水汽压差 VPD (kPa)
0.25 ± 0.01c
0.28 ± 0.01b
0.32 ± 0.02a
气孔导度 Gs (mol·m-2·s-1)
1.77 ± 0.09a
0.91 ± 0.05b
0.32 ± 0.02c
蒸腾速率 Tr (mmol·m-2·s-1)
3.02 ± 0.15c
5.26 ± 0.26b
6.81 ± 0.34a
净光合速率 Pn (μmol·m-2·s-1)
3.69 ± 0.08c
8.83 ± 0.34b
16.63 ± 0.83a
胞间CO2浓度 Ci (μmol·mol-1)
0.31 ± 0.02a
0.29 ± 0.02a
0.27 ± 0.01a
光合有效辐射 PAR (μmol·m-2·s-1)
782.30 ± 39.12c
1105.20 ± 55.26b
1562.20 ± 78.12a
Ci, intercellular CO2 concentration; Gs, stomatal conductance; PAR, photosynthetically active radiation; Pn, net photosynthetic rate; Tr, transpiration rate; VPD, vapor pressure deficit. Different lowercase letters in the same column indicate significant differences among plots (p < 0.05).同列不同小写字母表示样地间差异显著(p < 0.05)。 新窗口打开
2.4 不同样地旱柳叶脉密度与WUE的关系分析
如图2所示, 样地I和样地III旱柳叶脉密度与WUE间均存在极显著正相关(p < 0.01), 而样地II存在显著正相关关系(p < 0.05, 图2)。在水分梯度上随着旱柳叶脉密度的增加, WUE也呈增加趋势, 表明旱柳叶脉密度和WUE间存在一定协调性。不同水分梯度旱柳叶脉密度与WUE间的相关系数存在显著差异(p < 0.05), 样地I叶脉密度与WUE的相关系数显著小于1, 样地II和样地III的相关系数显著大于1 (p < 0.05), 表明样地I旱柳叶片WUE的增加速度小于叶脉密度的增加速度, 而样地II和样地III旱柳叶片WUE的增加速度要显著大于叶脉密度的增加速度。 显示原图|下载原图ZIP|生成PPT 图2不同样地旱柳叶脉密度与水分利用效率(WUE)的关系。A, 样地I; B, 样地II; C, 样地III。样地同表1。 -->Fig. 2Relationship between vein density and water use efficiency (WUE) of Salix matsudana among different plots. A, Plot I; B, Plot II; C, Plot III. Plot see Table 1. -->
AibaM, TakafumiH, HiuraT (2012). Interspecific differences in determinants of plant species distribution and the relationships with functional traits. Journal of Ecology, 100, 950-957. [本文引用: 1]
[2]
BerlynGP, MikscheJP (1976). Botanical Microtechnique and Cytochemistry. Iowa State University Press, Ames, USA. [本文引用: 1]
[3]
BlonderB, ViolleC, BentleyLP, EnquistBJ (2010). Venation networks and the origin of the leaf economics spectrum. Ecology Letters, 14, 91-100. [本文引用: 1]
[4]
BrodribbTJ, FeildTS (2010). Leaf hydraulic evolution led a surge in leaf photosynthetic capacity during early angiosperm diversification. Ecology Letters, 13, 175-183. [本文引用: 1]
[5]
BrodribbTJ, JordanGJ (2011). Water supply and demand remain balanced during leaf acclimation of Nothofagus cunninghamii trees. New Phytologist, 192, 437-448. [本文引用: 1]
[6]
Carins-MurphyMR, JordanGJ, BrodribbTJ (2014). Acclimation to humidity modifies the link between leaf size and the density of veins and stomata. Plant, Cell & Environment, 37, 124-131. [本文引用: 1]
[7]
DangJJ, ZhaoCZ, DongXG, YangQ, ZhaGD (2014). Response of Kobresia tibetica and Kobresia humilis population spatial pattern to soil moisture. Chinese Journal of Ecology, 33, 1734-1740. (in Chinese with English abstract)[党晶晶, 赵成章, 董小刚, 杨泉, 查高德 (2014). 藏嵩草和矮嵩草种群空间分布格局对水分的响应 . 生态学杂志, 33, 1734-1740.] [本文引用: 1]
EscheraP, PeukeaAD, BannisterbP, FinkcS, HartungdW, JiangeF, RennenbergH (2008). Transpiration, CO2 assimilation, WUE, and stomatal aperture in leaves of Viscum album (L.): Effect of abscisic acid (ABA) in the xylem sap of its host (Populus × euamericana). Plant Physiology and Biochemistry, 46, 64-70. [本文引用: 1]
