 ,1,3, 刘萍1, 黄丹丹1, 贾淑霞1, 张晓珂2, 张士秀,1, 梁文举2, 陈学文1, 张延1, 梁爱珍1
,1,3, 刘萍1, 黄丹丹1, 贾淑霞1, 张晓珂2, 张士秀,1, 梁文举2, 陈学文1, 张延1, 梁爱珍1Response of Nematode Community to Soil Disturbance After Long-Term No-Tillage Practice in the Black Soil of Northeast China
ZHANG MengTing,1,3, LIU Ping1, HUANG DanDan1, JIA ShuXia1, ZHANG XiaoKe2, ZHANG ShiXiu,1, LIANG WenJu2, CHEN XueWen1, ZHANG Yan1, LIANG AiZhen1通讯作者:
责任编辑: 李云霞
收稿日期:2020-11-25接受日期:2021-05-6
| 基金资助: |
Received:2020-11-25Accepted:2021-05-6
作者简介 About authors
张梦亭,Tel:0431-85542234;E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (1694KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
张梦亭, 刘萍, 黄丹丹, 贾淑霞, 张晓珂, 张士秀, 梁文举, 陈学文, 张延, 梁爱珍. 东北黑土线虫群落对长期免耕后土壤扰动的响应. 中国农业科学, 2021, 54(22): 4840-4850 doi:10.3864/j.issn.0578-1752.2021.22.011
ZHANG MengTing, LIU Ping, HUANG DanDan, JIA ShuXia, ZHANG XiaoKe, ZHANG ShiXiu, LIANG WenJu, CHEN XueWen, ZHANG Yan, LIANG AiZhen.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】免耕是指在作物春播时不扰动土壤,并且在作物收获后用残茬覆盖至少30%土壤表面的耕作方式[1,2]。与传统耕作相比,免耕不仅减少了耕作频率而且增加了有机物料还田,因此免耕在防止土壤侵蚀、提高土壤生产力及维持土壤健康等方面具有重要的积极作用。然而这些积极效应的实现,尤其是提高或稳定作物产量,需要长期连续地实施免耕耕作方式。一般来说,当连续实施免耕大于5年后,土壤环境会达到一个新的平衡点,此时土壤的物理、化学和生物学特性都达到相对稳定的状态[3,4]。最近有些研究指出[5,6]在长期连续实施免耕后会造成耕层土壤板结的现象,并提出进行适当的土壤扰动,如翻耕土壤,可以打破土壤板结,有效改善土壤环境水、肥、气、热的状况。然而,目前对长期免耕土壤实施扰动后,土壤生态系统稳定性的响应情况还尚不了解。与土壤物理、化学特性相比,土壤生物学特性更能对外界的扰动做出灵敏迅速的响应。在种类繁多的土壤生物中,土壤线虫是地球上最为丰富的后生动物,也是土壤生物群落中的优势组分,约占陆地所有动物数量的80%[7]。土壤线虫占据了土壤食物网中的多个营养级,在维持土壤生态系统的稳定性、促进物质循环和能量流动等方面发挥了重要的生态功能,因而土壤线虫常被广泛地用作土壤生物活性的重要指示者来评估土壤的健康状况及生态系统的稳定性[8,9]。因此,本研究选择土壤线虫作为指示生物,研究土壤线虫群落结构对长期免耕扰动前和扰动后的响应情况,以为东北黑土区合理选择耕作措施提供科学依据和参考。【前人研究进展】耕作方式作为最常见的农田管理措施,会对土壤线虫群落产生重要影响。