0 引言
【研究意义】柑橘是世界范围内栽培面积最大的果树作物之一,但由于高温、伏旱等原因导致了柑橘大量异常落花落果,严重影响了产量。植物CEP基因可参与逆境响应,并对激素处理也具有不同的响应模式。因此,本研究基于柑橘全基因组对CEP基因家族进行全面分析,深入了解柑橘生长发育和逆境响应的分子基础,研究结果对进一步培育或改良柑橘抗性品种具有重要意义。【前人研究进展】许多植物多肽(多肽激素)具有类似生长素(IAA)、乙烯(ETH)和赤霉素(GA3)等激素的调节功能,广泛参与植物的一系列生长发育过程、花粉与柱头识别、防御反应、介导植物细胞间信号的转导和传递等[1,2,3,4,5,6,7,8,9]。多肽激素以配基的形式与细胞膜表面的相应受体激酶互作,从而激活下游基因、启动相关信号转导[10]。编码植物多肽激素的基因家族主要包括CLE家族、RGF家族、ERF家族和CEP家族等[10,11,12,13,14,15,16]。其中CEP(C-TERMINALLY ENCODED PEPTIDE)基因广泛存在于种子植物中,在进化上比较保守。CEP蛋白含一个或者多个CEP结构域[16],在CEP结构域的C-末端存在保守的蛋白序列(SPGV/IGH)[17,18,19]。在拟南芥中,ROBERTS等[19]根据结构域中脯氨酸数目的不同将CEP家族分为Group I和Group II,其中,Group I的CEP结构域含有3个脯氨酸,包括AtCEP1—AtCEP12,Group II的CEP结构域含有2个脯氨酸,包括AtCEP13、AtCEP14、AtCEP15。单子叶植物和双子叶植物的CEP结构域不同,其中单子叶植物的CEP结构域中有2个位点缺少苯丙氨酸残基,然而,这2个位点在双子叶植物中却非常保守,并且所有双子叶植物的CEP结构域的最后一个氨基酸均为组氨酸,而单子叶植物的CEP结构域末端氨基酸为组氨酸或天冬氨酸[18]。OGILVIE等[17]研究发现,禾本科植物的CEP基因碱基序列中GC含量明显高于其他植物种类。在拟南芥中,AtCEP5与受体XYLEM INTERMINXED WITH PHLOEM 1(XIP1)/CEP RECEPTOR 1(CEPR1)作用,从而调控主根长度和侧根密度[20,21]。AtCEP2的超表达使得根系变短,同时出现开花期延迟、叶片数目减少,其他CEP基因成员也呈现类似功能,但也存在差异[18,19]。在其他植物中,CEPs也参与植物根/茎生长、侧根发育以及根瘤分化等生长发育过程[16,18-19,22]。ROBERTS等通过在线数据库eFP[23]和Genevestigator[24]分析了拟南芥CEP基因在不同器官组织中的表达,如AtCEP1主要在顶端分生组织,AtCEP3和AtCEP9主要在芽顶端分生组织中表达,其中AtCEP3也在茎尖分生组织中表达,AtCEP1和AtCEP15在主根的根尖分生组织中表达,AtCEP3、AtCEP9、AtCEP13和AtCEP15主要在子叶或叶中表达,在侧根形成和发育过程中,这些CEP基因均可转录表达[19]。进一步探索发现CEP家族基因在同一组织器官的不同位置其表达模式也有差异,例如在5日龄幼苗的子叶中,AtCEP2在叶脉中表达,AtCEP3在叶尖端表达,在两周龄的叶中,AtCEP1在叶缘中表达,AtCEP2和AtCEP5在叶柄中表达[19]。此外,已有研究表明CEP基因可参与逆境响应,例如拟南芥AtCEP5和AtCEP15在水杨酸(SA)处理的条件下表达量降低;而赤霉素能诱导拟南芥AtCEP5和AtCEP13的表达;脱落酸(ABA)能使AtCEP15表达下调;茉莉酸能使AtCEP12表达上调等;AtCEPs家族成员间对生长素的应答模式存在差异,如该激素可诱导AtCEP1和AtCEP3表达升高,却抑制AtCEP5和AtCEP9的表达;同样,油菜素内酯可以使AtCEP5的表达降低而诱导AtCEP15的表达;另外,乙烯、内脂、细胞分裂素等对AtCEPs家族基因的表达没有显著的影响[19],超表达MtCEP1能够显著抑制侧根数目,RNA干扰MtCEP1、MtCEP2、MtCEP5、MtCEP11能够促进侧根生长[6,10,22,25]。【本研究切入点】CEP肽激素广泛参与植物根系在逆境下的生长发育调控,因此受到众多研究者的关注,但是目前对CEP家族的研究主要集中在模式植物如拟南芥、玉米、水稻和苜蓿,在柑橘中对该基因家族进行鉴定和分析尚未见报道。【拟解决的关键问题】基于柑橘全基因组鉴定柑橘CEP家族成员,并对该基因家族进行生物信息学分析,同时利用实时荧光定量PCR技术(qRT-PCR)检测该基因在不同组织中以及多种胁迫条件下的表达模式,解析CEPs调控柑橘根系生长发育及其逆境响应的分子基础。1 材料与方法
