0 引言
【研究意义】玉米株高、穗位高、总叶片数等农艺性状不仅是决定田间种植密度和抗倒伏性的重要因素,而且影响着玉米籽粒产量的形成,是玉米育种中重要的改良目标性状。这三个性状具有较高的杂种优势,常被作为研究杂种优势遗传机理的主要农艺性状。因此,鉴定玉米株高、穗位高等性状的杂种优势基因位点不仅可以为杂种优势遗传机理研究提供材料基础,而且可以为选育耐密、抗倒优良玉米杂交种提供重要的理论基础。【前人研究进展】杂种优势是农作物育种中广泛应用的一种生物学现象,杂种优势的利用对提高玉米、水稻、高粱、油菜等农作物产量,增强抗逆性和稳产性具有重要的意义[1-3]。但杂种优势是生物体的整体表现,是多个性状相互作用与基因间互作的综合结果,这决定了杂种优势遗传机理研究的复杂性和难度。为了解释这个复杂的现象,前人提出了显性、超显性和上位性等假说[4-8]。随着分子标记的出现和应用,研究者借助分子标记和不同的分离群体对植物的杂种优势遗传机制进行了分析[9-12]。单片段代换系与非轮回亲本构建的测交群体,由于与非轮回亲本之间只存在一个或几个染色体片段之间的差异,可以直接鉴定出杂种优势位点所在的染色体片段,是杂种优势基因位点定位的理想群体,该类型群体已被用于番茄[13-14]、棉花[15]、拟南芥[16]、水稻[17]等杂种优势位点的鉴定。玉米是经典遗传学和杂种优势研究的模式植物[18-19],研究者对玉米株高、穗位高20]、产量及其构成因子[9,21]等性状的杂种优势遗传机理进行了一定数量的研究,为揭示杂种优势形成的遗传机理提供了研究基础。【本研究切入点】虽然前人利用不同的分离群体对植物杂种优势现象进行了研究,但是受限于这些群体复杂的遗传背景,玉米杂种优势遗传机理还不清楚,有待进一步研究。【拟解决的关键问题】本研究以前期构建的一套以优良玉米自交系许178为背景的综3单片段代换系为基础,对单片段代换系与许178的测交群体的株高、穗位高等主要农艺性状的杂种优势位点进行了鉴定,以期为玉米株高等农艺性状杂种优势基因克隆与遗传机理剖析提供理论依据。1 材料与方法
1.1 试验材料
利用203个玉米自交系许178背景的综3单片段代换系(SSSLs)群体为基础材料。该群体是以综3为供体亲本、许178为受体亲本,通过多代回交和自交结合SSR分子标记辅助选择,在BC4F2、BC4F3和BC4F4筛选到的纯合单片段代换系材料,包含150个不同染色体片段的SSSLs,分布在玉米的10条染色体上,代换片段平均长度为47.67 cM,导入片段总长11 394.22 cM,覆盖率玉米基因组的67.56%[21]。1.2 田间试验设计
2013年冬在海南以203个SSSLs为母本,受体许178为父本,利用人工授粉的方式获得203个SSSLs的测交群体。2014年夏播在鹤壁市农业科学院(浚县)、新乡市农业科学院(新乡)和许昌长葛对203单片段代换系及其回交群体进行了田间鉴定。田间试验采取随机区组设计,3次重复,单行区,行长4 m,行距0.6 m,密度为67 500株/hm2。为保证试验数据的准确性,SSSLs群体和2个亲本采用同样的田间设计播种在同一试验田中,田间管理措施同常规大田生产。1.3 性状田间测定
从苗期开始,每一行从第3株开始在不同的发育时期选择10株标记叶片数。散粉后选择10株表型一致的单株进行田间农艺性状调查,主要包括株高(plant height,PH)、穗位高(ear height,EH)、总叶片数(leaf number,LN)。株高指植株从地面到雄穗顶端的高度(cm);穗位高指植株从地面到第一个穗所在节的高度(cm)。1.4 QTL和杂种优势位点分析
采用SPSS17.0统计软件,对203个SSSLs群体所调查的性状进行单因素的方差分析和Duncan’s多重比较。如果某一性状与许178在P<0.05水平上差异显著,即认为该单片段代换系代换片段上存在一个目标性状的QTL[22]。加性效应值=(单片段代换系的表型值-对照的表型值)/2;贡献率=(加性效应值/对照的表型值)×100%。利用3个地点的中亲优势值(HMP)来检测杂种优势位点(heterotic loci,HL)[11]。中亲优势值计算公式如下:HMP(%)=(F1 -MP)/MP×100%。其中,MP= (单片段代换系的表型值+对照许178的表型值)/2;F1指对应测交群体的表型值。如果测交群体的某一性状表型值与对应MP在P<0.05上差异显著,即认为该测交群体对应的单片段代换系代换片段上存在一个杂种优势位点(heterotic loci,HL)[21]。
2 结果
2.1 单片段代换系及其测交群体主要农艺性状和杂种优势表现