[10]
FlexasJ, Díaz-EspejoA, ConesaMA, CoopmanRE, DoutheC, GagoJ, GalléA, GalmésJ, MedranoH, Ribas-CarboM, TomàsM, NiinemetsÜ (2015). Mesophyll conductance to CO2 and Rubisco as targets for improving intrinsic water use efficiency in C3 plants. Plant, Cell & Environment, 39, 965-982. [本文引用: 1]
[11]
GangCC, WangZQ, ChenYZ, YangY, LiJL, ChengJM, QiJG, OdeheI (2016). Drought-induced dynamics of carbon and water use efficiency of global grasslands from 2000 to 2011. Ecological Indicators, 67, 788-797. [本文引用: 1]
[12]
GaoL, YangJ, LiuRX (2009). Effects of soil moisture levels on photosynthesis, transpiration, and moisture use efficiency of female and male plants of Hippophae rhamnoidesssp ssp. sinensi. Acta Ecologica Sinica, 29, 6025-6034. (in Chinese with English abstract)[高丽, 杨劼, 刘瑞香 (2009). 不同土壤水分条件下中国沙棘雌雄株光合作用、蒸腾作用及水分利用效率特征 . 生态学报, 29, 6025-6034.] [本文引用: 1]
[13]
GaoY, XiaJB, ZhaoZG, KongQX, GuGY, ZhangL (2013). Effect of simulated soil water change on photosynthetic characteristics of Salix matsudana in shell sand habitat. Acta Botanica Boreali-Occidentalia Sinica, 33, 2467-2473. (in Chinese with English abstract)[高源, 夏江宝, 赵自国, 孔庆仙, 谷广义, 张亮 (2013). 模拟贝壳砂水分变化对旱柳光合特性的影响 . 西北植物学报, 33, 2467-2473.] [本文引用: 1]
[14]
GaoY, ZhuXJ, YuGR, HeNP, WangQF, TianJ (2014). Water use efficiency threshold for terrestrial ecosystem carbon sequestration in China under afforestation. Agricultural and Forest Meteorology, 195-196, 32-37. [本文引用: 1]
[15]
GlennDM (2016). Effect of highly processed calcined kaolin residues on apple water use efficiency. Scientia Horticulturae, 23, 127-132. [本文引用: 1]
[16]
GongR, GaoQ (2015). Research progress in the effects of leaf hydraulic characteristics on plant physiological functions. Chinese Journal of Plant Ecology, 39, 300-308. (in English with Chinese abstract)[龚容, 高琼 (2015). 叶片结构的水力学特性对植物生理功能影响的研究进展 . 植物生态学报, 39, 300-308.] [本文引用: 2]
[17]
GurevitchJ, ScheinerSM, FoxGA (2002). The Ecology of Plants . Sinauer Associates, Sunderland, USA. [本文引用: 1]
[18]
HeCX, LiJY, MengP, ZhangJS (2013). Changes of leaf traits and WUE with crown height of four tall tree species. Acta Ecologica Sinica, 33, 5644-5654. (in English with Chinese abstract)[何春霞, 李吉跃, 孟平, 张劲松 (2013). 4种高大树木的叶片性状及WUE随树高的变化 . 生态学报, 33, 5644-5654.] [本文引用: 3]
[19]
KenneyAM, McKayJK, RichardsJH, JuengerTE (2014). Direct and indirect selection on flowering time, water-use efficiency (WUE, δ13C), and WUE plasticity to drought in Arabidopsis thaliana. Ecology and Evolution, 4, 4505-4521. [本文引用: 1]
[20]
LiMX, YangYZ, ZhuQ, ChenH, PengCH (2016). Evaluating water use efficiency patterns of Qinling Mountains under climate change. Acta Ecologica Sinica, 36, 936-945. (in Chinese with English abstract)[李明旭, 杨延征, 朱求安, 陈槐, 彭长辉 (2016). 气候变化背景下秦岭地区陆地生态系统水分利用率变化趋势 . 生态学报, 36, 936-945.] [本文引用: 1]
