0 引言
【研究意义】漆酶(EC1.10.3.2)是一类在催化中心含有多个铜离子的多酚氧化酶,也被称为多铜氧化酶[1]。在真菌中,漆酶主要参与病菌与寄主间的互作。以玉米大斑病菌(Setosphaeria turcica)漆酶基因Stlac2为出发点,探索其生物学特性,可为研究其在玉米大斑病菌致病过程中的作用打下基础,对病害防治具有重要意义。【前人研究进展】根据铜离子的配位清空和光谱特性,可将漆酶分为3种类型(类型I、类型II、类型III)[2]。漆酶的I型铜原子和氨基酸残基结合成为单核中心,II型和III型铜原子构成3核中心,I型铜原子作为初级电子的接受者,它参与分子内的电子传递,把电子从底物传递到其他铜原子上;之后电子结合于3核位点,该位点进一步把电子传递给活性中心的第二底物氧分子,使之还原为水[3]。漆酶广泛存在于高等植物和真菌中,在昆虫[4]和细菌中也有发现[5-8]。在高等植物中,漆酶在木质素形成过程中起到了主要作用[9-13]。而在真菌研究中发现,漆酶主要参与病菌的致病性、形态建成和木质素降解等过程[14];如Cryphonectria parasitica漆酶基因的缺失导致其毒力和致病性较野生型菌株均有所下降;同样,Sinorhizobium meliloti 中漆酶基因的缺失导致其抗逆性和致病性降低。漆酶能够氧化酚、多酚、芳香胺和一些非酚类化合物,将底物分子上的电子转移到氧分子上,从而使氧还原成水[15]。自然条件下伴随着木质素的降解过程会产生一些小分子物质,例如香草醛、4-羟基苯甲醛、丁香醛、香草酸、4-羟基苯甲酸和香兰素等[16-17],这些小分子物质严重影响了木质素的后续降解过程[18-19]。漆酶能够氧化这些小分子物质,同样在这些物质存在的条件下,也能诱导漆酶的活性及产量的增加,如香草酸可影响真菌漆酶的活性及产量[20],香兰素和丁香酸对Trametes sp. AH28-2漆酶活性有明显的诱导作用[21],阿魏酸和香草醛可使Plemoutus pulmonarius漆酶产量提高10倍[22],4-羟基苯甲酸对Volvariella volvacea漆酶基因lac1有显著的诱导作用[23],香草酸和壬基酚对Clacariopsis aquatica漆酶基因lacc4有显著诱导作用,添加1 mmol·L-1香草酸的样品在第10天时漆酶基因的表达量是对照的7倍左右[24]。玉米大斑病是由玉米大斑病菌引起的一种真菌病害,在世界各个玉米产区危害严重[25-26]。曹志艳[27]通过RACE技术从玉米大斑病菌基因组中克隆到Stlac2,曹可可[28]通过多铜氧化酶结合铜离子的保守结构域比对玉米大斑病菌基因组,预测到了9个具有典型铜离子结合结构域的类漆酶基因,其中EOA90070与曹志艳克隆到的Stlac2基因序列一致,并将其命名为Stlac2,根据表达的蛋白质序列与已报道的多个真菌漆酶进行比对分析,发现 EOA90070(Stlac2)与小麦黄斑病菌(Pyrenophora tritici- repentis)等致病漆酶同源性较高,属于狭义的子囊菌漆酶,推测该基因在玉米大斑病菌侵染寄主过程中起重要作用。詹旭等[29]研究发现,玉米大斑病菌、玉米小斑病菌(Cochliobolus heterostrophus)、玉米弯孢叶斑病菌(Curvularia lunata)等10个产漆酶的植物病原真菌均具有降解木质素的能力,其中玉米大斑病菌产漆酶活性最高,为18.984 U·mL-1。曹可可等[30]建立了以漆酶活性为响应值的多元二次回归模型,对玉米大斑病菌发酵条件进行优化,最佳条件下漆酶活性最高达(40.00±1.20)U·mL-1。【本研究切入点】在笔者实验室前人研究的基础上,进一步探究玉米大斑病菌漆酶基因Stlac2的结构及其蛋白活性,筛选确定影响该基因表达的底物分子。【拟解决的关键问题】通过对Stlac2进行生物信息学分析,检验基因所属类别,通过分析其在不同小分子物质存在条件下的表达情况,研究Stlac2在降解木质素过程中起到的作用,并通过原核表达来探究其催化条件,为进一步研究基因及蛋白功能打下基础。1 材料与方法
试验于2015年10月至2016年8月在河北农业大学真菌毒素与植物分子病理学实验室完成。1.1 试验材料
