0 引言
【研究意义】小麦是最重要的粮食作物之一,其稳定发展对保障中国的粮食安全具有重要意义。近十年来,中国小麦连年丰收,除了栽培技术不断改进外,新品种选育与其大面积推广在增产中也发挥了重要作用。从20世纪50年代到目前为止,主产麦区共经历了8—9次品种更新换代,不同时期的小麦新品种为中国小麦单产和总产的提高做出了重要贡献[1]。因此,从不同角度对具有代表性小麦新品种的遗传构成进行分析,挖掘其内在遗传规律,对于新品种的育种应用和亲本选配具有重要的指导意义。【前人研究进展】近年来,前人对中国不同时期骨干亲本的遗传机制进行了较多研究,结果表明,骨干亲本中含有与产量特性和抗病性相关的染色体区段,并且这些区段更容易被育种家所选择,而携带重要片段的后代往往性状优良最终成为不同时期的代表性品种[2,3,4,5,6,7,8,9]。目前,关于正在大面积推广品种或有较大应用潜力新品种遗传特性的研究相对较少。李小军等[10]分析了小麦品种百农AK58的遗传构成,发现其更多地继承了亲本周麦11的遗传物质,且其多数特异染色体位点与已知重要农艺性状相关。李俊等[11]对小麦新品种川麦104的遗传构成进行了分析,认为亲本川麦42对其遗传贡献更大,且来源于双亲的14个与产量相关染色体片段是形成川麦104高产的遗传基础。邹少奎等[12]分析了小麦新品种周麦23的遗传构成,发现其63.31%的遗传物质来源于母本周麦13,并且筛选到一个周麦23的特异引物,可用来鉴定周麦23的真实性。利用相同的方法,邹少奎等[13]还对小麦新品种周麦22的遗传基础进行了研究,发现亲本温麦6号对其遗传贡献最大。【本研究切入点】对目前正在推广小麦新品种的遗传构成进行分析,可有助于育种家了解当前推广品种的遗传规律,进而有目的地指导品种改良。淮麦33由江苏徐淮地区淮阴农业科学研究所以烟农19为母本、郑麦991为父本杂交,采用系谱法经7代系统选育而成。在2012年国家区试和2013年国家生产试验中均居参试品种第一位,较对照品种周麦18增产极显著,2014年通过国家审定(国审麦2014001)[14]。在生产上,淮麦33表现出高产、稳产、适应性好、抗倒性强及抗逆性突出等优点,已在黄淮南片麦区大面积种植。淮麦33优良性状的遗传力较高,目前,以淮麦33为亲本还育成优异品系淮麦4046,正在参加2017—2018年度国家生产试验。淮麦33的母本烟农19产量潜力高且品质优良,多年来一直是江苏、安徽淮北麦区主导品种,但抗倒性差的缺点限制了其进一步推广应用,这一直是育种家对其改良的重点。父本郑麦991的分蘖性强,茎秆较矮且抗倒性好,已广泛为育种家所利用。同两亲本相比,淮麦33优良性状的构成如何?双亲对这些优良性状又有哪些遗传贡献?这些问题都尚未得到解决。【拟解决的关键问题】本研究以淮麦33及其双亲为材料,通过农艺性状表型鉴定、高分子量麦谷蛋白亚基组成及SSR标记分析等多个角度,解析其遗传构成特征,从而为小麦品种改良及亲本选配提供依据。1 材料与方法
1.1 材料
供试材料为淮麦33及其双亲烟农19和郑麦991,均由江苏徐淮地区淮阴农业科学研究所小麦研究室提供。1.2 农艺及品质性状分析
供试材料于2014—2016年连续3年种植于淮安市农科院科研创新基地,各年度按照随机区组设计,3次重复,每小区种植8行,行长7.5 m,行距20 cm。条沟点播,出苗后定苗,确保各小区基本苗数一致,定苗后进行常规田间管理。田间调查每平方米穗数。成熟期取样考种,测量株高和每穗粒数。收获整个小区测定小区产量,取样测千粒重,并用波通DA7200多功能近红外分析仪(瑞士产)测定总蛋白质和湿面筋含量。利用SPSS v19.0软件进行显著性分析和多重比较。
1.3 高分子量谷麦蛋白亚基组成分析
高分子量谷蛋白亚基的提取参照张玲丽等[15]试验方法稍加改进进行,并利用SDS-PAGE电泳分析。根据PAYNE等[16]方法为高分子量麦谷蛋白亚基命名。对照品种为中国春(null、7+8和2+12)。1.4 分子标记分析