1.1 试材与处理
选用‘资阳香橙’(‘Citrus junos Ziyang’)为试材,其种子为多胚,苗由珠心胚发育而来,具有一致的遗传背景。2016年11月在中国农业科学院柑桔研究所国家柑橘种质资源圃采集‘资阳香橙’种子,去除内外种皮,放在湿润的纱布上,塑料膜保湿;然后放入28℃的培养箱中暗培养10 d,进行催芽。当根长至2 cm左右时将幼苗转入营养液(250 µmol·L-1 KH2PO4,3 000 µmol·L-1 KNO3,2 000 µmol·L-1 Ca(NO3)2,250 µmol·L-1 MgSO4,25 µmol·L-1 MgCl2,12.5 µmol·L-1 H3BO3,1 µmol·L-1 MnSO4,1 µmol·L-1 ZnSO4,0.25 µmol·L-1 CuSO4,0.25 µmol·L-1 (NH4)6Mo7O24,25 µmol·L-1 Fe-Na-EDTA,pH 5.8)中进行水培[26],当第一片真叶充分展开后,选取长势一致的幼苗进行激素和非生物胁迫处理。激素包括ABA(100 µmol·L-1)、SA(100 µmol·L-1)、MeJA(茉莉酸甲酯,100 µmol·L-1)、IAA(20 µmol·L-1)、GA3(0.5 m mol·L-1)、ETH(1.5 mmol·L-1)、6-BA(6-苄氨基腺嘌呤,0.05 mmol·L-1),具体步骤如下:在室温下,将幼苗洗净,放入盛有激素或纯净水(平行对照)的灭菌瓶中摇匀,使根系充分接触液体。在处理后的0(对照)、3、6、12、24 h分别收集幼苗根系,立刻放入液氮中进行速冻,-80℃贮藏备用。高盐NaCl(300 mmol·L-1)处理及取样方法与激素处理相同[27]。干旱胁迫采用纸巾包裹幼苗根系,放置室温下使其脱水,分别在0、1、3、6和12 h收集根系,液氮速冻,-80℃贮藏备用。每个处理至少5株幼苗。采集‘资阳香橙’实生幼苗的根、茎、叶、子叶,及其成熟果子的果皮和种子,液氮速冻,-80℃贮藏,用于分析柑橘CEP家族成员表达的组织特异性。
1.2 总RNA提取与cDNA合成
植物RNA提取试剂盒(RNA prep pure Kit)购自天根生化科技有限公司,用于抽提总RNA。使用TaKaRa公司的反转录试剂盒(Prime ScriptTM RT Reagent Kit)合成第一条cDNA链。上述试验均按照试剂盒说明书进行。1.3 基因的结构与序列分析
以拟南芥CEP蛋白为参考序列,采用BLASTp在phytozome数据库(https://www.chinaagrisci.com/article/2018/0578-1752/www.phytozome.net)中进行比对,鉴定出柑橘CEP基因(CitCEPs)成员;用EXPASY软件(http://web.expasy.org/protparam/)分析蛋白编码的氨基酸数目、相对分子量以及等电点等理化性质[28];ProtScale Tool软件(http://web.expasy.org/protscale/)分析蛋白质的亲水性/疏水性[29],基因结构采用GSDS软件(http://gsds.cbi.pku.edu.cn/)进行分析并绘制结构图。用CLUSTALX对蛋白多序列进行比对,采用MEGA6.0软件邻接法构建NJ系统进化树。采用MBC(http://cello.life.nctu.edu.tw)软件对CitCEP蛋白进行亚细胞定位预测。采用在线软件plantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)对启动子顺式元件进行分析。1.4 实时荧光定量分析
引物由Primer5.0设计,引物信息见表1。以柑橘Actin为内参,在Bio-Rad CFX3.1荧光定量PCR仪上对CEP基因各成员进行定量分析。反应体系为10 µL:5 µL荧光染料(Bio-Rad公司)、3 µL无菌水、0.5 µL正向引物、0.5 µL反向引物、1 µL cDNA。反应程序为:95℃ 30 s;95℃ 5 s,55℃ 5 s,39个循环。每个处理3次重复。相对表达量采用2-ΔΔCt法计算,用SPSS19.0软件对数据进行统计分析。Table 1
表1
表1本试验所用引物
Table 1Primers used in this study
| 引物名称 Primer name | 正向引物序列 Forward primer sequence | 反向引物序列 Reverse primer sequence |
|---|---|---|
| CitCEP1 | CCGAGCAAAGGCTTCTAA | CATCCGCTGAAAGTCTGG |