玉米大斑病菌01-23由河北农业大学真菌毒素与植物分子病理学实验室保存。RNA提取试剂盒,TransStart Top Green qPCR SuperMix,E. coli BL21(DE3)购于北京全式金生物技术有限公司;限制性内切酶BamH I、Hind III、LA Taq DNA酶、T4 Ligase、PrimeScriptTM RT Reagent Kit with gDNA Eraser(Perfect Real Time)反转录试剂盒等购于TaKaRa公司。1.2 供试培养基及试剂
PDA培养基(g·L-1):马铃薯200,葡萄糖20,琼脂粉13,自然pH;PD培养基(g·L-1):马铃薯200,葡萄糖20。以上培养基均需经过1×105 Pa,121℃灭菌20 min后使用。ABTS购于生工生物工程(上海)股份有限公司。柠檬酸缓冲液(pH 3.0):0.1 mol·L-1柠檬酸18.6 mL,0.1 mol·L-1柠檬酸三钠1.4 mL。1.3 方法
1.3.1 Stlac2生物信息学分析 通过NCBI查找玉米大斑病菌Stlac2(EOA90070)与已知漆酶基因,如马尔尼菲青霉菌(Penicillium marneffei)PbrB(PMAA_082060)、烟曲霉(Aspergillus fumigatus)Abr2 (AFUA_2G17530)、构巢曲菌(Aspergillus nidulans)YA(AN6635)用ClustalX进行蛋白序列比对,查看Stlac2中的铜离子结合位点;利用在线软件SOPMA(http://nhjy.hzau.edu.cn/kech/swxxx/jakj/dianzi/ Bioinf7/Expasy/Expasy8.htm)和ProtParam(http://web. expasy.org/protparam/)对其二级结构及理化性质进行检测,最后用SWISS-MODEL(http://swissmodel. expasy.org/)对Stlac2蛋白的三维结构进行预测,并用SAVES(http://services.mbi.ucla.edu/SAVES/)对三维结构进行评价。1.3.2 玉米大斑病菌总RNA的提取 将生长在PDA培养基上4 d的玉米大斑病菌分别接种至含有0.01 g·L-1的香草醛、4-羟基苯甲醛、紫丁香醛、香草酸、4-羟基苯甲酸和丁香酸的PD培养基中,于25℃,黑暗静置培养7 d后,搜集菌丝,用TransZol Up试剂盒提取总RNA用于Stlac2的表达分析及cDNA克隆。
1.3.3 Stlac2在不同木质素降解产物中的表达分析 将1.3.2中提取的玉米大斑病菌总RNA根据PrimeScriptTM RT Reagent Kit with gDNA Eraser(Perfect Real Time)反转录试剂盒操作说明,将RNA反转录成cDNA。置于-80℃保存。根据Stlac2基因序列设计RT-qPCR引物,并以Tubulin作为内参。用Eppendorf Mastercyclyer ep realplex实时荧光定量PCR仪,对Stlac2在不同小分子物质存在下的表达情况进行检测,每组试验重复3次。引物序列见表1。
Table 1
表1
表1玉米大斑病菌漆酶表达分析引物
Table 1The primers used for RT-qPCR analysis
| 名称 Name | 序列 Sequence (5′-3′) | 大小 Size (bp) |
|---|---|---|
| RT-lac2-Fw | GGAACAACATCTCCGAGTAGCCACC | 170 |
| RT-lac2-Rev | CAACTGGGGTGATTGGATTCTGTGG | |
| Tubulin-Fw | GGGAACTCCTCACGGATGTTG | 156 |
| Tubulin-Rev | TAACAACTGGGCAAAGGGTCA |
新窗口打开