分子标记分析主要在南京农业大学作物遗传与种质创新国家重点实验室进行。选取每个品种的幼嫩叶片,用CTAB法提取基因组DNA。参考XUE等[17]研究,选用625个覆盖小麦全基因组的SSR标记(包括Xgwm、Xwmc、Xbarc、Xgdm、Xcfa和Xcfd系列)进行PCR扩增。PCR反应在Mastercycler nexus X1 PCR仪(Eppendorf,德国)上进行,PCR体系为10×PCR Buffer(Mg2+ Plus) 1.25 µL、dNTP Mix(各2.5 mmol·L-1)1.0 µL、10 µmol·L-1的上下游引物各0.5 µL、模板DNA(30—50 ng·µL-1)1 µL和Taq DNA聚合酶(5 U·µL-1)0.0625 µL。PCR程序为94℃ 3 min;94℃ 30 s,60℃ 40 s,72℃ 40 s,10个循环(touch-down,每循环降低1.0℃);94℃ 30 s,50℃ 40 s,72℃ 40 s,25个循环;72℃ 10 min。
扩增产物采用6%聚丙烯酰胺凝胶电泳检测。分析电泳结果,进行带型统计,在相同迁移率处,有带记为1,无带记为0,形成0、1数据库。依据标记多态性结果,计算双亲对后代的遗传贡献率,某一亲本对后代的遗传贡献率为后代中同该亲本相同的特异位点数与所有亲本特异位点总数的百分比。利用NTSYSpc2.10e软件计算遗传相似系数。结合SOMER等[18]的遗传连锁图和GrainGenes2.0(http://wheat.pw. usda.gov/)的相关信息,估计SSR位点间的遗传距离,利用GGT2.0软件绘制品种基因型图谱。
2 结果
2.1 淮麦33及其亲本部分农艺性状和品质指标比较
比较各试验年度淮麦33与双亲在5个农艺性状及2个品质性状间的差异性(表1)。其中,同烟农19相比,淮麦33的株高有了显著降低。分析其产量构成因素,淮麦33的每平方米穗数和千粒重平均值均介于两亲本之间,其中,千粒重与两亲本差异不显著;每平方米穗数显著低于郑麦991,但与烟农19未达显著水平;穗粒数在3个年度都显著高于两亲本。淮麦33连续3年小区产量显著高于双亲,平均较烟农19增产7.7%,较郑麦991增产12.1%。另外,对于所测定的2个品质性状,在蛋白质含量上,淮麦33平均值为13.6%,其在2014—2016年度蛋白质的平均含量高于郑麦991,而低于烟农19,但均未达到显著水平;淮麦33的湿面筋含量年度平均值均显著低于烟农19,而高于郑麦991。Table 1
表1
表1淮麦33与其亲本部分农艺性状及品质性状差异性比较
Table 1Statistical analysis of the agronomical and quality traits evaluated between Huaimai33 and its parents
| 年份 Year | 品种 Cultivar | 株高 Plant height (cm) | 每平方米穗数 Spike number per square meter | 每穗粒数 Grain number per spike | 千粒重 1000-grain weight (g) | 产量 Grain yield (kg·hm-2) | 蛋白质含量 Protein content (%) | 湿面筋含量 Wet gluten content (%) |
|---|---|---|---|---|---|---|---|---|
| 2014—2015 | 淮麦33 Huaimai33 | 80.6b | 490.0b | 47.8b | 45.7a | 9326.5b | 13.5ab | 31.0b |
| 烟农19 Yannong19 | 83.2c | 476.3b | 43.5a | 46.5a | 8762.1a | 14.1b | 32.4b | |
| 郑麦991 Zhengmai991 | 70.8a | 526.7a | 39.8a | 43.8a | 8313.7a | 12.2a | 28.9a | |
| 2015—2016 | 淮麦33 Huaimai33 | 78.5b | 482.5b | 45.3c | 43.0a | 8882.3c | 13.2b | 30.5b |