| CitCEP2 | GCTGCTAAAAGATTGTGGAG | CCCTGAAAGTCTGGGAAT |
| CitCEP3 | CCTGCGGCTTATTACCAT | AACATCTCCTTGCCTTCTG |
| CitCEP4 | TGTCCGTGGAGGGAAGAA | CCTTGGCTTGGCGTGATT |
| CitCEP5 | CCTTTGTTTGTCTTCTCCTC | ACTGTAGTGTACTCATCACCGT |
| CitCEP6 | GTTCTCCTCCTAAGCAAAAG | CAACACCAGGGCTATGTC |
| CitCEP7 | CATGGCGTGGTTCAAGAT | GCTAGGACCCGAGTTCTTT |
| CitCEP8 | GCGACGTTGAAGGAAGAA | AACCCCTCCGTAGTTTTAG |
| CitCEP9 | CTGGAAACAGCCCTGGAGT | GTGATGTGGGGCGGAAAT |
| CitCEP10 | TCTTTGTCGTGTTTGCTTCG | CGGCACCATGAGTCTTCTT |
| CitCEP11 | GAGAAGATCGACAGCAGC | CAGAATCCGATCAATGCC |
| Actin | CCCCATCGTTACCGTCCAG | CGCCTTGCCAGTTGAATATCC |
新窗口打开
2 结果
2.1 CitCEP基因家族成员的鉴定与生物信息学分析
2.1.1 柑橘CitCEP基因的鉴定 以拟南芥CEP蛋白序列为参考,基于Phytozome数据库采用BLASTp从柑橘全基因组中鉴定出11个CitCEP成员(表2),其中只存在于克林曼丁橘的有4个,即CitCEP3、CitCEP4、CitCEP7、CitCEP10;甜橙的有2个,CitCEP9和CitCEP11,而CitCEP1、CitCEP2、CitCEP5、CitCEP6、CitCEP8在甜橙和克林曼丁橘基因组上均含有。通过蛋白序列比对发现,C-末端均存在该家族的特征结构域(SPGV/IGH)(图1)。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1CitCEP家族蛋白序列的氨基酸聚类分析
-->Fig. 1Amino acid sequence alignment of citrus CitCEPs
-->
Table 2
表2
表2CitCEP家族成员蛋白质的理化性质
Table 2Physicochemical property of CitCEP proteins
| 基因 Gene | 登录号 Locus ID | 基因长度 Genome length (bp) | 氨基酸数目 Number of amino acid | 分子量 Molecular weight (kD) | 等电点 pI | 不稳定指数 Instability index | 脂肪系数 Aliphatic index | 平均疏水指数 Grand average of hydropathicity |
|---|---|---|---|---|---|---|---|---|
| CitCEP1 | Ciclev10030344/orange1.1g048132 | 354 | 117 | 12144.33 | 6.15 | 30.70 | 63.42 | -0.456 |
| CitCEP2 | Ciclev10003677/orange1.1g041852 | 351 | 116 | 12002.22 | 6.15 | 31.91 | 67.33 | -0.366 |
| CitCEP3 | Ciclev10033210 | 255 | 84 | 9449.75 | 7.89 | 42.76 | 76.79 | -0.332 |
| CitCEP4 | Ciclev10027212 | 273 | 90 | 9826.49 | 10.37 | 52.87 | 81.22 | -0.084 |
| CitCEP5 | Ciclev10027209/orange1.1g040634 | 360 | 119 | 13157.97 | 6.90 | 48.82 | 85.21 | -0.521 |
| CitCEP6 | Ciclev10017790/orange1.1g038829 | 261 | 86 | 9339.88 | 10.21 | 49.11 | 82.79 | -0.242 |
| CitCEP7 | Ciclev10029992 | 303 | 100 | 10921.70 | 9.80 | 48.32 | 78.00 | -0.305 |
