0 引言
【研究意义】芝麻(Sesamum indicum L.)是中国重要的优质油料作物,分布广,但主要集中在江淮、黄淮地区,雨热同季,易受到多种真菌、细菌病害的侵染,其中最严重的病害是茎点枯病(Macrophomina phaseolina Ashby)[1]。茎点枯病的病原菌主要通过种子、土壤及病残体越冬传播,能以菌核或分生孢子形式存活10个月以上,侵染玉米、大豆、芝麻等100多个属的500多种作物[2-3],一般造成减产10%—20%,严重时达到80%以上[4]。因此,迫切需要选育高抗病或高耐病的芝麻品种。但是,现有芝麻栽培品种均来自于同一个种(S. indicum),遗传基础相对狭窄[5-6]。现有的芝麻种质资源里缺少免疫类型,而高抗类型也很少,导致芝麻茎点枯病的遗传改良无法取得突破性进展[4, 7-8]。研究发现,芝麻属中部分野生种具有高抗病抗逆等优良特性[9-10],因此,发掘、利用芝麻野生种的抗茎点枯病优良基因,丰富芝麻栽培种的基因库,是芝麻茎点枯病遗传改良的重要途径[3]。【前人研究进展】近30年来,国内外的专家****先后开展了大量的芝麻远缘杂交研究,以期利用野生芝麻的优良性状。已有研究表明,芝麻种间杂交存在一定的杂交不亲和性或杂种发育障碍,利用常规杂交方法很难获得杂种后代[11-12]。借助胚胎培养拯救技术是获得种间杂交后代的有效方法,已在野生种S. schinzianum、S. radiatum、S. alatum等与栽培种的杂交上取得成功[9, 12-15]。在杂交种真实性的鉴定上,除形态学观察外,也普遍使用细胞学、分子标记等方法[9,13-15]。RAJESWARI等[13]以S. alatum和栽培种杂交,获得抗变叶病材料。【本研究切入点】尽管前人在一些芝麻野生种与栽培种的杂交上取得成功,但关于野芝3号(S. indicatum)茎点枯病的抗性转移研究未见报道。【拟解决的关键问题】本研究通过对野芝3号与栽培种中芝14、中芝14同源四倍体进行种间杂交,借助幼胚培养技术及细胞学、分子标记、茎点枯病人工接种鉴定技术,获得高抗茎点枯病新种质,为芝麻茎点枯病抗病育种提供基础遗传材料。1 材料与方法
1.1 试验材料
供试材料亲本为P1:中芝14(2n=26)、P2:中芝14同源四倍体(2n=52)、P3:野芝3号(2n=58 S. indicatum)。茎点枯病抗性鉴定的对照材料:芝麻育种课题组经过多年在湖北武汉病害鉴定圃鉴定出的稳定新株系,包括抗病对照BJ2236、中抗对照BJ2018和感病对照BJ2002。
菌种:由河南省农业科学院植物保护研究所刘红彦研究员提供。
1.2 种间杂交以及F1植株的获得
种间杂交试验于2014年夏在中国农业科学院油料作物研究所武昌基地进行。于开花前一天下午摘掉母本的花冠和雄蕊,第二天上午用父本花粉授粉。取杂交授粉后16 d的幼蒴,进行表面消毒,切开幼蒴,剥取幼胚,接种到幼胚诱导培养基上(MS+30 g·L-1蔗糖+8 g·L-1琼脂粉+0.5 g·L-1 GA3+1 g·L-1 6-BA,pH=5.8)。14 d后转入分化培养基(MS+30 g·L-1蔗糖+8 g·L-1琼脂粉+0.3 g·L-1 IBA,pH=5.8)继续培养,20 d后在生根培养基(MS +30 g·L-1蔗糖+8 g·L-1琼脂粉+0.5 g·L-1 NAA,pH=5.8)上壮苗生根,直至发育成幼苗,控制温度28—30℃、光周期8—16 h·d-1、光照强度1 500—2 000 lx。将杂种F1 植株在室内自然光下开瓶炼苗2—3 d,洗净附着在其表面的培养基,将其移栽到装有营养土(泥炭﹕珍珠岩﹕蛭石=1﹕1﹕1)的穴盘中,在27—30℃、60%—70%相对湿度条件下培养5 d后带土移栽大田。幼胚萌发并形成健壮小苗时统计幼胚成苗率。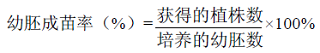
1.3 细胞学观察
以植株花期的子房为材料,进行细胞有丝分裂观察。子房的大小为2 mm左右,在解剖镜下除去花药,常温下用0.002 mol·L-1 8-羟基喹啉混合液避光处理4 h,加入新鲜卡诺固定液(冰乙酸﹕无水乙醇=1﹕3)在4℃冰箱中固定18—24 h。清水冲洗后,转入1 mol·L-1 HCl 60℃恒温解离8 min,用蒸馏水漂洗,卡宝品红染色,蒸馏水压片,敲片,镜检时取30个分裂相良好的细胞观察染色体的形态,统计染色体数目,用OLYMPUS显微镜(BX-51)拍照。1.4 DNA提取、检测和SSR分析