| 烟农19 Yannong19 | 80.8b | 452.0b | 40.2b | 43.5a | 8136.5b | 14.5c | 33.0c | |
| 郑麦991 Zhengmai991 | 70.3a | 511.2a | 35.3a | 41.6a | 7856.6a | 12.0a | 28.5a | |
| 2016—2017 | 淮麦33 Huaimai33 | 81.1b | 518.5b | 46.9b | 43.2b | 9545.0b | 14.2b | 32.0ab |
| 烟农19 Yannong19 | 85.4c | 475.2c | 42.4a | 44.2b | 8876.4a | 15.1c | 33.8b | |
| 郑麦991 Zhengmai991 | 74.2a | 568.4a | 38.2a | 40.5a | 8589.8a | 12.9a | 29.5a | |
| 平均 Mean | 淮麦33 Huaimai33 | 80.1b | 497.0b | 46.7c | 44.0a | 9251.3b | 13.6ab | 31.2b |
| 烟农19 Yannong19 | 83.1c | 467.8b | 42.0b | 44.7a | 8591.7a | 14.6b | 33.1c | |
| 郑麦991 Zhengmai991 | 71.8a | 535.4a | 37.8a | 42.0a | 8253.4 a | 12.4a | 29.0a |
新窗口打开
2.2 淮麦33及其亲本高分子量麦谷蛋白亚基组成分析
SDS-PAGE分析结果表明(图1),淮麦33及其亲本的高分子量麦谷蛋白亚基组成各不相同,其中淮麦33为(1、17+18和2+12),烟农19为(1、17+18和5+10),郑麦991为(null、7+9和2+12),表明淮麦33在Glu-A1位点的1亚基和Glu-B1位点的17+18亚基均来自于母本烟农19,而在Glu-D1的2+12亚基则由父本郑麦991所贡献。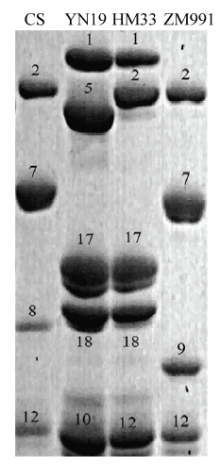 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1淮麦33及亲本高分子量麦谷蛋白亚基组成的SDS- PAGE分析
CS:中国春;YN19:烟农19;HM33:淮麦33;ZM991:郑麦991。图中数字为相应的高分子量谷蛋白亚基名称
-->Fig. 1SDS-PAGE analysis for HMW-GS composition in Huaimai33 and its patents
CS: Chinese Spring; YN19: Yannong19; HM33: Huaimai33; ZM991: Zhengmai991. The number represents the name of corresponding HMW-GS
-->
2.3 淮麦33的遗传构成分析
在625个SSR标记中,其中475个能扩增出清晰的带型,涉及全基因组水平614个位点(个别标记存在2个或2个以上位点)。其中,在250个位点上,淮麦33与两亲本带型一致;在38个位点上与两亲本均不一致,未能确定等位基因来源;剩余的326个位点,241个位点来源于烟农19,85个位点来源于郑麦991。双亲烟农19和郑麦991对淮麦33的遗传贡献率分别为73.9%和26.1%,说明两亲本遗传物质在后代中发生了较大的偏分离,且淮麦33更多地继承了烟农19的遗传物质。对淮麦33和两亲本的遗传相似度进行分析,发现淮麦33与烟农19的遗传相似系数为0.78,与郑麦991的遗传相似系数为0.49,表明淮麦33与母本烟农19的亲缘关系更近。图2为部分SSR标记在供试品种中的扩增结果。其中,Xgwm192和Xwmc52在3个供试材料中没有多态性,Xwmc236和Xwmc169在淮麦33中的扩增结果与烟农19一致,Xgdm125和Xgwm169在淮麦33中的扩增结果与郑麦991一致,Xwmc488和Xgwm156为淮麦33特异标记。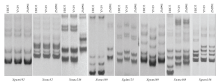 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2部分SSR引物电泳图谱