[21]
LiX, XieYH, HuangJS, ChenXS (2009). Research progresses on the formation mechanism of vegetation distribution pattern in wetlands. Wetland Science, 7, 280-288. (in Chinese with English abstract)[李旭, 谢永宏, 黄继山, 陈心胜 (2009). 湿地植被格局成因研究进展 . 湿地科学, 7, 280-288.] [本文引用: 1]
[22]
LiuXJ, MaKP (2015). Plant functional traits—Concepts, applications and future directions. Scientia Sinica Vitae, 45, 325-339. (in Chinese with English abstract)[刘晓娟, 马克平 (2015). 植物功能性状研究进展 . 中国科学: 生命科学, 45, 325-339.] [本文引用: 1]
[23]
Matthew-OgburnR, EdwardsEJ (2013). Repeated origin of three-dimensional leaf venation releases constraints on the evolution of succulence in plants. Current Biology, 23, 722-726. [本文引用: 1]
[24]
MengTT, NiJ, WangGH (2007). Plant functional traits, environments and ecosystem functioning. Journal of Plant Ecology ( Chinese Version), 31, 150-165. (in Chinese with English abstract)[孟婷婷, 倪健, 王国宏 (2007). 植物功能性状与环境和生态系统功能 . 植物生态学报, 31, 150-165.] [本文引用: 1]
[25]
NardiniA, LuglioJ (2014). Leaf hydraulic capacity and drought vulnerability: Possible trade-offs and correlations with climate across three major biomes. Functional Ecology, 28, 810-818. [本文引用: 1]
[26]
NardiniA, RaimondoF, LoGulloMA (2010). Leaf miners help us understand leaf hydraulic design . Plant, Cell and Environment, 33, 1091-1100. [本文引用: 1]
[27]
NiuYY, LiaoK, JiaY, PangHX, XuGX, JiangZB (2015). Analysis on differences in photosynthetic characteristics of korla fragrant pear among different planting densities. Agricultural Science & Technology, 16, 862-867. [本文引用: 1]
[28]
PaganoM, StorchiP (2015). Leaf vein density: A possible role as cooling system. Journal of Life Sciences, 9, 299-303. [本文引用: 1]
[29]
PradhanS, SehgalVK, SanooRN, BandyopadhyayKK, SinghR (2014). Yield, water, radiation and nitrogen use efficiencies of wheat (Triticum aestivum) as influenced by nitrogen levels in a semi-arid environment. Indian Journal of Agronomy 59, 267-275. [本文引用: 1]
[30]
QuanXK, WangCK (2015). Comparison of foliar water use efficiency among 17 provenances of Larix gmelinii in the Mao’ershan area. Chinese Journal of Plant Ecology, 39, 352-361. (in English with Chinese abstract)[全先奎, 王传宽 (2015). 帽儿山17个种源落叶松针叶的水分利用效率比较 . 植物生态学报, 39, 352-361.] [本文引用: 1]
[31]
RenQJ, LiHL, BuHY (2015). Comparison of physiological and leaf morphological traits for photosynthesis of the 51 plant species in the Maqu alpine swamp meadow. Chinese Journal of Plant Ecology, 39, 593-603.[任青吉, 李宏林, 卜海燕 (2015). 玛曲高寒沼泽化草甸51种植物光合生理和叶片形态特征的比较 . 植物生态学报, 39, 593-603.] [本文引用: 1]
[32]
SackL, FroleK (2006). Leaf structural diversity is related to hydraulic capacity in tropical rain forest trees. Ecology, 87, 483-491. [本文引用: 1]
[33]
SackL, ScoffoniC (2013). Leaf venation: Structure, function, development, evolution, ecology and applications in the past, present and future. New Phytologist, 198, 983-1000. [本文引用: 2]