1.3.4 Stlac2的原核表达及活性检测 以pET-30a为框架,用Stlac2-F:5′- CGCGGATCCGCGATGTCTT ACAATG -3′,Stlac2-R:5′-CCCAAGCTTGGGCAGGC CCGAGTCG -3′(下划线为加入酶切位点BamH I和Hind III)扩增Stlac2 cDNA全序列,构建Stlac2原核表达载体pET-Stlac2,之后将表达载体转入BL21(DE3)E. coli感受态细胞中,挑取阳性克隆接种至含有50 μg·mL-1卡那霉素的LB培养基中,于37℃,220 r/min条件下培养,待其OD600达0.7—0.8,用1 mmol·L-1的IPTG诱导,2、4 h后分别取样1.5 mL,6 000 r/min室温离心2 min。用30 μL 5×SDS上样缓冲液重悬沉淀,沸水浴15 min,5 000 r/min室温离心1 min,取10 μL上清液进行聚丙烯酰胺凝胶电泳(12%分离胶,5%浓缩胶),检测蛋白表达情况。Stlac2蛋白的纯化步骤如下:(1)15℃诱导 12 h,收集细胞,并用140 mL冰浴的Buffer A(20 mmol·L-1 Tris-HCl,300 mmol·L-1 NaCl,1% Triton-100,pH 8.0)重悬,超声裂解细胞,离心后上清过Ni-IDA柱;(2)200 mL Buffer E(20 mmol·L-1 Tris-HCl pH 8.0,2 mol·L-1 NaCl,0.1% TritonX-100)洗涤 Ni-IDA 柱;(3)50 mL Buffer F(20 mmol·L-1 Tris-HCl pH 8.0,50 mmol·L-1 NaCl,0.1% TritonX-100,20 mmol·L-1咪唑)洗涤Ni-IDA柱;(4)用Buffer G(20 mmol·L-1 Tris-HCl pH 8.0,50 mmol·L-1 NaCl,0.1% TritonX-100,250 mmol·L-1咪唑)洗脱 Ni-IDA柱。
Stlac2蛋白的活性检测:漆酶活性的检测采用ABTS法。5 mL反应体系中含有100 mmol·L-1的柠檬酸缓冲液(pH 3.0),2 mmol·L-1的ABTS,50 μL的Stlac2纯化蛋白,于30℃反应5 min,测定420 nm下吸光度的变化值。每分钟使1 μmol ABTS 氧化所需要的酶量定义为一个酶活力单位。公式如下:
U·L-1=n×△A×106/36 000/5
其中,n为稀释倍数,△A为反应液5 min中内在420 nm处吸光度变化值,消光系数ξ为36 000 mol·L-1·cm-1。
2 结果
2.1 Stlac2生物信息
通过ClustalX软件对玉米大斑病菌漆酶基因Stlac2与已知漆酶基因进行序列比对,并对其不同类型的Cu离子结合位点进行分析(图1)。发现在Stlac2中的所有铜离子结合位点均与已知漆酶一致,说明Stlac2为典型的漆酶基因。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1Stlac2与已知漆酶基因的蛋白序列比对1、2、3 代表不同类型的铜离子结合位点,#代表可能的质子供给位点
-->Fig. 1The protein alignment of the Stlac2 with the known laccases1, 2, and 3 were different types of Cu ion binging site. The hash (#) indicated the potential proton donor for the reaction intermediates
-->
通过SOPMA在线软件对Stlac2蛋白的二级结构进行预测分析(图2),得出α-螺旋、延伸链、β-转角和无规则卷曲在554个氨基酸中所占的比例分别为18.59%、25.63%、11.91%和43.86%(表2)。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2 Stlac2 二级结构分析e:β-折叠 β-strand;c:无规则卷曲 Random coil;t:β-转角 β-turn;h:α-螺旋 α-helix
-->Fig. 2The secondary structure analysis of Stlac2
-->
Table 2
表2