HM33:淮麦33;YN19:烟农19;ZM991:郑麦991
-->Fig. 2Electrophoresis pattern for partial SSR markers
HM33: Huaimai33; YN19: Yannong19; ZM991: Zhengmai991
-->
分析在基因组和染色体水平上两亲本对淮麦33的遗传贡献率(图3)。在基因组水平上,两亲本对淮麦33的遗传贡献率存在一定的不均衡性,其中烟农19在A、B和D 3个基因组水平上的遗传贡献率均较高,分别为75.1%、69.4%和68.7%;而郑麦991的遗传贡献率分别为24.9%、30.6%和31.3%。在染色体水平上,两亲本对淮麦33的遗传贡献率差异较大。在6A染色体上,烟农19的遗传贡献率为25%,而郑麦991为75%,除此之外,烟农19在其他染色体的遗传贡献率均高于郑麦991,其中在2A染色体上遗传贡献率达到100%,在1A、3A、2B、3B和4B等6条染色体上的遗传贡献率均超过90%。
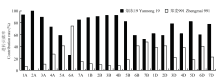 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3亲本对淮麦33在不同染色体的遗传贡献率
-->Fig. 3Percentages of the parental contributions to different chromosomes of Huaimai33
-->
2.4 淮麦33来源于双亲的染色体区段
利用GGT2.0软件绘制了淮麦33的基因型图谱(图4)。在21条染色体上,遗传距离大于5 cM的染色体区段中,淮麦33来源于双亲的有41个,共包括163个位点,其中有34个来源于烟农19,7个来源于郑麦991。来源于烟农19的区段在2D和1A染色体上较多,分别为4个和3个;但来源郑麦991的区段在5A染色体上最多,为2个。在A、B和D 3个基因组中,以A基因组来源于烟农19的区段最多,为13个,其中除6A(未发现来源于烟农19的区段)和1A染色体(3个)外,其余5条染色体均为2个;B基因组中来源于烟农19的区段有9个,D基因组中有12个。来源于郑麦991的7个染色体区段中,A和D基因组各有3个,B基因组有1个。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4淮麦33的21条染色体SSR基因型图谱
-->Fig. 4Genotypic maps of 21 chromosomes from Huaimai33 based on SSR markers
 烟农19位点 Allele from Yannong19;
烟农19位点 Allele from Yannong19; 郑麦991位点 Allele from Zhengmai991;
郑麦991位点 Allele from Zhengmai991; 烟农19和郑麦991相同位点 Common allele of Yannong 19 and Zhengmai991;
烟农19和郑麦991相同位点 Common allele of Yannong 19 and Zhengmai991; 淮麦33特异位点 Huaimai33-specific allele
淮麦33特异位点 Huaimai33-specific allele-->
2.5 淮麦33的特异染色体位点
比较淮麦33与2个亲本SSR多态性,发现31个SSR标记与2亲本带型均不相同,共涉及38个位点,分布于1B、1D、2A、2B、2D、3A、3B、3D、4A、4B、5A、5B、6B、6D和7A等15条染色体上(图4)。淮麦33在2A染色体上的特异位点最多,为9个;其次为3B、3D、4A、4B和5A,各有3个。淮麦33在2A染色体上形成特异QTL区段Xgwm339—Xgwm55,共包括4个位点,标记间遗传距离为2 cM;在4A和6D染色体上形成特异区段Xbarc170—Xgwm637和Xgdm98—Xwmc773,标记间遗传距离分别为10和11 cM。3 讨论