| CitCEP8 | Ciclev10027106/orange1.1g045305 | 282 | 93 | 10051.52 | 8.35 | 40.80 | 72.37 | -0.242 |
| CitCEP9 | orange1.1g043660 | 345 | 114 | 12122.28 | 6.04 | 57.86 | 59.12 | -0.811 |
| CitCEP10 | Ciclev10018148 | 865 | 141 | 15528.08 | 10.09 | 45.50 | 98.23 | 0.198 |
| CitCEP11 | orange1.1g047704 | 852 | 98 | 10757.37 | 10.43 | 48.41 | 88.57 | -0.348 |
新窗口打开
2.1.2 基因结构与预测蛋白特性分析 基于柑橘全基因组,分别获得柑橘CitCEP成员的mRNA及其基因组DNA序列,利用GSDS在线软件对各成员的基因结构进行了分析。结果表明,CitCEP1—CitCEP9均无内含子,而CitCEP10和CitCEP11含有1个内含子(图2)。采用EXPASY软件对预测蛋白的氨基酸数目、等电点、分子量、不稳定系数、脂肪系数、平均疏水指数进行分析(表2)。结果显示,柑橘CEP家族成员均为亲水性蛋白,CitCEP5的第12位氨基酸的亲水性最强,其值为3.711,CitCEP4的第32位氨基酸疏水性最强,其值为-2.778。从表2可看出,CitCEP成员之间基因组长度变化明显,其中CitCEP3最短,只含有255个碱基,可编码84个氨基酸,而CitCEP10最长,具有865个碱基,可编码141个氨基酸,但其分子量最高,为15 528.08 kD。该基因家族成员的等电点介于6.04—10.43,不稳定指数介于30.70—57.86,脂肪系数介于59.12—98.23,平均疏水指数介于-0.811—0.198。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2CitCEP成员的基因结构
-->Fig. 2Gene structure of CitCEP genes
-->
2.1.3 CitCEP蛋白的亚细胞定位预测与亲缘关系 利用Cello软件对柑橘CEP家族成员的亚细胞定位进行了分析,结果表明CitCEP1、CitCEP2、CitCEP4、CitCEP5、CitCEP6、CitCEP8定位在细胞外,CitCEP3和CitCEP7在细胞外和线粒体中均存在,CitCEP9定位在细胞核中,CitCEP10定位于细胞质膜上,而CitCEP11存在于线粒体和细胞核中。
为了解柑橘CEP成员间的亲缘关系,推测其生物功能,利用MEGA6.0软件构建了拟南芥和柑橘CEP蛋白成员的NJ进化树(图3)。从图3可以看出,拟南芥CEP成员与柑橘CEP成员相互嵌合在一起,形成3大分支(G1、G2和G3),表明植物CEP基因在进化过程中相对保守。G1含有成员最多共计17个,包括4个柑橘CitCEPs和13个AtCEPs;G2由CitCEP1、CitCEP2、CitCEP3和AtCEP5组成;G3有CitCEP8、CitCEP9、CitCEP10、CitCEP11和AtCEP3组成。在这些成员中CitCEP5与AtCEP7,CitCEP4与AtCEP1,CitCEP1、CitCEP2与AtCEP5,CitCEP9与AtCEP3的亲缘关系较近,推测这些CEP具有类似的生物功能;其他柑橘CitCEP成员与拟南芥成员的亲缘关系较远,表明这些成员之间的生物功能可能差异较大。
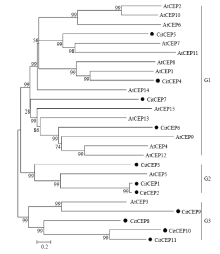 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3柑橘与拟南芥CEP家族蛋白的系统进化树(邻接法)
-->Fig. 3Phylogenetic tree of CEP protein sequences from citrus and Arabidopsis (NJ method)
-->
2.2 CitCEP家族基因的表达分析
2.2.1 组织特异性 柑橘CitCEP成员间具有明显不同的组织特异性。其中,CitCEP1、CitCEP4、CitCEP5、CitCEP6、CitCEP8和CitCEP9在各组织中均未检测到明显的表达信号。CitCEP2在茎中表达量最高,叶中次之,在根和子叶中较低,果皮和种子中几乎不表达。CitCEP3主要在根和子叶中表达,在茎、叶、种子中表达很低,而在果皮中几乎不表达;CitCEP10和CitCEP11均显著在果皮中表达,而在茎、果皮、种子和叶中表达极低或不表达(图4)。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4CitCEP2、CitCEP3、CitCEP10、CitCEP11在不同组织中的相对表达量