2.1.1 单片段代换系及其测交群体的主要农艺性状表现 SSSLs群体在浚县、新乡和许昌点的株高平均值分别为216.07、201.22和183.65 cm(表1),且3个试点的变异与亲本相比,都比低值亲本综3低,比高值亲本许178高。穗位高的平均值分别为91.37、82.85和76.19 cm,在3个试点间的高度变化趋势与株高相同;与株高的变异不同,穗位高在浚县和许昌的最低高度未低于低值亲本。与株高和穗位高相比,总叶片数在亲本间和SSSL群体间的变异都较小。SSSLs群体的3个主要农艺性状在3个环境中与轮回亲本许178的表型值基本相同。受体亲本许178的株高和穗位高在所有测试点均比供体亲本综3高,叶片数则比综3少。Table 1
表1
表1SSSLs及其测交群体的主要农艺性状的表现
Table 1Performance of main morphological traits in SSSLs population and its testcross population
| 性状 Trait | 地点 Environ. | 许178 Xu178 | 综3 Zong3 | SSSLs群体 SSSLs population | 测交群体 Test-cross population | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 平均值 Mean | 平均值 Mean | 平均值 Mean ± SD | 变异范围 Range | 偏斜度Ske. | 峰度 Kur. | 平均值 Mean ± SD | 变异范围 Range | 偏斜度 Ske. | 峰度 Kurt. | |||
| 株高 PH (cm) | 浚县Xunxian | 219.33 | 192.07 | 216.07±14.12 | 155.33—247.93 | -0.740 | 1.629 | 227.92±7.48 | 204.27—246.60 | -0.199 | 0.024 | |
| 新乡Xinxiang | 208.13 | 190.33 | 201.22±13.08 | 152.00—227.73 | -0.745 | 1.045 | 211.98±7.17 | 192.93—230.87 | -0.159 | 0.029 | ||
| 许昌Xuchang | 184.13 | 176.70 | 183.65±11.58 | 131.61—208.93 | -0.813 | 1.927 | 182.97±10.21 | 153.60—218.00 | 0.179 | 0.712 | ||
| 穗位高 EH (cm) | 浚县Xunxian | 92.60 | 57.27 | 91.37±7.50 | 58.20—110.70 | -0.740 | 2.328 | 97.49±4.16 | 84.80—107.20 | -0.294 | -0.096 | |
| 新乡Xinxiang | 82.87 | 74.07 | 82.85±6.16 | 60.33—97.27 | -0.732 | 1.584 | 89.24±4.06 | 75.33—101.13 | -0.176 | 0.188 | ||
| 许昌Xuchang | 79.00 | 54.03 | 76.19±5.86 | 55.00—88.40 | -0.907 | 1.326 | 71.71±5.96 | 50.00—87.50 | -0.221 | 0.827 | ||
| 总叶片数 LN | 浚县Xunxian | 17.70 | 18.33 | 18.21±0.53 | 16.50—19.73 | 0.216 | 0.437 | 18.16±0.32 | 16.80—19.00 | -0.519 | 2.654 | |
| 新乡Xinxiang | 18.00 | 19.03 | 18.32±0.47 | 17.15—19.72 | 0.145 | 0.194 | 18.32±0.29 | 17.82—19.40 | 0.795 | 0.516 | ||
| 许昌Xuchang | 18.75 | 17.8 | 18.36±0.47 | 16.49—19.64 | -0.505 | 1.810 | 18.07±0.57 | 16.38—20.00 | 0.125 | 1.214 | ||
新窗口打开
SSSLs测交群体在浚县、新乡和许昌点的株高平均值分别为227.92、211.98和182.97 cm;穗位高平均值分别为97.49、89.24和71.71 cm(表1)。除许昌点外,测交群体株高和穗位高的平均值均高于对应的SSSLs群体和轮回亲本许178。测交群体总叶片数的平均值与SSSLs群体和许178的平均值相当。株高、穗位高、总叶片数的广义遗传率分别为65.0%、63.7%和80.8%。