大量研究发现[10,11,12,13],与常规耕作相比,免耕不仅会促进线虫总多度的增加,还会提高食微线虫(包括食细菌线虫和食真菌线虫)及杂食/捕食线虫的多度。此外,免耕系统中的线虫生态指数,如多样性指数、成熟度、结构指数、通道指数和功能代谢足迹等,通常高于常规耕作系统,表明相较于常规耕作,土壤扰动相对较少的免耕系统中线虫的多样性程度和丰富程度更高,使土壤线虫微食物网的结构趋于稳定[14,15]。这主要是由于免耕有利于c-p值较高的线虫类群的生存发展。BONGERS[16]于1990年依据生活史对策将各科线虫划分为r-对策者向k-对策者过渡的5个不同的c-p(colonizer-persister)类群,c-p值越高,世代时间越长,对环境压力越敏感,可有效反映土壤生态系统受外界干扰的程度。为了获得作物的最优产量并维持农业生态系统的可持续发展,免耕与作物轮作常常配套实施[17]。与单一作物连作相比,作物轮作通过影响输入土壤中有机物料的质量,调节食物资源自下而上调控土壤线虫群落结构[18]。已有的研究表明,免耕玉米-大豆轮作下线虫多度及结构指数显著高于免耕玉米连作,说明免耕玉米-大豆轮作下土壤线虫微食物网更为复杂,且其结构更为稳定[19,20]。【本研究切入点】综上所述,可见关于土壤线虫群落结构对免耕及免耕轮作配施的响应情况已开展了大量研究,并得出了较为一致的结论,即:实施免耕有利于形成稳定的土壤线虫群落结构。然而,目前关于长期免耕扰动后对土壤线虫群落结构的影响还鲜有报道,这限制了我们对长期免耕扰动后土壤生态系统稳定性的认识。【拟解决的关键问题】依托中国科学院东北地理与农业生态研究所黑土农业试验示范平台,对长期(≥10年)实施免耕不同轮作方式下的土壤进行深翻和秸秆移除,探究土壤线虫群落结构对长期免耕扰动前和扰动后的响应情况。我们假设深翻和移除秸秆能够破坏长期免耕下线虫群落结构及其稳定性,但破坏程度因轮作方式而异。研究结果将为黑土可持续利用提供理论依据。1 材料与方法
1.1 试验地概况
试验地点位于吉林省德惠市米沙子乡(44°12′N,125°33′E)中国科学院东北地理与农业生态研究所黑土农业试验示范基地,海拔约为177 m。试验地属于温带大陆性季风气候,冬季寒冷漫长(11月至次年3月)且干旱,积雪覆盖一般低于0.25 m,季节性积雪覆盖通常始于11月,次年4月初融化。年平均气温4.4℃,年平均降水量520 mm,其中6、7、8月份降水量占70 %以上。土壤类型为中层典型黑土(Typic Hapludoll,美国农业部土壤分类法)[21],由36.0%的黏粒、24.5%的粉粒和39.5%的砂粒组成的黏质壤土。在试验建立之前,一直采用传统耕作方式种植玉米(>30年)[22]。1.2 试验设计
本试验具有两个阶段,均依托于同一试验区。阶段一:确定长期免耕对线虫群落结构的影响情况,以常规耕作样地为对照;阶段二:对长期免耕土壤进行一次扰动,包括土壤翻动整地及秸秆移除等措施,以评估长期免耕扰动后线虫群落结构的变化情况。因此,本研究具有两种长期免耕耕作模式,但需要注意的是阶段一的采样时间是在连续实施免耕10年后,阶段二的采样时间是在连续实施免耕15年进行一次土壤扰动。两次采样具有时间间隔,但本文关注重点不是相同耕作方式下时间序列上的比较,而是相同采样时间下不同耕作方式之间的比较。此外,在长期连续(>5年)实施免耕后土壤会达到新的平衡状态[3-4,23],形成相对稳定的土壤环境,因此免耕实施10年或15年都属于长期免耕阶段,可用来评估长期免耕扰动前或扰动后线虫群落结构相对常规耕作的变化情况。试验区于2001年秋采用单因素裂区、随机区组设计,4次重复。耕作方式作为主因素,轮作方式作为副因素。每个小区的面积是5.2 m×20 m。农作物于每年5月播种,10月收获。每次收获后都有一个休耕期(大约7个月)。耕作方式包括常规耕作(conventional tillage,CT)、垄作(ridge tillage,RT)和免耕(no tillage,NT),在此研究中只考虑CT和NT处理。轮作方式包括为玉米-大豆轮作(CS)和玉米连作(CC)。2017年春,即第15年作物收获后,对NT处理小区采取传统耕作方式进行深翻(约20 cm)整地,并将所有地上秸秆移除。扰动后的NT命名为DNT(disturbed no-tillage)。