-->Fig. 4Relative expression of CitCEP2, CitCEP3, CitCEP10, CitCEP11 in different tissues
-->
2.2.2 非生物胁迫与激素对CitCEP表达的影响 CitCEP1、CitCEP4、CitCEP5、CitCEP6、CitCEP7、CitCEP8和CitCEP9对干旱、盐胁迫、ABA等各处理响应不显著,其表达很低或不表达。而CitCEP2、CitCEP3、CitCEP10和CitCEP11对各处理表现出不同的表达模式。在盐胁迫和干旱条件下,CitCEP2的表达量均受到抑制,而CitCEP3在盐胁迫条件下表达量先上调后下降,最后恢复至对照水平,在干旱条件下其表达量显著上调;在干旱处理下,CitCEP10和CitCEP11的表达量均显著上调,呈现相似趋势,而在盐胁迫下,CitCEP10表达显著受到诱导,CitCEP11却在处理3 h后出现显著上调后恢复到对照水平。上述结果表明,CitCEP2、CitCEP3、CitCEP10和CitCEP11参与了柑橘的非生物胁迫应答(图5)。在ETH、ABA处理下,CitCEP2的表达量明显受到抑制,而在MeJA、SA、IAA、GA3处理下均表现为先上升、后下降的趋势;在6-BA、IAA、GA3、ABA处理下CitCEP3也表现出类似趋势。CitCEP10在ETH条件下表达显著上升,在6-BA、SA、ABA处理条件表达呈现先下降后上升的趋势,在MeJA、IAA、GA3处理下无明显规律。CitCEP11在ETH、MeJA、SA、IAA、GA3、ABA处理下均呈现表达下调趋势,与CitCEP3在ETH、SA处理下的表达模式类似。在6-BA条件下,CitCEP11表达呈现先下降后上升趋势(图6)。综上所述,CitCEP2、CitCEP3、CitCEP10和CitCEP11可能与柑橘的生长发育和胁迫响应有关。
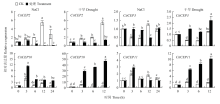 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5非生物胁迫下CitCEP2、CitCEP3、CitCEP10、CitCEP11的相对表达量
-->Fig. 5Relative expression of CitCEP2, CitCEP3, CitCEP10, CitCEP11 under different abiotic stresses
-->
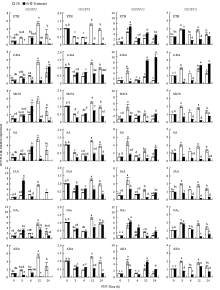 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图6不同外源激素处理条件下CitCEP2、CitCEP3、CitCEP10、CitCEP11的相对表达量
-->Fig. 6Relative expression of CitCEP2, CitCEP3, CitCEP10, CitCEP11 under different treatments of exogenous plant hormones
-->
为了进一步说明CitCEP2、CitCEP3、CitCEP10和CitCEP11对各处理的应答机制,利用柑橘全基因组分别获得各基因起始密码子前2 000 bp的启动子序列。采用plantCARE在线软件对顺式作用元件分别进行了预测分析,结果显示CitCEP2、CitCEP3、CitCEP10和CitCEP11的启动子中包含了多个与逆境和激素相关的顺式作用元件(表3)。其中,CitCEP2、CitCEP3、CitCEP10和CitCEP11均包含光响应元件(ATCT-motif、G-box)、热应激响应元件(HSE)和水杨酸响应元件(TCA-element)。另外,CitCEP3、CitCEP10和CitCEP11还含有ACE光响应元件、脱落酸响应元件(ABRE)和低温响应元件(LTR),而赤霉素响应元件(GARE-motif)只存在于CitCEP2、CitCEP10、CitCEP11启动子中。CitCEP2和CitCEP3启动子含有与干旱相关的MBS元件。此外,CitCEP2启动子还含有冷和脱水胁迫相关的响应元件(C-reapt/DRE),CitCEP10和CitCEP11含有MeJA响应元件(TGACG-motif)。