[34]
SackL, ScoffoniC, JohnGP, PoorterH, MasonCM, Mendez-AlonzoR, DonovanLA (2013). How do leaf veins influence the worldwide leaf economic spectrum? Review and synthesis. Journal of Experimental Botany, 64, 4053-4080. [本文引用: 1]
[35]
SackL, ScoffoniC, McKownAD, FroleK, RawlsM, HavranJC, TranH, TranT (2012). Developmentally based scaling of leaf venation architecture explains global ecological patterns. Nature Communications, 3, 837. [本文引用: 2]
[36]
SilvertownJ, CharlesworthD (2001). Introduction to Plant Population Biology . Blackwell, London. [本文引用: 1]
[37]
SongLQ, HuCM, HouXL, ShiL, LiuLA, YangJC, JiangCD (2015). Relationship between photosynthetic characteristics and leaf vein density in Sorghum bicolor and Perilla frutescens. Chinese Bulletin of Botany, 50, 100-106. (in English with Chinese abstract)[宋丽清, 胡春梅, 侯喜林, 石雷, 刘立安, 杨景成, 姜闯道 (2015). 高粱、紫苏叶脉密度与光合特性的关系 . 植物学报, 50, 100-106.] [本文引用: 1]
[38]
SongYC (2001). Vegetation Ecology. East China Normal University Press, Shanghai. 551-552. (in Chinese)[宋永昌 (2001). 植被生态学. 华东师范大学出版社, 上海. 551-552.] [本文引用: 1]
[39]
TanXJ, ZhaoXS (2006). Spatial distribution and ecological adaptability of wetland vegetation in Yellow River Delta along a water table depth gradient. Chinese Journal of Ecology, 25, 1460-1464. (in Chinese with English abstract)[谭学界, 赵欣胜 (2006). 水深梯度下湿地植被空间分布与生态适应 . 生态学杂志, 25, 1460-1464.] [本文引用: 1]
[40]
VillagraM, CampanelloPI, BucciSJ, GoldsteinG (2013). Functional relationships between leaf hydraulics and leaf economic traits in response to nutrient addition in subtropical tree species. Tree Physiology, 33, 1308-1318. [本文引用: 1]
[41]
ViolleC, NavasML, VileD, KazakouE, FortunelC, HummelI, GarnierE (2007). Let the concept of trait be functional! Oikos, 116, 882-892. [本文引用: 1]
[42]
WangJL, YuYR, FangQX, JiangDF, QiH, WangQF (2008). Responses of water use efficiency of nine plant species to light and CO2 and it’s modeling. Acta Ecologica Sinica, 28, 525-533. (in Chinese with English abstract)[王建林, 于贵瑞, 房全孝, 姜德锋, 齐华, 王秋凤 (2008). 不同植物叶片水分利用效率对光和CO2的响应与模拟 . 生态学报, 28, 525-533.] [本文引用: 1]
[43]
WangLL, WangL, ZhangLF, LiuYY, XuSJ (2015). Structure and dynamic characteristics of Gymnocarpos przewalskii in different habitats. Chinese Journal of Plant Ecology, 39, 980-989. (in Chinese with English abstract)[王立龙, 王亮, 张丽芳, 刘玉洋, 徐世健 (2015). 浙江天童木本植物叶片性状空间变异的地统计学分析 . 植物生态学报, 39, 980-989.] [本文引用: 1]
[44]
WangYP, GaoHH, LiuYS, MuP, YuXP, AnLZ, ZhangF (2013). Adaptation mechanisms of alpine plants photosynthetic apparatus against adverse stress: A review. Chinese Journal of Applied Ecology, 24, 2049-2055. (in Chinese with English abstract)[王玉萍, 高会会, 刘悦善, 慕平, 鱼小军, 安黎哲, 张峰 (2013). 高山植物光合机构耐受胁迫的适应机制 . 应用生态学报, 24, 2049-2055.] [本文引用: 1]
[45]
WestobyM, FalsterDS, MolesAT (2002). Plant ecological strategies: Some leading dimensions of variation between species. Annual Review of Ecology Systematics, 33, 125-159. [本文引用: 1]