各耕作方式的管理如下:CT,秋收后进行耕翻(深度约为20 cm),春季进行整地(深度约为7.5—10 cm)、播种、中耕和起垄作业;NT,除用播种机(KINZE- 3000,美国)直接播种外,全年不再扰动土壤;DNT,使用拖拉机和三角铧犁对长期免耕土壤进行翻耕,深度约为20 cm,春季按CT处理方式进行整地(深度约为7.5—10 cm)、播种、中耕和起垄等一次作业。作物收获后,CT处理下的所有秸秆全部移除;NT处理下,玉米留茬35 cm,并将长约30 cm的秸秆直接返还于土壤表面,大豆地上部秸秆直接覆盖于土壤表面;DNT处理下的所有秸秆用打捆机打捆后全部移除。
施肥措施在所有耕作处理中一致,播种时用播种机施入。玉米施底肥和追肥,氮肥、磷肥和钾肥的施入量分别为135 kg N·hm-2、51 kg P2O5·hm-2和51 kg K2O·hm-2。大豆只施底肥,氮肥、磷肥和钾肥的施用量分别为40 kg N·hm-2、49 kg P2O5·hm-2和53 kg K2O·hm-2。
1.3 土样采集
土样于2012年及2018年的4月下旬(播种前)进行采集。使用土钻(直径2.64 cm)在每个小区随机选取5点进行采样,采样深度为0—15 cm。5点采集的土壤汇合在一起作为一个重复,共24份(2种耕作方式×2种轮作方式×3次重复×2次采样时间)土壤样品。同时,用环刀法测定土壤容重(BD)。新鲜土样置于自封袋中,用采样箱运回实验室,保存在4 ℃下。1.4 基本理化性质的测定
风干土壤过2 mm筛后,采用水土比5﹕1测定土壤pH;过100目筛后,用元素分析仪测定土壤有机碳(SOC)和全氮(TN),测定的土壤理化性质见表1。Table 1
表1
表1土壤基本理化性质(均值±标准误)
Table 1
| 年份Year | 轮作方式Rotation | 耕作方式Tillage | 容重BD (g∙cm-3) | 土壤有机碳SOC (g∙kg-1) | 全氮TN (g∙kg-1) | pH |
|---|---|---|---|---|---|---|
| 2012 | CC | NT | 1.31±0.08a | 18.86±1.48a | 1.85±0.02a | 5.73±0.17a |
| CT | 1.03±0.07b | 18.00±0.56b | 1.65±0.01b | 5.95±0.31a | ||
| CS | NT | 1.27±0.04a | 17.27±0.53c | 1.70±0.01b | 5.98±0.22a | |
| CT | 1.10±0.07b | 16.90±0.33d | 1.65±0.01b | 6.11±0.10a | ||
| 2018 | CC | DNT | 1.13±0.03a | 17.84±0.26ab | 1.64±0.03a | 5.29±0.04ab |
| CT | 1.20±0.03a | 18.62±0.27a | 1.69±0.01a | 5.17±0.02b | ||
| CS | DNT | 1.12±0.01a | 18.11±0.57ab | 1.64±0.06a | 5.17±0.08ab | |
| CT | 1.11±0.03a | 17.30±0.09b | 1.60±0.03a | 5.42±0.01a |