2.1.2 单片段代换系测交群体主要农艺性状的中亲优势表现 单片段代换系测交群体在浚县、新乡和许昌点株高的中亲优势值分别为4.74%、3.61%和1.09%,变异范围分别为-3.27%—14.88%、-6.07%—13.28%和-4.99—10.92%(表2)。穗位高的中亲优势值分别为6.06%、7.77%和7.51%,变异范围分别为-7.95%—20.25%、-4.96%—24.30%和-13.24%—29.58%。总叶片数的中亲优势值分别为1.14%、0.90%和2.17%。不同试点中亲优势值和变异的数据表明,株高、穗位高的中亲优势值受环境影响较大,总叶片数中亲优势值也受到环境影响,但影响效应比株高和穗位高的小。
Table 2
表2
表2测交群体主要农艺性状的中亲优势表现
Table 2Mid-parent heterosis of main morphological traits in the SSSLs testcross population
| 性状Trait | 地点Environment | 平均值Mean ± SD | 变异范围Range | 偏斜度Ske. | 峰度Kur. |
|---|---|---|---|---|---|
| 株高 PH (%) | 浚县Xunxian | 4.74 ± 2.88 | -3.27—14.88 | 0.339 | 1.207 |
| 新乡Xinxiang | 3.61 ± 3.13 | -6.07—13.28 | 0.339 | 0.891 | |
| 许昌Xuchang | 1.09 ± 3.86 | -4.99—10.92 | 0.492 | -0.492 | |
| 穗位高 EH (%) | 浚县Xunxian | 6.06 ± 4.28 | -7.95—20.25 | 0.058 | 1.063 |
| 新乡Xinxiang | 7.77 ± 4.77 | -4.96—24.30 | 0.138 | 0.247 | |
| 许昌Xuchang | 7.51 ± 7.78 | -13.24—29.58 | 0.150 | 0.328 | |
| 总叶片数 LN (%) | 浚县Xunxian | 1.14 ± 1.68 | -5.35—5.42 | -0.523 | 1.765 |
| 新乡Xinxiang | 0.90 ± 1.30 | -2.15—5.81 | 0.435 | 0.760 | |
| 许昌Xuchang | 2.17 ±2.74 | -7.43—7.22 | 0.452 | 0.599 |
新窗口打开
2.2 玉米主要农艺性状的QTL定位
在3个地点共检测出9个株高的QTL,其中,qPH1b和qPH3b在3个环境中同时被检测到(表3)。qPH1b在浚县、新乡和许昌点对株高的贡献率分别为7.46%、10.11%和5.87%;qPH3b对株高的贡献率分别为8.31%、9.00%和6.97%,均由来自于综3的等位基因起增效作用。qPH10仅在一个环境中检测到,或为环境特异的QTL,而其余6个株高QTL可在2个环境中检测到。穗位高共检测到10个QTL,分布在除第6、7、9以外其他染色体上,有2个QTL(qEH3a和qEH5)在3个环境中同时被检测到。qEH3a在浚县、新乡和许昌点对穗位高的贡献率分别为19.00%、13.60%和13.90%;而qEH5在3个地点对穗位高的贡献率分别为7.20%、9.98%和6.49%。在检测到的株高和穗位高QTL中,qPH3b和qEH3c、qPH5和qEH5分别受相同片段umc1170、bnlg1006—umc1478—umc1587上的基因控制。总叶片数共检测到5个QTL,分别位于第1、5和10染色体上,只有qLN5在浚县和新乡2个环境中同时被检测,对玉米总叶片数的贡献率分别为5.74%和4.17%,可使总叶片数分别增加1.02和0.75个。2.3 玉米主要农艺性状的杂种优势位点分析
利用SSSL测交群体的表现值及其中亲优势进行显著性分析和杂种优势位点分析,在P<0.05的显著水平上株高性状共检测到6个HL,有3个HL在3个地点间被同时检测到,其中hPH1a在浚县、新乡和许昌点对株高中亲优势的贡献率分别为7.8%、7.8%和9.5%,表现为超显性效应(表4)。hPH3在浚县、新乡和许昌点对株高中亲优势的贡献率分别为8.9%、11.9%和7.8%,表现为超显性效应;而hPH8对株高中亲优势的贡献率分别为5.6%、5.3%和3.4%,表现为显性效应。穗位高共检测到8个HL,分布在除第5、6、9、10外的其余6条染色体上,只有hEH3在3个环境中同时被检测到,在浚县、新乡和许昌对穗位高中亲优势的贡献率分别为9.9%、12.3%和5.1%,表现为显性效应。
Table 3