表2漆酶基因二级结构组成分析
Table 2The laccases secondary structure compositon analysis
| 漆酶基因 Laccase gene | α-螺旋 α-helix (%) | 延伸链 Extended strand (%) | β-转角 β-turn (%) | 无规则卷曲 Random coil (%) |
|---|---|---|---|---|
| Arb2 | 18.91 | 28.45 | 8.52 | 44.12 |
| PbrB | 16.02 | 33.22 | 10.79 | 39.97 |
| YA | 20.20 | 28.74 | 12.32 | 38.75 |
| Stlac2 | 18.59 | 25.63 | 11.91 | 43.86 |
新窗口打开
通过在线软件ProtParam对其理化性质进行推测,发现Stlac2分子量为61.64 kD,等电点为5.00,不稳定系数为30.23,280 nm处吸光值介于1.627—1.635,总体疏水性(GRAVY)为-0.37,表明其为亲水蛋白(表3)。
Table 3
表3
表3漆酶基因理化性质分析
Table 3The biochemical properties analysis of laccases
| 漆酶基因 Laccase gene | 氨基酸数 Amino acid number | 分子量 Molecular weight (kD) | 等电点 pI | 不稳定系数 Instability index | 280 nm吸光值 Abs 280 nm | 平均疏水性 Grand average of hydropathicity |
|---|---|---|---|---|---|---|
| Arb2 | 587 | 65.29 | 4.99 | 33.88 | 1.789-1.792 | -0.329 |
| PbrB | 593 | 66.47 | 5.03 | 35.56 | 2.110-2.118 | -0.262 |
| YA | 609 | 67.98 | 4.54 | 33.77 | 1.775-1.779 | -0.172 |
| Stlac2 | 554 | 61.64 | 5.00 | 30.23 | 1.627-1.635 | -0.367 |
新窗口打开
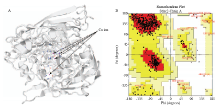
利用SWISS-MODEL对Stlac2蛋白质的三维结构进行预测(图3-A),并用SAVES在线软件对三维结构进行质量评估(图3-B)。核心区域的残基位点占83.2%,其他允许区域残基位点占16.1%,不合理残基位点占0.7%。3D-1D score≥0.2的残基位点占83.24%。表明Stlac2的三维构象比较合理。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3Stlac2 三维构象(A)和拉氏构象图(B)分析紫色圆球为铜离子
-->Fig. 3The three-dimentional (A) and Ramachandran Plot (B) analysis of Stlac2Fuchsia sphere represented Cu ion
-->
2.2 木质素降解产物存在下玉米大斑病菌胞外漆酶的产生情况
将玉米大斑病菌培养在含有0.01 g·L-1的木质素降解产物(丁香醛、4-羟基苯甲醛、香草酸、香兰素、4-羟基苯甲酸)的PDA中,并添加终浓度为0.3 mmol·L-1的ABTS和终浓度为30 mmol·L-1的KNO3,培养5 d后发现,与CK对比,菌落周围的紫色深浅依次为香草酸>香兰素>4-羟基苯甲醛>4-羟基苯甲酸>丁香醛>CK,表明在这些小分子有机物存在的条件下,玉米大斑病菌漆酶基因的表达量有所增加,其中在添加香草酸、香兰素、4-羟基苯甲醛时,表现尤其明显(图4)。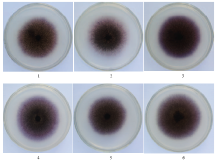 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4小分子物质存在条件下玉米大斑病菌胞外漆酶产生情况1:CK;2:丁香醛 Syringaldehyde;3:香草酸 Vanillic acid;4:香兰素 Vanillin;5:4-羟基苯甲酸 4-Hydroxybenzoic acid;6:4-羟基苯甲醛 4-Hydroxybenzaidehyde