Table 3
表3
表3PlantCARE预测CitCEP2、CitCEP3、CitCEP10、CitCEP11启动子区顺式作用元件
Table 3Cis-acting regulatory elements in promoter of CitCEP2, CitCEP3, CitCEP10, CitCEP11 predicted by plantCARE
| 顺式元件 Cis-element | 典型序列 Typical sequence | 特性 Characteristic | 基因 Gene |
|---|---|---|---|
| ATCT-motif | AATCT | 光响应Part of a conserved DNA module involved in light responsiveness | CitCEP2, CitCEP3, CitCEP10, CitCEP11 |
| G-box | CACGT | 光响应Cis-acting regulatory element involved in light responsiveness | CitCEP2, CitCEP3, CitCEP10, CitCEP11 |
| ACE | AAAACGTTTA | 光响应Cis-acting element involved in light responsiveness | CitCEP3, CitCEP10, CitCEP11 |
| GARE-motif | TCTGTTG, AAACAGA | 赤霉素响应Gibberellin-responsive element | CitCEP2, CitCEP10, CitCEP11 |
| HSE | AGAAAATTCG, AAAAAATTTC | 热应激响应Cis-acting element involved in heat stress responsiveness | CitCEP2, CitCEP3, CitCEP10, CitCEP11 |
| MBS | TAACTG | MYB结合位点参与干旱诱导MYB binding site involved in drought-induction | CitCEP2, CitCEP3 |
| TCA-element | CAGAAAAGGA, CCATCTTTTT | 水杨酸响应Cis-acting element involved in salicylic acid responsiveness | CitCEP2, CitCEP3, CitCEP10, CitCEP11 |
| ABRE | GACACGTGGC TACGTG | 脱落酸响应Cis-acting element involved in the abscisic acid responsiveness | CitCEP3, CitCEP10, CitCEP11 |
| C-repeat/DRE | TGGCCGAC | 冷和脱水Regulatory element involved in cold- and dehydration-responsiveness | CitCEP2 |
| LTR | CCGAAA | 低温反应Cis-acting element involved in low-temperature responsiveness | CitCEP3, CitCEP10, CitCEP11 |
| TGACG-motif | TGACG | MeJA响应Cis-acting regulatory element involved in the MeJA-responsiveness | CitCEP10, CitCEP11 |
新窗口打开
3 讨论
植物多肽激素大约包含20个氨基酸,是一类经过剪切后形成的具有特殊功能的、成熟的多肽[30]。自1991年在番茄中发现了第一个多肽激素系统素,随后植物多肽激素受到越来越多的关注[31],CEP蛋白是由N末端的分泌信号、变异结构、一个或者多个CEP结构域和C-末端序列组成[16]。目前在植物基因组中鉴定出的CEP多肽主要存在于种子植物中,其中在拟南芥基因组共鉴定出15个,但有4个AtCEP家族基因(AtCEP7、AtCEP8、AtCEP10和AtCEP11)未被注释[16-19,22,32];水稻中鉴定出15个,玉米中鉴定到12个[10],苜蓿中鉴定出9个,苹果中迄今只报道了1个[33]。本研究从柑橘全基因组数据库中获得了11个CEP基因成员,与玉米中CEP成员数相当,而少于水稻和拟南芥中的成员数,表明不同物种的CEP成员数变化较大。