表3
表3在SSSL群体中检测到的玉米主要农艺性状QTL
Table 3QTL identified for main morphological traits of maize in the SSSLs population
| 性状Trait | QTL | Bin | 位点 Position (cM) | 代换片段 Substitution segment | 加性效应值The additive effect | 贡献率 Contribution (%) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 浚县Xunxian | 新乡Xinxiang | 许昌Xuchang | 浚县Xunxian | 新乡Xinxiang | 许昌Xuchang | ||||||
| 株高PH | qPH1a | 1.01 | 79.25—190.53 | umc1106-umc1269-phi427913 -bnlg1007 | -15.11 | -14.12 | 7.26 | 7.67 | |||
| qPH1b | 1.06 | 457.00—532.80 | umc1703-umc1754-bnlg1598 | -16.36 | -21.04 | -10.81 | 7.46 | 10.11 | 5.87 | ||
| qPH3a | 3.02 | 48.44—103.30 | umc2377-bnlg1144-bnlg1647 | -12.51 | -14.03 | 6.01 | 7.62 | ||||
| qPH3b | 3.09 | 34.20—116.10 | umc1170 | -18.23 | -18.73 | -12.83 | 8.31 | 9.00 | 6.97 | ||
| qPH5 | 5.01 | 47.50—156.90 | bnlg1006-umc1478-umc1587 | -24.31 | -9.89 | 11.68 | 5.37 | ||||
| qPH6 | 6.05 | 244.75—335.45 | umc1614-umc1020-umc1462 | -13.27 | -14.34 | 6.05 | 6.89 | ||||
| qPH8 | 8.06 | 412.90—571.50 | umc2356-umc1724-phi080 | 14.30 | 9.30 | 6.52 | 5.05 | ||||
| qPH9 | 9.04 | 317.08—321.60 | umc1771-umc1990-umc1519 | -26.21 | -16.24 | 11.95 | 8.82 | ||||
| qPH10 | 10.04 | 272.20—344.80 | umc1053-umc2350-umc1506 | -19.44 | 9.34 | ||||||
| 穗位高EH | qEH1 | 1.01 | 86.30—190.53 | umc1269-phi427913-bnlg1007 | 7.04 | 6.00 | 7.60 | 7.24 | |||
| qEH2 | 2.04 | 250.10—320.70 | umc1024-umc1541-umc1579- phi083-bnlg1018-umc2079 | -6.70 | -6.64 | 8.09 | 8.40 | ||||
| qEH3a | 3.02 | 77.00—165.00 | bnlg1144-bnlg1647-umc1425 | -17.59 | -11.27 | -10.98 | 19.00 | 13.60 | 13.90 | ||
| qEH3b | 3.02 | 48.44—103.30 | umc2377-bnlg1144-bnlg1647 | -11.11 | -6.94 | 12.00 | 8.78 | ||||
| qEH3c | 3.09 | 34.20—116.10 | umc1170 | -8.22 | 10.40 | ||||||
| qEH4 | 4.11 | 599.63—731.60 | umc1989-umc1109-phi076 | -9.40 | 11.90 | ||||||
| qEH5 | 5.01 | 47.50—156.90 | bnlg1006-umc1478-umc1587 | -6.67 | -8.27 | -5.13 | 7.20 | 9.98 | 6.49 | ||
| qEH8 | 8.03 | 244.90—274.90 | umc2075-bnlg1863-phi100175 | 6.37 | 6.88 | ||||||