-->Fig. 4The production of extracellular laccase under the small molecular organics
-->
2.3 木质素降解产物存在下Stlac2的相对表达分析
用TransZol Up提取试剂盒提取PD中培养7 d的玉米大斑病菌的总RNA,进行RT-qPCR试验,结果显示,在加入4-羟基苯甲酸和4-羟基苯甲醛后Stlac2的表达量显著升高,约为CK的5—8倍,而在加入香兰素时,Stlac2的相对表达量有所下降,在加入其他物质时表达量变化不显著(图5)。说明4-羟基苯甲酸和4-羟基苯甲醛能够诱导Stlac2的表达,在木质素降解过程中,Stlac2发挥了一定的作用。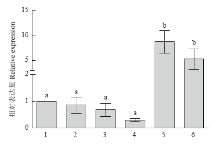 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5不同物质下Stlac2的相对表达量1:CK;2:丁香醛 Syringaldehyde;3:香草酸 Vanillic acid;4:香兰素 Vanillin;5:4-羟基苯甲酸 4-Hydroxybenzoic acid;6:4-羟基苯甲醛 4-Hydroxybenzaidehyde。图中数值为平均值±标准误,柱状图上不同的字母代表基因相对表达差异显著(P<0.05)
-->Fig. 5The relative expression of Stlac2 under different organicsThe values were mean±standard error, the different letters above the bar represented the significant differences (P<0.05)
-->
2.4 Stlac2的原核表达及活性测定
以pET-30a载体为框架,用Hind III和BamH I双酶切Stlac2片段连接到载体上,构建pET-Stlac2表达质粒(图6)。将成功构建的表达质粒转入大肠杆菌感受态细胞中,挑取阳性单克隆,然后提取质粒进行双酶切验证(图7)。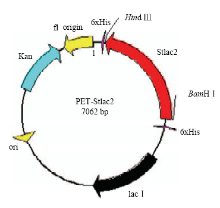 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图6pET-Stlac2 重组质粒图谱
-->Fig. 6Construction of the pET-Stlac2 expressed plasmid
-->
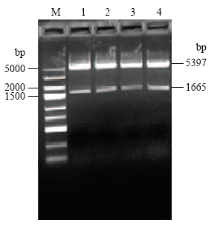 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图7pET-Stlac2质粒双酶切验证M:DL5000 DNA Maker;1—4:pET-Stlac2质粒 pET-Stlac2 plasmid
-->Fig. 7Double restriction enzyme digestion analysis of pET-Stlac2
-->
将双酶切验证正确的表达质粒转入BL21(DE3)感受态细胞中,加入终浓度为1 mmol·L-1的IPTG进行诱导表达,经SDS-PAGE检测后,结果显示,在未经IPTG诱导的菌液中,培养2 h和4 h后,重组质粒在67 kD(61.64 kD Stlac2 + 5.6 kD空载)左右未产生的明显的蛋白条带,而经IPTG诱导的菌液中,在67 kD左右有清晰的蛋白条带,证明Stlac2被成功表达(图8)。纯化后检测漆酶活性为(40.7± 0.3)U·L-1。