[46]
WuHW, LiXY, JingZY, LiJ, ZhengXR, ZhaoDZ (2015). Variations in water use for Achnatherum splendens in Lake Qinghai watershed based on δD and δ18O. Acta Ecologica Sinica, 35, 8174-8183. (in Chinese with English abstract)[吴华武, 李小雁, 蒋志云, 李静, 郑肖然, 赵殿智 (2015). 基于δD和δ18O的青海湖流域芨芨草水分利用来源变化研究 . 生态学报, 35, 8174-8183.] [本文引用: 1]
[47]
XuMS, ZhaoYT, YangXD, ShiQR, ZhouLL, ZhangQQ, ArshadA, YanER (2016). Geostatistical analysis of spatial variations in leaf traits of woody plants in Tiantong, Zhejiang Province. Chinese Journal of Plant Ecology, 40, 48-59. (in Chinese with English abstract)[许洺山, 赵延涛, 杨晓东, 史青茹, 周刘丽, 张晴晴, AliARSHAD, 阎恩荣 (2016). 浙江天童木本植物叶片性状空间变异的地统计学分析 . 植物生态学报, 40, 48-59.] [本文引用: 2]
[48]
YangWD, ChenYT (2008). Membrane leakage and antioxidant enzyme activities in roots and leaves of Salix matsudana with cadmium stress. Acta Botanica Boreali-Occidentalia Sinica, 28, 2263-2269. (in Chinese with English abstract)[杨卫东, 陈益泰 (2008). 镉胁迫对旱柳细胞膜透性和抗氧化酶活性的影响 . 西北植物学报, 28, 2263-2269.] [本文引用: 1]
[49]
YuWY, JiRP, FengR, ZhaoXL, ZhangYS (2015). Response of water stress on photosynthetic characteristics and water use efficiency of maize leaves in different growth stage. Acta Ecologica Sinica, 35, 2902-2909. (in Chinese with English abstract)[于文颖, 纪瑞鹏, 冯锐, 赵先丽, 张玉书 (2015). 不同生育期玉米叶片光合特性及水分利用效率对水分胁迫的响应 . 生态学报, 35, 2902-2909.] [本文引用: 1]
[50]
ZhangYQ, LiSW, FuW, WenHD (2014). Effects of nitrogen application on yield, photosynthetic characteristics and water use efficiency of hybrid millet. Journal of Plant Nutrition and Fertilizer, 20, 1119-1126. (in Chinese with English abstract)[张亚琦, 李淑文, 付巍, 文宏达 (2014). 施氮对杂交谷子产量与光合特性及水分利用效率的影响 . 植物营养与肥料学报, 20, 1119-1126.] [本文引用: 1]
Interspecific differences in determinants of plant species distribution and the relationships with functional traits. 1 2012
Transpiration, CO2 assimilation, WUE, and stomatal aperture in leaves of Viscum album (L.): Effect of abscisic acid (ABA) in the xylem sap of its host (Populus × euamericana). 1 2008
... 旱柳(Salix matsudana)是杨柳科柳属的落叶乔木, 具有根系发达、抗风能力强、生长快、易繁殖、耐寒、耐旱及较好的防风固沙和保持水土功能等优点, 广泛分布于河滩、湖泊堤岸、渠道及道路边, 已成为中国北方道路防护林、沙荒及盐碱湿地造林的主要树种(高源等, 2013), 对改善生态环境起着重要作用(杨卫东和陈益泰, 2008).近年来, 学术界对生态系统WUE对气候变化的响应(李明旭等, 2016), WUE与光合速率、蒸腾速率(Escher et al., 2008; Glenn, 2016)、CO2浓度(王建林等, 2008; Flexas et al., 2015)等的关系, 不同生育期(于文颖等, 2015)、土壤水分(高丽等, 2009; Gang et al., 2016)、氮素含量(Pradhan et al., 2014; 张亚琦等, 2014)等条件下植物WUE的变化规律, 树高对WUE的影响(何春霞等, 2013), WUE与叶脉密度与叶大小(Sack & Scoffoin, 2013)、光合特性(Brodribb & Field, 2010; 宋丽清等, 2015)的关系及对环境因子的响应(Blonder et al., 2010; Matthew-Ogburn & Edwards, 2013)等方面有较多研究, 解释了植物适应特定环境的性状塑造, 而对不同水分条件下植物叶脉密度与WUE关联性的机理研究尚不深入.鉴于此, 本研究以黑河中游张掖段泛洪平原湿地的天然旱柳林为研究对象, 通过研究旱柳在水分梯度上叶脉密度与WUE的关系, 试图明晰: (1)不同水分条件下旱柳叶脉密度与WUE间存在何种关系?(2)导致这种关系存在的主要原因有哪些?旨在丰富对湿地植物叶性状建成的种群适应性策略的认识和理解. ...