| qEH10a | 10.04 | 253.30—283.50 | umc1077-umc1053-umc2350 | 7.27 | 4.17 | 7.85 | 5.03 | ||||
| qEH10b | 10.05 | 283.50—380.50 | umc2350-umc1506-bnlg1028 | 9.05 | 6.26 | 9.77 | 7.56 | ||||
| 总叶片数LN | qLN1a | 1.05 | 439.11—457.00 | umc1689-umc1124-umc1703 | 0.93 | 5.23 | |||||
| qLN1b | 1.08 | 722.40—842.30 | umc1013-dupssr12-umc2047 | -0.86 | 4.56 | ||||||
| qLN5 | 5.06 | 156.90—493.50 | umc1587-mmc0081-umc2072- umc2305-umc1680 | 1.02 | 0.75 | 5.74 | 4.17 | ||||
| qLN10a | 10.04 | 253.30—283.50 | umc1077-umc1053-umc2350 | 0.86 | 4.77 | ||||||
| qLN10b | 10.05 | 283.50—380.50 | umc2350-umc1506-bnlg1028 | 0.89 | 5.01 | ||||||
新窗口打开
Table 4
表4
表4在SSSLs测交群体中检测到的主要农艺性状杂种优势位点
Table 4Heterotic loci identified for morphological traits in the SSSLs testcross population
| 性状 Trait | 杂种优势位点Heterotic loci | Bin | 位点 Position (cM) | 代换片段 Substitution segment | HMP(%; Effect) | ||
|---|---|---|---|---|---|---|---|
| 浚县 Xunxian | 新乡 Xinxiang | 许昌 Xuchang | |||||
| 株高 PH | hPH1a | 1.05 | 439.11—457.00 | umc1689-umc1124-umc1703 | 7.8 (OD) | 7.8 (OD) | 9.5 (OD) |
| hPH1b | 1.06 | 599.89—722.40 | umc1122-umc2396-umc1013 | 6.2 (D) | |||
| hPH2 | 2.04 | 274.90—320.70 | umc1541-umc1579-phi083- bnlg1018-umc2079 | 5.4 (D) | |||
| hPH3 | 3.02 | 77.00—165.00 | bnlg1144-bnlg1647-umc1425 | 8.9 (OD) | 11.9 (OD) | 7.8 (OD) | |
| hPH4 | 4.08 | 462.50—481.19 | bnlg1444-phi066-umc2041 | 9.5 (OD) | |||
| hPH8 | 8.06 | 412.90—571.50 | umc2356-umc1724-phi080 | 5.6 (D) | 5.3 (D) | 3.4 (D) | |
| 穗位高 EH | hEH1 | 1.04 | 287.20—398.20 | bnlg182-umc2217-bnlg2295 | 14.5 (OD) | ||
| hEH2 | 2.06 | 323.30—376.39 | umc1485-bnlg1831-bnlg1396 | 10.6 (D) | |||
| hEH3 | 3.02 | 77.00—165.00 | bnlg1144-bnlg1647-umc1425 | 9.9 (D) | 12.3 (D) | 5.1 (D) | |
| hEH4 | 4.08 | 462.50—481.19 | bnlg1444-phi066-umc2041 | 21.8 (OD) | |||
| hEH7 | 7.02 | 264.83—302.88 | umc1585-bnlg1808-bnlg1305 | 22.8 (OD) | |||
| hEH8a | 8.03 | 244.90—274.90 | umc2075-bnlg1863-phi100175 | 6.5 (D) | |||
| hEH8b | 8.05 | 274.90—412.90 | bnlg2046-bnlg162-umc1889-umc2401 | 10.2 (OD) | |||