选取健康无病害供试材料的幼嫩叶片50 mg,参照PATERSON等[16]改良的CTAB法提取基因组DNA,-20℃保存备用。随机从已合成的EST-SSR引物中挑选265对引物[17]进行双亲之间的多态性筛选。通过选用与双亲有特异性标记的引物对杂种F1进行分子鉴定。PCR扩增反应体系和扩增程序参考WU等[17]。扩增产物在6%聚丙烯酰胺凝胶中电泳,银染,检测,同时具有父母本特异性带的即为真杂种。1.5 形态学观察
2015年夏分别在现蕾期、花期和成熟期调查后代植株及亲本形态性状,主要包括植株生长习性、株型、茎秆颜色、花冠长唇缘颜色、每叶腋花数、花粉粒育性、蒴果形状、单株蒴果数、种皮颜色、全生育期等。1.6 茎点枯病抗性鉴定
2016年对抗病、中抗、感病等3个稳定的对照材料、种间杂交的3个亲本(野芝3号、中芝14及其同源四倍体)以及杂种后代F1株系进行茎点枯病菌的接种抗性鉴定。参考王建方[18]的接菌方法,在盛花期进行,每个材料接种5株,重复3次,挂牌记录。选取灭菌后的棉花,在菌丝液中浸泡待用,取灭菌后的镊子在芝麻茎杆中下部戳一个3 mm左右的小孔,用棉花蘸取菌液对小孔周围进行均匀涂抹,高度为2 cm。对照:只涂抹无菌水。在7 d后进行植株发病情况调查记录,参照江诗洋等[19]的方法测量发病植株的病斑长度,结果取平均值。在SPSS软件上进行差异显著性方差分析,用Origin软件绘图。2 结果
2.1 种间杂交
野芝3号(P3)与栽培种(P1和P2)共配制4个正、反杂交组合,授粉511朵花,获得277个蒴果。在田间自然条件下,虽然能获得蒴果但是蒴果成熟后多数瘪而小,内无籽或仅有一两粒种子,多数无发芽能力。而采用胚胎拯救技术对授粉后16 d的幼胚进行组织培养,在接种的773个幼胚中,有155个幼胚发育成苗,平均幼胚成苗率为20.05%(表1)。发现正反交组合的幼胚成苗率存在明显差异,以野芝3号为母本组合(P3×P1、P3×P2)的幼胚成苗率均高于其反交组合(P1×P3、P2×P3),其中,P3×P1组合成苗率高达32.75%。以野芝3号为父本的反交组合的幼胚成苗率均低于平均值,其中较高的P2×P3组合成苗率为13.41%。结果表明,亲本的基因型在很大程度上影响远缘杂交的成苗率。Table 1
表1
表1野芝3号与2个栽培种种间杂交后代成苗率比较
Table 1Comparison of the seedling ratios among the reciprocal crosses between No.3 wild sesame and the two cultivated accessions
| 杂交组合 Combination | 母本 Female parent | 父本 Male parent | 授粉花数 No. of flowers pollinated | 蒴果数 No. of capsules obtained | 幼胚数Immature embryo number | F1株系数 F1 line number | 幼胚成苗率 Seedling ratio (%) |
|---|---|---|---|---|---|---|---|
| P3×P1 | 野芝3号(P3) S. indicatum | 中芝14(P1) Zhongzhi 14 | 135 | 89 | 229 | 75 | 32.75 |
| P3×P2 | 野芝3号(P3) S. indicatum | 中芝14同源四倍体(P2) Autotetraploid of Zhongzhi 14 | 128 | 71 | 199 | 42 | 21.11 |
| P1×P3 | 中芝14(P1) Zhongzhi 14 | 野芝3号(P3) S. indicatum | 130 | 53 | 181 | 16 | 8.84 |