本研究对淮麦33及其亲本主要农艺性状的差异性分析结果表明,淮麦33的每平方米穗数和千粒重均介于两亲本之间,处于适中且相对较高的水平,而每穗粒数表现出明显的超亲优势。在株高方面,相比烟农19,淮麦33的株高有了显著降低,抗倒伏能力显著增强。可见,协调的产量三因素以及生产上所表现出的抗倒伏、抗寒、耐旱和抗病等优点共同形成了淮麦33高产稳产的基础。在品质方面,淮麦33在Glu-B1位点含有17+18优质亚基,且为非1B/1R易位系(待发表),这为淮麦33的优异品质做出了积极贡献,其蛋白质和湿面筋含量均较高。据黄淮南片区域试验品质总结,淮麦33在2011—2012及2012—2013两年度平均蛋白质含量(干基)高达14.78%,平均湿面筋含量达到33%。在Glu-D1位点含有2+12亚基,而LÜ等[19]的研究表明小麦5+10亚基对面粉品质贡献优于2+12亚基,这也可能是导致淮麦33综合品质指标未能达到烟农19优质强筋标准的主要因素。前人对小麦产量相关QTL进行了大量研究,发现其在小麦整个染色体组上均有分布[20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47]。对比已报道的小麦产量QTL位点信息,淮麦33来源于烟农19的241个位点中,有39个与产量性状相关,占比16.2%,主要分布在1A、1D、2A、2B、2D、3A、3B、3D、4A、4B、4D、5A、5B、6A、6B、7A和7D等17条染色体上(表2)。一些产量相关SSR位点在染色体上成簇分布,如在1A染色体上由Xwmc120—Xgwm164—Xwmc333构成标记间遗传距离为7 cM的染色体区段(图4),被发现与小麦株高相关[20,22];在2D染色体上的Xgwm261—Xgwm296区段标记间遗传距离为1 cM,富集了与小麦产量[21,28]、粒重[26,29]和穗粒数[21]相关的QTL;3B染色体上的Xbarc173—Xwmc505—Xbarc68—Xbarc73区段标记间遗传距离为6 cM,与控制粒重[31]和产量[30,31]的基因相关联;4B染色体上的Xgwm107—Xwmc48—Xgwm495区段与粒重[22,32]、株高[32,34]、穗粒数[22,26,35-36]和有效穗数[35]等性状相关;4D染色体上Xwmc617—Xwmc48区段分布着与穗粒数[29]、粒重[25]、株高[25,29]相关的QTL;7A染色体上的Xgwm282—Xgwm332区段控制着有效穗数[21]、穗粒数[32]和千粒重[32]等性状。淮麦33来源于郑麦991的85个位点中,与产量相关的有17个,占比20%,主要分布在1A、1B、1D、2D、3D、4A、4B、5A、6B、6D和7B等11条染色体上(表2)。其中,位于1D染色体上的标记Xcfd72和Xbarc169相距7 cM,两位点分别与产量[37]和粒重[34]QTL相关联;4A染色体上的Xwmc232—Xbarc78区段定位了与株高[20]和有效穗数[41]相关的QTL;5A染色体上的Xbarc56—Xbarc186区段存在控制粒重[30]、有效穗数[22]和产量[30]的QTL;5A染色体上的Xwmc410—Xwmc524区段存在控制株高(Rht12)[27]和产量[29]的基因。有趣的是,淮麦33中来源于不同亲本与产量相关的染色体区段分别存在于不同的染色体上,表明淮麦33聚合了来自于2个亲本的产量相关区段,这些染色体区段可能是构成其高产的分子基础。但是,淮麦33中的产量相关区段是否对产量有正向效应还需要更深入的研究,为此,笔者正在构建烟农19和郑麦991的重组自交系群体,希望通过QTL作图的方法评价这些产量相关区段的遗传效应。另外,通过比对淮麦33及其双亲中矮杆基因Rht-B1、Rht-D1和Rht8的基因型差异,发现淮麦33中除了Rht8基因型与烟农19一致外,另外2个基因位点的基因型在双亲间均没有差异,表明上述已知矮源不是淮麦33株高下降的原因(待发表)。鉴于淮麦33株高降低有限(3—4 cm)且偏向于高秆亲本烟农19,导致淮麦33株高降低的主要因素可能来自于一些微效株高QTL的重组,这些微效株高QTL是否来自于矮杆亲本郑麦991有待更深入研究。