Mesophyll conductance to CO2 and Rubisco as targets for improving intrinsic water use efficiency in C3 plants. 1 2015
... 旱柳(Salix matsudana)是杨柳科柳属的落叶乔木, 具有根系发达、抗风能力强、生长快、易繁殖、耐寒、耐旱及较好的防风固沙和保持水土功能等优点, 广泛分布于河滩、湖泊堤岸、渠道及道路边, 已成为中国北方道路防护林、沙荒及盐碱湿地造林的主要树种(高源等, 2013), 对改善生态环境起着重要作用(杨卫东和陈益泰, 2008).近年来, 学术界对生态系统WUE对气候变化的响应(李明旭等, 2016), WUE与光合速率、蒸腾速率(Escher et al., 2008; Glenn, 2016)、CO2浓度(王建林等, 2008; Flexas et al., 2015)等的关系, 不同生育期(于文颖等, 2015)、土壤水分(高丽等, 2009; Gang et al., 2016)、氮素含量(Pradhan et al., 2014; 张亚琦等, 2014)等条件下植物WUE的变化规律, 树高对WUE的影响(何春霞等, 2013), WUE与叶脉密度与叶大小(Sack & Scoffoin, 2013)、光合特性(Brodribb & Field, 2010; 宋丽清等, 2015)的关系及对环境因子的响应(Blonder et al., 2010; Matthew-Ogburn & Edwards, 2013)等方面有较多研究, 解释了植物适应特定环境的性状塑造, 而对不同水分条件下植物叶脉密度与WUE关联性的机理研究尚不深入.鉴于此, 本研究以黑河中游张掖段泛洪平原湿地的天然旱柳林为研究对象, 通过研究旱柳在水分梯度上叶脉密度与WUE的关系, 试图明晰: (1)不同水分条件下旱柳叶脉密度与WUE间存在何种关系?(2)导致这种关系存在的主要原因有哪些?旨在丰富对湿地植物叶性状建成的种群适应性策略的认识和理解. ...
Drought-induced dynamics of carbon and water use efficiency of global grasslands from 2000 to 2011. 1 2016
... 旱柳(Salix matsudana)是杨柳科柳属的落叶乔木, 具有根系发达、抗风能力强、生长快、易繁殖、耐寒、耐旱及较好的防风固沙和保持水土功能等优点, 广泛分布于河滩、湖泊堤岸、渠道及道路边, 已成为中国北方道路防护林、沙荒及盐碱湿地造林的主要树种(高源等, 2013), 对改善生态环境起着重要作用(杨卫东和陈益泰, 2008).近年来, 学术界对生态系统WUE对气候变化的响应(李明旭等, 2016), WUE与光合速率、蒸腾速率(Escher et al., 2008; Glenn, 2016)、CO2浓度(王建林等, 2008; Flexas et al., 2015)等的关系, 不同生育期(于文颖等, 2015)、土壤水分(高丽等, 2009; Gang et al., 2016)、氮素含量(Pradhan et al., 2014; 张亚琦等, 2014)等条件下植物WUE的变化规律, 树高对WUE的影响(何春霞等, 2013), WUE与叶脉密度与叶大小(Sack & Scoffoin, 2013)、光合特性(Brodribb & Field, 2010; 宋丽清等, 2015)的关系及对环境因子的响应(Blonder et al., 2010; Matthew-Ogburn & Edwards, 2013)等方面有较多研究, 解释了植物适应特定环境的性状塑造, 而对不同水分条件下植物叶脉密度与WUE关联性的机理研究尚不深入.鉴于此, 本研究以黑河中游张掖段泛洪平原湿地的天然旱柳林为研究对象, 通过研究旱柳在水分梯度上叶脉密度与WUE的关系, 试图明晰: (1)不同水分条件下旱柳叶脉密度与WUE间存在何种关系?(2)导致这种关系存在的主要原因有哪些?旨在丰富对湿地植物叶性状建成的种群适应性策略的认识和理解. ...
不同土壤水分条件下中国沙棘雌雄株光合作用、蒸腾作用及水分利用效率特征 1 2009
... 旱柳(Salix matsudana)是杨柳科柳属的落叶乔木, 具有根系发达、抗风能力强、生长快、易繁殖、耐寒、耐旱及较好的防风固沙和保持水土功能等优点, 广泛分布于河滩、湖泊堤岸、渠道及道路边, 已成为中国北方道路防护林、沙荒及盐碱湿地造林的主要树种(高源等, 2013), 对改善生态环境起着重要作用(杨卫东和陈益泰, 2008).近年来, 学术界对生态系统WUE对气候变化的响应(李明旭等, 2016), WUE与光合速率、蒸腾速率(Escher et al., 2008; Glenn, 2016)、CO2浓度(王建林等, 2008; Flexas et al., 2015)等的关系, 不同生育期(于文颖等, 2015)、土壤水分(高丽等, 2009; Gang et al., 2016)、氮素含量(Pradhan et al., 2014; 张亚琦等, 2014)等条件下植物WUE的变化规律, 树高对WUE的影响(何春霞等, 2013), WUE与叶脉密度与叶大小(Sack & Scoffoin, 2013)、光合特性(Brodribb & Field, 2010; 宋丽清等, 2015)的关系及对环境因子的响应(Blonder et al., 2010; Matthew-Ogburn & Edwards, 2013)等方面有较多研究, 解释了植物适应特定环境的性状塑造, 而对不同水分条件下植物叶脉密度与WUE关联性的机理研究尚不深入.鉴于此, 本研究以黑河中游张掖段泛洪平原湿地的天然旱柳林为研究对象, 通过研究旱柳在水分梯度上叶脉密度与WUE的关系, 试图明晰: (1)不同水分条件下旱柳叶脉密度与WUE间存在何种关系?(2)导致这种关系存在的主要原因有哪些?旨在丰富对湿地植物叶性状建成的种群适应性策略的认识和理解. ...