该基因家族成员均为亲水性蛋白,且亚细胞定位预测结果显示主要分布于细胞外、线粒体、细胞质膜、细胞核中。序列分析发现,CitCEP10与CitCEP11的序列基本一致,且数据表达显示对各激素处理以及非生物胁迫的结果响应趋势相似度很高,但来源于不同的柑橘品种,因此推测CitCEP10与CitCEP11为不同等位变异类型,此种变异类型在水稻OsCEP中也有报道[10]。聚类分析发现,柑橘与拟南芥CEP成员相互嵌合分布于不同分支中,表明CEP在进化过程中保守性较强,这与ROBERTS等[19]的研究结果一致,即CEP家族在被子植物中的进化较为保守。柑橘CitCEP1、CitCEP2与拟南芥AtCEP5位于一个亚分支中,具有较近的亲缘关系。在拟南芥中敲除AtCEP5导致植物生长势有所增加;相反,过量表达AtCEP5致使转基因植株发育迟缓且不表现出向地性[19],由此推测CitCEP1、CitCEP2可能也与柑橘的生长发育和向地性有关。已有研究表明在渗透强度增加以及盐水平升高的条件下使得拟南芥芽中AtCEP1的表达量上调,但在根中却无响应[18],过量表达AtCEP1可抑制转基因植株的主根生长[16]。本试验中CitCEP4与AtCEP1具有较高的相似度,表明CitCEP4可能具有调控柑橘主根生长的生物功能。CitCEP5与AtCEP7相似度最高,但目前拟南芥AtCEP7功能还没有得到验证,有待于进一步研究。CitCEP9与拟南芥AtCEP3相似性最高,已有研究表明高氮、钾水平能使AtCEP3表达下调[19],超表达拟南芥AtCEP3可抑制根生长、导致叶片变小及黄化[18,19],由此可见CitCEP9可能与柑橘的营养调控有关,从而影响叶片和根系生长。其余柑橘CEP成员如CitCEP11与拟南芥CEP各成员的亲缘关系较远,表明它们之间的生物功能差异较大,有待于进一步研究。
本试验中,CitCEP1、CitCEP4、CitCEP5、CitCEP6、CitCEP7、CitCEP8和CitCEP9在所采集的幼苗时期的根、茎、叶、子叶以及果皮和种子中均无表达。这些成员可能为假基因或在其他特定组织和时期中表达[25]。已有研究表明,CEP基因参与了根、茎、芽、花的生长发育[10,16,18,22,32]。本试验中CitCEP2主要在茎、叶、子叶中表达,CitCEP3主要在根和子叶中表达,而CitCEP10和CitCEP11均在果皮中显著表达,表明CEP基因除了调控根、茎、芽、花的生长发育,同时可能也参与了叶、子叶和果皮发育。
本研究中,CitCEP1、CitCEP4、CitCEP5、CitCEP6、CitCEP7、CitCEP8和CitCEP9对各处理均无响应,此现象在其他基因家族中也存在[26]。拟南芥中,AtCEP的转录水平受逆境、营养水平和激素等调控,DELAY等对拟南芥9个AtCEP(AtCEP1、AtCEP2、AtCEP3、AtCEP4、AtCEP5、AtCEP9、AtCEP13、AtCEP14、AtCEP15)在不同浓度的N含量、盐胁迫及CO2处理条件下的表达进行了分析,表明除AtCEP15外其余8个基因均有不同程度的响应[18]。另外,这些基因的表达水平受水杨酸、赤霉素、脱落酸、生长素等激素处理影响[19]。在本研究中,CitCEP2、CitCEP3、CitCEP10与CitCEP11对激素、干旱和盐胁迫均有响应,与前人研究结果类似[19]。由此推测CitCEP2、CitCEP3、CitCEP10与CitCEP11可能参与了植物生长发育以及逆境响应调控。对CitCEP2、CitCEP3、CitCEP10与CitCEP11的启动子分析发现,其启动子均含有逆境、激素、温度等响应顺式作用元件如G-box光响应元件、TCA水杨酸响应元件、HSE热应激响应元件等,该结果为柑橘CEP基因对逆境和激素的应答机制提供了进一步证明。
4 结论
从柑橘全基因组上鉴定出了11个CitCEP基因成员,其蛋白均为亲水蛋白且含有SPGV/IGH保守结构域。除CitCEP10和CitCEP11含有1个内含子外,其余成员均无内含子,亚细胞定位预测显示柑橘CEP家族基因成员在细胞外、线粒体、细胞核、细胞质膜中均存在,通过实时荧光定量PCR技术对柑橘CEP基因家族进行表达分析,结果表明CitCEP1、CitCEP4、CitCEP5、CitCEP6、CitCEP7、CitCEP8和CitCEP9在‘资阳香橙’的根、叶、子叶、果皮、种子中几乎不表达,对激素与非生物胁迫均不响应。CitCEP2、CitCEP3、CitCEP10、CitCEP11的组织特异性差异明显,且对激素和非生物胁迫有不同程度的响应。(责任编辑 岳梅)
The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}