| P2×P3 | 中芝14同源四倍体(P2) Autotetraploid of Zhongzhi 14 | 野芝3号(P3) S. indicatum | 118 | 64 | 164 | 22 | 13.41 |
新窗口打开
2.2 种间杂种F1的细胞学和分子标记鉴定
对野芝3号与中芝14、中芝14同源四倍体以及种间杂交后代的子房有丝分裂中期染色体数目进行统计,结果表明,P3×P1 、P1×P3 组合F1植株染色体数目为42(图1-A4)。P3×P2、P2×P3 组合F1染色体数目为55(图1-A5)。正反交杂种F1株系大部分花粉粒形态独特,形状规则但多无内含物,为高度不育类型;部分F1株系有少量的可育花粉,为部分不育型(图1-B4—图1-B5)。调查发现F1植株在田间有自交结实现象,F1植株自交所得籽粒颜色分离,体积较大,部分饱满,有一定的萌发率。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1亲本及种间杂交后代的染色体和花粉粒比较
-->Fig. 1Chromosome and pollen comparison between parents and interspecific hybrids
-->
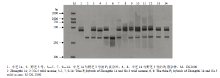
从265对SSR引物[17]进行的双亲多态性筛选中,检测出12对标记在亲本间具有多态性。选用其中部分多态性较好的SSR标记对种间正反交F1株系进行分子鉴定。图2中所用的引物为HS142(F:5′-ATT GTCGTTGTCGTTGTCGT-3′,R:5′-AACTCCATCAA CCTATGCCC-3′)。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2种间杂种F1的SSR标记扩增图谱(HS142)
-->Fig. 2Detection of F1 interspecific hybrids by SSR marker (HS142)
-->
如图2所示,HS142引物在亲本中芝14中能清晰地扩增出2条特异性条带(约460 bp、500 bp),在野芝3号则扩增出1条特异性条带(约380 bp)。12个后代中,有10个杂种F1植株同时具有3条父、母本特异条带,另2株仅出现母本或父本带。结合其他引物PCR结果,判定该10株为真杂种,另外2株为假杂种。田间观察发现,假杂种表型性状与父本或母本无差异,证实了PCR结果。
2.3 杂种F1形态学特征
随机选取20个正反杂交组合的F1植株,分别在现蕾期、花期和成熟期观察其形态特征(表2),正反交杂种F1的蒴果形状、叶片基部边缘锯齿状、每叶腋花数为单花等都趋向于亲本野芝3号;花冠长唇缘颜色浅紫、茎秆颜色浅紫等则介于双亲之间(图3)。不同正反杂交组合的杂种F1植株生长习性、株型、全生育期、种皮颜色等性状均存在差异,如以野芝3号为母本的组合P3×P1、P3×P2杂种F1植株形态上表现为匍匐型、分枝,种皮颜色均为黑色,与野生种亲本相似;反交组合P1×P3的杂种F1种皮颜色为白色,与栽培种亲本相似;植株形态则大部分为半直立,株型也多为单秆或少分枝,性状居于两亲本之间(表2)。Table 2
表2
表2种间杂种后代及亲本特征比较
Table 2Morphological comparison between the parents and hybrids
新窗口打开
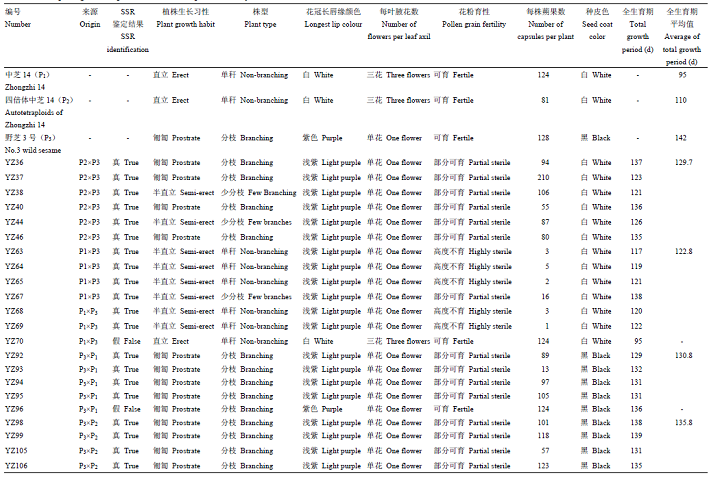

图3种间杂交F1与亲本的生物学性状比较
-->Fig. 3Morphological comparison between the F1 interspecific hybrids and the parents
-->
2.4 亲本及杂种F1茎点枯病抗性鉴定