新窗口打开|下载CSV
1.5 线虫提取与鉴定
在样品采集后的2周内完成线虫的提取,以确保线虫具有较强的生命活动。采用改良的浅盘法将50 g鲜土进行线虫的分离提取,60℃温热杀死后,用4%福尔马林固定,线虫总数通过解剖镜直接确定,然后按测得的土壤水分,折算成100 g干土中土壤线虫的数量。从每个样品中随机抽100条线虫(不足100条的全部鉴定),在光学显微镜下进行科属鉴定[16,25-26],依据取食习性和食道特征划分为4个营养类群:食细菌线虫(bacterivores,Ba)、食真菌线虫(fungivores,Fu)、植物寄生线虫(plant-parasites,PP)和杂食/捕食线虫(omnivores-predators,OP)。本试验鉴定出的线虫属已列入电子附表1中。1.6 线虫生态指数计算
根据土壤线虫的分类结果计算生态指数[25,26,27,28]:香农多样性指数(H'):H'=-∑pi(lnpi)
式中,pi是第i个分类单元中个体所占的比例。
均匀度指数(J):J=H'/lnS
式中,H’是香农多样性指数;S为鉴定分类单元的数目。
丰富度指数(SR):SR=(S-1/lnN)
式中,S为鉴定分类单元的数目,N为鉴定的线虫个体总数。
线虫通道指数(NCR):NCR=B/(B+F)
式中,B和F分别为食细菌线虫和食真菌线虫数量占线虫总数的相对丰度。
线虫成熟度指数:包括自由生活线虫成熟度指数(MI)和植物寄生线虫成熟度指数(PPI):
MI或PPI=-∑v(i)f(i)
式中,v(i)为第i种自由生活线虫或植物寄生线虫的c-p值,f(i)为第i种自由生活线虫或植物寄生线虫的个体数占总个体数的比例。
线虫代谢足迹(nematode metabolic footprint, F):
F=∑{Nt[0.1Wt/mt+0.273(W0.75 t)]}
式中,Wt代表第t个属线虫的个体鲜重;mt代表第t个属线虫的c-p值[29];个体线虫鲜重及c-p值可登陆
1.7 数据分析
使用效应值(lnR)来度量相对于常规耕作(即对照组),长期免耕及长期免耕后扰动对土壤线虫属数、多度和生态指数的影响效应大小[30,31]:
式中,Xn与Xc分别表示不同免耕条件下及其对应的采样当年常规耕作处理下线虫属数或多度、生态指数。
效应值(lnR)的变异系数,通过公式(2)计算:
式中,VlnR为效应值lnR的变异系数,SDc与SDn分别为常规耕作和不同免耕条件下的标准差,Nc和Nn分别为常规耕作和不同免耕条件处理的样本数。
该指标是Hedges在1999年提出的一个综合衡量指标,常用于评估生态因子的效应大小[32]。lnR数值越大,表明不同免耕条件对土壤线虫属数、多度或生态指数影响越大;lnR>0表示NT或DNT处理对土壤线虫属数、多度和生态指数有促进效应;lnR<0则表示NT或DNT处理对土壤线虫属数、多度和生态指数有抑制效应[33]。
采用Excel 2010进行数据的整理和计算;利用Origin 2021完成长期免耕或长期免耕扰动对土壤线虫属数、多度和生态指数影响的作图。对数据进行正态分布和方差齐性检验,对不满足正态分布和方差齐性的数据进行lg转换;运用T检验检验长期免耕或长期免耕后扰动对线虫群落结构的影响,以常规耕作作为对照组,P<0.05时免耕与常规耕作之间差异达到显著水平。选择至少在一个处理中相对多度大于1%且在所有处理中总多度大于5%的线虫属,利用Canoco 5.0的主成分分析(PCR)进行各处理中土壤线虫的群落组成分析,冗余分析(RDA)分析进行土壤线虫群落组成与土壤因子之间的相关性分析。
2 结果
2.1 线虫属