模拟贝壳砂水分变化对旱柳光合特性的影响 1 2013
... 旱柳(Salix matsudana)是杨柳科柳属的落叶乔木, 具有根系发达、抗风能力强、生长快、易繁殖、耐寒、耐旱及较好的防风固沙和保持水土功能等优点, 广泛分布于河滩、湖泊堤岸、渠道及道路边, 已成为中国北方道路防护林、沙荒及盐碱湿地造林的主要树种(高源等, 2013), 对改善生态环境起着重要作用(杨卫东和陈益泰, 2008).近年来, 学术界对生态系统WUE对气候变化的响应(李明旭等, 2016), WUE与光合速率、蒸腾速率(Escher et al., 2008; Glenn, 2016)、CO2浓度(王建林等, 2008; Flexas et al., 2015)等的关系, 不同生育期(于文颖等, 2015)、土壤水分(高丽等, 2009; Gang et al., 2016)、氮素含量(Pradhan et al., 2014; 张亚琦等, 2014)等条件下植物WUE的变化规律, 树高对WUE的影响(何春霞等, 2013), WUE与叶脉密度与叶大小(Sack & Scoffoin, 2013)、光合特性(Brodribb & Field, 2010; 宋丽清等, 2015)的关系及对环境因子的响应(Blonder et al., 2010; Matthew-Ogburn & Edwards, 2013)等方面有较多研究, 解释了植物适应特定环境的性状塑造, 而对不同水分条件下植物叶脉密度与WUE关联性的机理研究尚不深入.鉴于此, 本研究以黑河中游张掖段泛洪平原湿地的天然旱柳林为研究对象, 通过研究旱柳在水分梯度上叶脉密度与WUE的关系, 试图明晰: (1)不同水分条件下旱柳叶脉密度与WUE间存在何种关系?(2)导致这种关系存在的主要原因有哪些?旨在丰富对湿地植物叶性状建成的种群适应性策略的认识和理解. ...
Water use efficiency threshold for terrestrial ecosystem carbon sequestration in China under afforestation. 1 2014
 , 韩玲, 冯威, 段贝贝, 郑慧玲
, 韩玲, 冯威, 段贝贝, 郑慧玲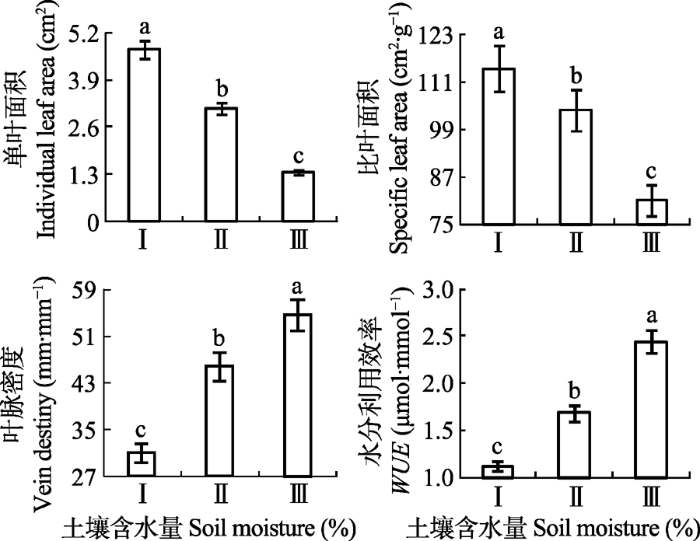 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}