芝麻茎秆在伤口处涂抹菌液后,伤口附近会形成棕黑色病斑并逐渐扩大,接菌3 d后感病材料BJ2002开始出现侵染症状,病斑迅速扩大;抗病材料BJ2236病斑扩大比较缓慢,而野芝3号(P3)以及其种间杂交后代伤口的棕黑色病斑无明显变化;接菌7 d后,不同对照材料的病斑长度差异非常明显,因此,作为品种抗性评价的依据。由图4可见,抗病对照材料BJ2236的病斑长度为11.41 cm;中抗对照材料BJ2018的病斑长度为15.03 cm;感病对照材料BJ2002的病斑长度为19.52 cm,3个对照材料的染病结果与之前多年的人工病圃鉴定结果表现基本一致,说明此试验准确性较好,用于芝麻茎点枯病抗性鉴定是合适的。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4基于3个对照、3个亲本、4个杂交后代的茎点枯病病情指数
-->Fig. 4Disease index of charcoal rot on 3 control materials, 3 parents and 4 hybrids
-->
本研究结果显示,抗病、中抗、感病对照材料、3个亲本以及杂种F1对茎点枯病菌的抗性存在显著差异。亲本野芝3号(P3)的抗性与抗病对照材料BJ2236有显著差异,抗性最强;中芝14(P1)与中抗对照材料BJ2018无显著差异;同源四倍体(P2)与抗病对照材料BJ2236无显著差异。中芝14(P1)的病斑长度显著高于同源四倍体(P2)的病斑长度(14.30 cm>11.46 cm),说明同源四倍体的抗性要显著强于中芝14。后代中,P3×P2的杂种F1病斑最短为6.65,显著低于其他杂交组合后代和抗病对照材料的病斑长度;其他杂交组合F1的病斑长度为8.90—9.90 cm,显著低于抗病对照材料BJ2236。野芝3号与同源四倍体的后代在茎点枯抗性上均强于与中芝14杂交的后代。同样的亲本,以野芝3号为母本的正交组合在后代茎点枯抗性上均强于其反交组合。所有正反杂交的杂种F1抗病性均超过其栽培种亲本。
3 讨论
芝麻野生种中具有很多栽培种缺乏的优异性状,尤其是抗病抗逆性,通过远缘杂交的方式将其优异基因转移到栽培种中,是创造种间遗传变异、丰富种质资源、改良芝麻抗性最有效的途径[5,14, 20-21]。由于芝麻种间杂交存在严重的生殖隔离、杂种胚败育等问题[14, 22],对种间杂种后代性状改良及利用研究的报道不多。本研究结果显示,在自然状态下野芝3号(S. indicatum)与栽培种杂交存在受精后的幼胚发育障碍,杂种幼胚发生败育。利用幼胚培养拯救技术显著提高获得种间杂交后代成功率,这也是开展远缘杂交普遍采用的方法[23-24]。在幼胚培养条件下,种间杂交组合正反交杂种胚的发育程度也存在很大的差异。染色体多的野生种作母本与栽培品种杂交其杂种胚更容易成苗。此外,研究还发现,将染色体少的栽培种预先加倍,使得染色体数目接近野生种的染色体数目,也可以提高F1幼胚成苗率,这与张海洋等[15]的研究报道一致。由SSR分子鉴定结果可知,种间杂种F1都有野芝3号的特异性带型,说明后代存在野芝3号的遗传物质,为芝麻远缘杂交后代鉴定提供了依据。
本文所用亲本野芝3号(S. indicatum,2n=58)植株匍匐,分布于印度,S. prostratum是野生种,分布于印度、非洲,植株匍匐,不易受病虫害影响,抗旱性也强,系栽培种(n=13)×S. prostratum(n=16)后代加倍获得的双二倍体[25]。已有研究表明,多倍体可通过表观遗传变异产生新的表现型,且能够将这种改变通过有性生殖传递给后代,从而提高基因表达的多样性,且多倍体物种比其二倍体祖先更能适应环境的变化[26]。本研究中芝14同源四倍体对茎点枯病的抗性要强于中芝14,且与野芝3号杂交的后代植株在茎点枯病抗性上也要强于其他杂交组合。
本研究通过野芝3号分别与中芝14、中芝14同源四倍体正反杂交,利用幼胚培养技术获得了真杂种F1株系,通过人工接菌,鉴定出了高抗茎点枯病的杂种F1株系,正反杂种F1的茎点枯病抗性均介于两亲本之间且高于其栽培种亲本,说明芝麻茎点枯病抗性可能由多基因控制,野芝3号是抗性提高的重要供体。研究发现杂交后代的抗性水平受到双亲抗性的影响,选择抗性较好的双亲在培育抗茎点枯病中起着重要的作用。
4 结论
通过幼胚培养技术成功获得芝麻栽培种与野生种的种间杂交F1株系,并通过形态学、细胞学和分子标记等方法得到证实。杂种F1的茎点枯病抗性介于双亲之间,但明显强于其栽培种亲本。The authors have declared that no competing interests exist.