Table 2
表2
表2淮麦33产量相关SSR位点
Table 2Yield-related SSR loci in Huaimai33
| 来源 Parental source | SSR位点 SSR loci | 性状 Trait | SSR位点 SSR loci | 性状 Trait | SSR位点 SSR loci | 性状 Trait | ||
|---|---|---|---|---|---|---|---|---|
| YN19 | Xgwm164-1A | PH[20] | Xwmc120-1A | GN[20], PH[20] | Xwmc333-1A | GW[21], PH[22] | ||
| Xcfa2129-1D | PH[23] | Xgwm232-1D | GY[21] | Xgwm372-2A | GY[24] | |||
| Xgwm257-2B | GY[25] | Xwmc317-2B | SN[26], PH[27] | Xgwm261-2D | GY[21,28], GW[26], GN[21] | |||
| Xgwm296-2D | GY[28], GW[29], GN[21] | Xgwm539-2D | SN[24] | Xwmc18-2D | GN[26], PH[25] | |||
| Xgwm391-3A | GW[26], SN[26] | Xbarc173-3B | GW[30] | Xbarc68-3B | GY[31] | |||
| Xbarc73-3B | GY[30] | Xwmc505-3B | GW[30] | Xgdm72-3D | GY[32] | |||
| Xgwm161-3D | GN[28, 32] | Xgwm397-4A | GY[33] | Xgwm494-4A | GW[25] | |||
| Xwmc48-4A | PH[34] | Xwmc48-4B | PH[34] | Xgwm495-4B | GW[22], GN[22, 26, 35-36], SN[35] | |||
| Xgwm6-4B | PH[34] | Xgwm107-4B | GW[32], PH[32] | Xwmc48-4D | GN[29], PH[29] | |||
| Xwmc617-4D | GW[25], PH[25] | Xgwm410-5A | SN[26] | Xbarc151-5A | GY[37], SN[30] | |||
| Xgwm604-5B | GY[38], GW[38] | Xgwm639-5B | GW[38] | Xbarc232-5B | PH[25] | |||
| Xgwm334-6A | GW[39] | Xwmc494-6B | GW[26] | Xgwm332-7A | SN[21] | |||
| Xwmc83-7A | GY[29], GW[26], SN[26] | Xgwm282-7A | GW[32], GN[32] | Xgwm44-7D | GW [24,40] | |||
| ZM991 | Xwmc93-1A | SN42] | Xgwm259-1B | GN[26] | Xbarc169-1D | GW[34] | ||
| Xcfd72-1D | GY[37] | Xgwm349-2D | GW[30] | Xgwm484-2D | GN[20] | |||
| Xwmc492-3D | PH[20] | Xbarc78-4A | SN[41] | Xwmc232-4A | PH[20] | |||
| Xgwm149-4B | GW[22,43], GN[22, 26], PH[22, 44] | Xbarc186-5A | GW[30] | Xbarc56-5A | SN[22], GY[30], GW[30] | |||