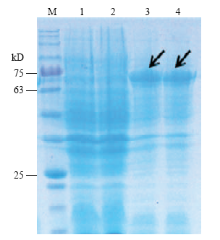 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图8诱导蛋白SDS-PAGE分析M:标准蛋白Marker Protein molecular weight marker;1:未诱导2 h Without IPTG induction for 2 h;2:未诱导4 h Without IPTG induction for 4 h;3:诱导2 h Cultivation with IPTG induction for 2 h;4:诱导4 h Cultivation with IPTG induction for 4 h。黑色箭头所指为目的条带
-->Fig. 8The analysis of SDS-PAGEThe black arrow indicated the obtained protein
-->
3 讨论
自1883年首次从漆树中发现漆酶以来,漆酶一直是从事化学和生物学等领域****所关注的热点话题。第一个被扩增出来的真菌漆酶为粗糙脉孢菌(Neurospora crassa)漆酶基因[31],之后更多的真菌漆酶被发现,特别是在以担子菌为主的白腐真菌中[32],随着研究深入及科学技术的发展,漆酶在真菌中的功能和作用越来越清楚,在病原性真菌中,漆酶在色素产生、子实体形成、菌体形态建成、病原菌致病性、植物与病原菌互作、脱毒和应激反应等方面具有重要作用[33-35]。在木质素降解过程中,漆酶是发现最早的能够降解木质素的酶类,而且不同于LiP(lignin peroxidase)、MnP(mangnase peroxidase),漆酶在氧化降解木质素的过程中不需要过氧化氢的参与[36-38]。在木质素的发酵降解过程中,会产生一类含量远低于碳水化合物,但却严重影响后续发酵的小分子芳香族化合物,例如香草醛、4-羟基苯甲醛、紫丁香醛、香草酸、4-羟基苯甲酸和紫丁香酸等[16-17]。由于漆酶氧化底物的广谱特性,使其在木质素降解过程中显得尤为重要。
研究表明,与木质素结构单元相似的芳香族小分子化合物或木质素降解过程中的中间产物,如香草酸等对真菌漆酶的活性及产量均有一定的影响。香兰素和丁香酸对Trametes sp. AH28-2漆酶的活性有明显的诱导作用[21],香草醛可使Plemoutus pulmonarius 漆酶产量提高10倍[22]。
近些年来,对病原性真菌漆酶在木质素降解中的作用研究的比较少,主要原因是在病原性真菌中,漆酶的产量及活性低于白腐真菌。本试验基于前人研究结果,在保证高产漆酶的条件下,在玉米大斑病菌中,根据与已知漆酶保守结构进行ClustalX蛋白序列比对,并利用Neighbour joining方法聚类发现EOA90070(Stlac2)与小麦黄斑病菌等致病漆酶同源性较高,属于典型的子囊菌漆酶。再通过生物信息学对Stlac2的生物学特性进行了预测与比对,判断其为漆酶基因。为研究其在木质素降解过程中是否起到了一定的作用,对其在小分子物质存在条件下的相对表达量进行了分析,发现其在4-羟基苯甲酸和4-羟基苯甲醛存在时表达量上升了5—8倍。已有研究表明,小分子物质诱导漆酶基因转录的机制可能与漆酶基因启动子上游的异生物质响应元件(xenobiotic response element,XRE)相关。XRE是真核生物中参与芳香族化合物激活某些特定基因转录的重要顺式作用元件[39-41],而在Stlac2中是否有此响应元件还有待研究。为了更深一步研究Stlac2的功能,本研究对该基因进行了克隆及原核表达,并成功表达出了相应大小的蛋白条带。表达出的目的蛋白在体外成功地检测出漆酶活性,这种真核漆酶基因原核表达的方式报道的很少,为后期研究Stlac2在真菌中的作用和功能打下了基础。
4 结论
生物信息学分析表明Stlac2具有漆酶基因的典型结构,为玉米大斑病菌漆酶基因;该基因在4-羟基苯甲醛和4-羟基苯甲酸存在下表达量显著提高,表明Stlac2在木质素降解过程中起到了一定作用;以pET-30a为表达载体框架,实现了玉米大斑病菌漆酶异源表达,胞外漆酶活性可达(40.7±0.3)U·L-1,该研究结果为后期研究蛋白性质打下了基础。The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}