| hEH8c | 8.06 | 412.90—571.50 | umc1724 | 15.7 (OD) | |||
| 总叶片数 LN | hLN1 | 1.05 | 439.11—457.00 | umc1689-umc1124-umc1703 | 1.7 (D) | 1.8 (D) | 10.0 (D) |
| hLN4a | 4.08 | 481.19—599.63 | umc2041-umc2365-umc1989 | 9.1 (OD) | |||
| hLN4b | 4.11 | 687.80—737.80 | umc1109-phi076-umc1058 | 5.1 (OD) | |||
| hLN5 | 5.06 | 156.90—493.50 | umc1587-mmc0081-umc2072- umc2305-umc1680 | 2.6 (D) | |||
| hLN9 | 9.02 | 24.30—131.10 | umc2078-umc1170 | 5.4 (OD) | |||
新窗口打开
总叶片数检测到5个HL,其中位于1.05 bin的hLN1在浚县、新乡和许昌点的中亲优势表型贡献率分别为1.7%、1.8%和10.0%,表现为显性效应。
在所有检测到的HL中,hPH3和hEH3受同一片段(bnlg1144-bnlg1647-umc1425)上杂种优势基因控制;hPH4和hEH4受同一片段(bnlg1444-phi066- umc2041)上杂种优势基因影响;而片段umc1689- umc1124-umc1703上的基因则同时影响株高和总叶片数杂种优势。
2.4 检测到的QTL位点和杂种优势位点在染色体上的分布
检测获得的QTL位点和杂种优势位点在玉米10条染色体上的分布极不均匀(图1)。其中,玉米第1条染色体上检测到的QTL位点和HL位点最多,分别为5个和4个;其次为第3、8、4、10染色体,检测到的QTL位点和HL位点总数都在5个以上;而第9染色体上只检测到1个株高QTL位点和1个叶片数HL。另外在第7染色体上只检测到1个穗位高HL,没有检测到QTL位点。而在第10染色体上只检测到5个QTL位点,没有检测到HL位点。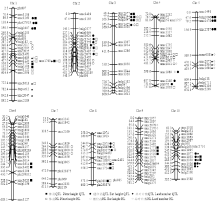 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1主要植株性状QTL和杂种优势位点在玉米染色体上的位置
-->Fig. 1Chromosomal location of QTL and heterotic loci (HL) for plant traits
-->
3 讨论
3.1 SSSL及其测交群体在重要性状遗传研究中的应用
不同类型的遗传分离群体在生物数量性状研究中发挥了重要作用。利用F2:3群体、DH群体、RIL群体、DH或RIL的测交群体、以及永久F2群体,研究者在作物中鉴定出多个重要农艺和产量性状的QTL和HL,揭示了重要农艺和产量性状的遗传基础,也为研究其生物学功能奠定了良好的基础。由于初级分离群体遗传背景复杂,对后续候选基因的克隆与功能分析造成困扰。为简化分离群体的遗传背景,ESHED等[23]通过回交与分子标记辅助选择的方法构建了一套番茄的单片段代换系群体,由于单片段代换系与受体亲本只存在一个供体染色体片段,在单片段代换系与受体亲本间鉴定出的QTL,可以直接对候选基因进行克隆。目前单片段代换系群体已经在小麦[24]、水稻[25]、玉米[26]等重要性状遗传机理研究中得到了广泛应用;同时通过片段的打断和将片段置于不同的遗传背景下,可以检测原先材料中被不利基因掩盖的有利基因或原背景下因互作受抑制的基因,如本研究中所检测到的9个株高QTL中,8个来自低值亲本综3,1个来自高值亲本许178,其原因即为此。单片段代换系的测交群体在分析方法上与单片段代换系群体相同,可以直接定位目标性状的杂种优势位点,近几年,被用于番茄[13-14]、棉花[15]、水稻[17]等作物的杂种优势研究中。在玉米中,利用SSSL的测交群体,对产量及产量构成因素[27];玉米籽粒性状[28-29],玉米株型相关性状[30]的杂种优势位点也进行了探究。本研究进一步分析了玉米的形态性状如株高、穗位高和总叶片数的杂种优势位点,共鉴定到19个HL。
利用SSSL测交群体的初级分析法只能检测到显性和超显性效应的HL位点,而无法检测位点之间的上位性,这是由SSSL测交群体自身单一背景的属性所决定的。在后续试验设计中,可利用已鉴定的HL位点,有目的的进行聚合从而可研究不同HL位点间的上位性作用,或证实不同HL位点效应的累加作用。
3.2 显性和超显性是株高、穗位高和总叶片数杂种优势的主要遗传基础