| Xwmc524-5A | GY[29] | Xwmc410-5A | PH[27] | Xwmc397-6B | GY[45] | |||
| Xbarc196-6D | GW[23] | Xgwm400-7B | GW[20] | |||||
| Specific loci | Xgwm636-2A | GW[46] | Xbarc124-2A | GW[23] | Xgwm95-2A | GY[32] | ||
| Xgwm114-3B | SN[28] | Xgwm493-3B | PH[47] | Xgwm513-4B | GW[21], SN[35], PH[25] | |||
| Xgwm156-5A | GY[30, 32], PH[32] | Xbarc117-5A | GW[41] | Xwmc773-6D | GY[22] | |||
| Xgdm98-6D | GW[34], GY[22] | Xwmc488-7A | GW[26], GN[26] |
新窗口打开
本研究中,淮麦33的特异位点比例较高,达到了10.4%,高于百农AK58(4.9%)[10]和周麦23(5.0%)[12],而低于周麦22(16.4%)[13],这些特异位点可能来自于杂交后代选育过程中双亲遗传物质的基因重组、自然突变或者小概率发生的异花授粉等。进一步的分析发现,在38个淮麦33特异位点中,有11个与产量性状相关,比例为28.9%,主要分布在2A、3B、4B、5A、6D和7A等6条染色体上(表2)。其中位于2A染色体上的Xbarc124和Xgwm636标记相距3 cM,两位点均定位了与粒重相关的QTL[23,46];位于6D染色体上的Xgdm98—Xwmc773区段标记间遗传距离为11 cM,存在着控制产量[22]和粒重[34]的基因。另外,特异位点Xgwm95-2A[32]、Xgwm156-5A[30,32]、Xwmc773-6D[22]和Xgdm98-6D[22]直接与小区产量有关,Xwmc488-7A[26]与穗粒数相关,这些位点可能是导致淮麦33产量和穗粒数超亲表现的主要因素,值得进一步研究。
理论上,双亲对单交品种的遗传贡献率均应为50%,但在实际的育种实践中,常常出现子代遗传物质偏向于某一亲本的现象,李玉刚等[9]称之为“偏向性选择”。“偏向性选择”现象在小麦育种中普遍存在,如青农2号中54.1%的遗传物质来源于三交亲本鲁麦14[9];百农AK58更多地继承了三交亲本周麦11的遗传物质(47.4%)[10];川麦104中60.8%的遗传物质来源于单交亲本川麦42[11];周麦23遗传了单交亲本周麦13高达63.3%的遗传成分[12];目前,育种家普遍采取的育种策略是改良当地主推品种或骨干亲本的主要缺陷,而其大部分遗传物质得以保留,从而使新品种拥有更好地适应性,这种在育种过程中对目标性状的定向选择,可能是产生“偏向性选择”的主要因素[9]。本研究中,淮麦33也出现了“偏向性选择”现象,其73.9%的遗传物质来源于母本烟农19,进一步的分析也表明,烟农19中与产量性状相关的重要染色体区段受到强烈选择而较多地保留在淮麦33中。在生产中,淮麦33较多地遗传了烟农19株叶型清秀、大穗多粒和高产稳产的优良特性,有效改良了烟农19高秆易倒伏的缺陷,具有大面积推广种植的潜力。
4 结论
绘制了小麦新品种淮麦33的基因型图谱,明确了其遗传构成,淮麦33更多地继承了母本烟农19的遗传物质;淮麦33产量三因素协调,丰产性优于双亲烟农19和郑麦991;在Glu-B1位点含有17+18优质亚基;共有10个染色体区段与产量性状相关,6个来源于烟农19,3个来源于郑麦991,1个为淮麦33特异区段。致谢:本研究得到南京农业大学马正强教授、谢全老师、冉从福老师和杨阳博士以及西北农林科技大学李学军教授、强琴琴博士和刘玉秀博士的大力帮助,在此表示感谢。
The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}