杂种优势是保证作物杂交种产量的一个重要遗传因素,其遗传基础得到广泛研究,并提出显性假说、超显性假说和上位性假说。目前,每一种假说都有分子遗传证据的支撑。然而,为不同作物或同一作物的不同性状或实验设计提炼出一个普适的杂种优势遗传理论依然是一项艰巨的挑战。番茄中利用SSSL探索杂种优势性状的遗传基础,结果表明特定染色体片段上超显性效应与产量相关性状相关,由此推测超显性QTL是利用杂种优势进行人工选择提高产量过程中被驯化[31]。拟南芥中,超显性或上位性是生物量相关性状杂种优势的主要作用类型[32]。在单位点水平上,玉米株高杂种优势位点表现以超显性效应为主[33],与本研究结论一致;除超显性效应外,显性效应对株高的杂种优势也起着重要的作用。尽管穗位高与株高的遗传基础相似[34],但两者杂种优势的位点和位点的主要效应却不尽相同。不同于株高,在所检测到的穗位高杂种优势位点中,多数位点表现为显性效应。玉米总叶片数与玉米开花期密切相关[35],然而关于玉米总叶片数遗传基础的研究报道较少。利用玉米近缘种大刍草与W22构建的BC2S3群体,LI等[36]发现玉米总叶片数由多数效应较小的QTL控制。本研究利用SSSL群体发现控制总叶片数的QTL的效应较小,与LI等[36]结论一致,但本研究中检测到的QTL数目较少,可能与构建群体的双亲间的差异较小有关。玉米叶片数变异较大[37],关于叶片数的杂种优势研究报道较少。本研究中首次从单位点水平剖析了玉米总叶片数杂种优势的遗传基础,发现其主要有显性和超显性效应为主。尽管叶片数性状也存在杂种优势,但其变异范围较小(表1),因此在杂交组合选育中,选择相近叶片数的亲本后可以忽略叶片数的变异而专注与其他性状的筛选。
由于株高和穗位高是决定玉米株型的重要农艺性状,而且两者均表现出较强的杂种优势,因此,在育种过程中可以利用分子标记辅助选择杂种优势弱的位点,从而选育出耐密抗倒的玉米新品种。
3.3 杂种优势QTL具有多效性
数量性状由多个基因控制,每个基因具有复杂的表达调控网络,可能影响植株发育的各个阶段,因而在QTL定位研究中经常发现一个QTL与多个性状相关。XUE等[38]在水稻中克隆的基因Ghd7,同时影响每穗粒数、株高和开花期。GUO等[39]在玉米中利用“永久F2”群体检测产量性状杂种优势的遗传基础时发现,杂种优势位点QTL并不孤立,它们或控制多个产量相关的性状,或存在相互作用。本研究中导入片段umc1689-umc1124-umc1703同时影响株高和总叶片数的杂种优势,表明杂种优势位点同样具有多效性现象。但是由于导入的片段中含有多个基因,因此不能排除这种单个片段上的“多效性”是由不同基因影响的可能,需要在后续试验设计中加以区分研究。3.4 杂种优势与环境条件的关系
数量性状易受环境条件影响,有时环境的影响甚至大于遗传效应本身,而且不同的性状受不同基因控制,对环境的响应亦表现不同。梁康迳[40]在水稻中的研究发现出了超显性、显性及互作外,环境会对不同性状的杂种优势产生作用,其中主穗粒数、主穗长、一次枝梗总长在两年的环境互作中表现正效应,一次和二次枝梗数则为负效应。本研究中检测到的HL位点中仅有4个HL在3个试点中同时检测到,其他HL位点受到地点环境影响较大。WEI等[30]利用相同的群体在2013年对株高、穗位高、总叶片数等性状进行了考察。与本研究结果比较发现,有4个HL位点在年份间可重复检测到,且其中3个HL位点在两年3个试点中共同检测到,其他HL位点则在年份间差异较大。因此,在玉米高产育种过程中,应有效减少一致性HL位点以维持一定株高和穗位高,从而保证杂交种产量;同时双亲遗传差异要适当,过大过小均不利于杂种优势水平的发挥。4 结论
单片段代换系测交群体的株高、穗位高中亲优势较大,总叶片数的中亲优势相对较小。定位到的24个QTL中,qPH1b、qPH3b、qEH3a和qEH5在不同环境间具有保守性,是所对应性状的主效QTL,其所处的染色体区域可能含有调控相关性状的主要基因。定位到的杂种优势位点,hPH1a、hPH3、hPH8、hEH3和hLN1具有环境保守性,这些主要的杂种优势位点分布区域可能含有调控相对应性状杂种优势的相关基因。株高、穗位高多数QTL/HL位点表现出超显性效应,而多数总叶片数相关的QTL/HL显示出显性效应,表明杂种优势主要来源于位点间的超显性效应。染色体片段umc1689-umc1124-umc1703同时影响株高和总叶片数的杂种优势,说明杂种优势同样存在多效性现象。株高和穗位高杂种优势与产量呈正相关,总叶片数多的品种具有更高的光合利用效率,上述主效QTL和HL可通过分子标记辅助选择,应用于理想株型育种。The authors have declared that no competing interests exist.

{kind=link}
{kind=link}