从鉴定出的土壤线虫属数来看(图1),NTCC(免耕玉米连作)促进了杂食/捕食线虫(lnR=0.1335)和植物寄生线虫(lnR=0.2877)属数量的增加,显著抑制了食真菌线虫(lnR=-0.0953)属数量的增加(P<0.05)。然而,DNTCC(免耕后扰动玉米连作)显著(P<0.05)抑制杂食/捕食线虫属数(lnR=-0.2877),对植物寄生线虫属数(lnR=0)既不促进也不抑制。图1
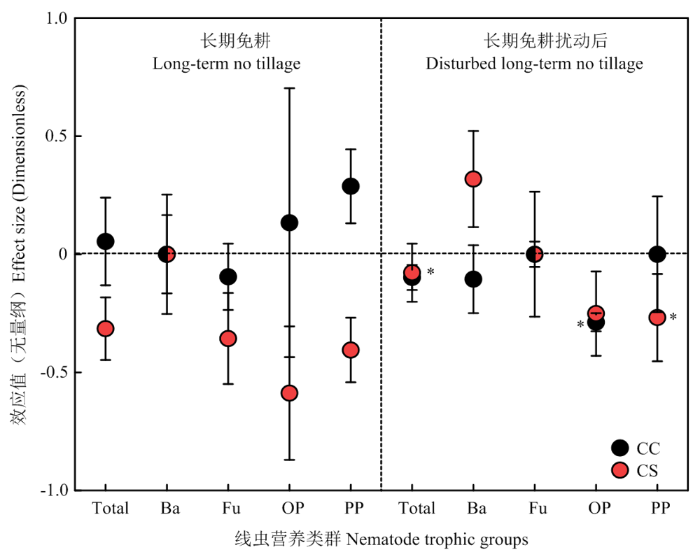 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1长期免耕或长期免耕后扰动对线虫属数的效应大小
Total:总线虫;Ba:食细菌线虫;Fu:食真菌线虫;OP:杂食/捕食线虫;PP:植物寄生线虫;CC:玉米连作;CS:玉米-大豆轮作。**和*分别表示显著性水平为P<0.01和P<0.05,CC显著性标在左侧,CS显著性标在右侧。
Fig. 1Effect sizes of long-term no-tillage or disturbed no-tillage on the number of nematode genera
Total: Total nematodes; Ba: Bacterivores; Fu: Fungivores; OP: Omnivores-predators; PP: Plant-parasites; CC: Corn continuous cropping; CS: Corn-soybean rotation. **,* Indicate significance levels at P<0.01 and P<0.05, respectively. CC significance is marked on the left, and CS significance is marked on the right. The same as
NTCS(免耕玉米-大豆轮作)对所有营养类群的属数均无促进效应;而DNTCS(免耕后扰动玉米-大豆轮作)促进了食细菌线虫(lnR=0.3185)属数量的增加,对线虫属总数(lnR=-0.0780)和植物寄生线虫(lnR=-0.2683)属数量抑制效应显著(P<0.05)(图1)。
2.2 线虫多度
从线虫多度来看(图2),NTCC除对杂食/捕食线虫的多度有促进效应之外,对其他营养类群的多度均无促进效应;DNTCC对所有营养类群多度都有较强的抑制效应,其中显著(P<0.05)抑制食真菌线虫多度(lnR=-1.0901)。图2
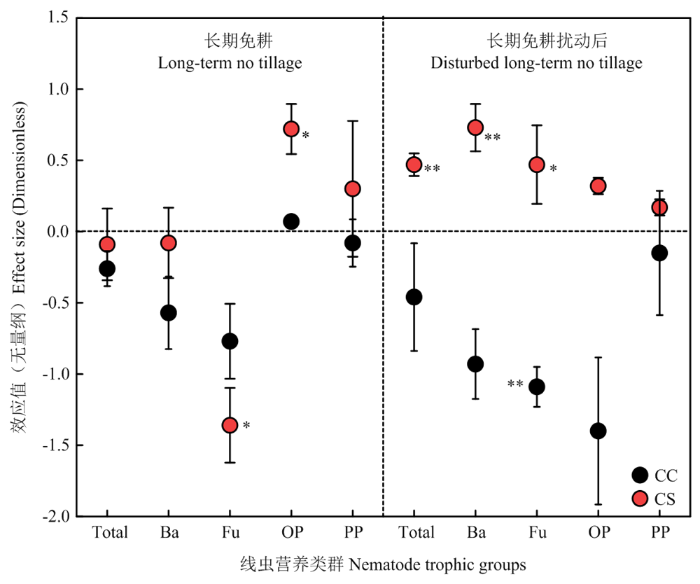 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2长期免耕或长期免耕后扰动对线虫营养类群多度的效应大小
Fig. 2Effect sizes of long-term no-tillage or disturbed no-tillage on nematode abundance
NTCS对杂食/捕食线虫多度(lnR=0.7237)促进效应显著(P<0.05),而对食真菌线虫多度(lnR=-1.3606)抑制效应显著(P<0.05)。DNTCS对所有营养类群的多度都有一定程度的促进效应,尤其对食细菌线虫(lnR=0.7276)、食真菌线虫(lnR=0.4651)和线虫总多度(lnR=0.4749)有显著(P<0.05)的促进效应(图2)。
2.3 线虫群落组成
主成分(PCA)分析发现,长期免耕阶段(图3-A),前两个轴共解释了线虫群落组成47.26%的变化,并且可显著区分不同耕作方式下土壤线虫群落组成,而无法区分不同轮作方式下线虫群落组成。NT优先促进了狭咽属(Discolaimium,OP5)、叉针属(Boleodorus,PP2)的分布。在长期免耕扰动后(图3-B),PCA分析发现前两个轴共解释了线虫群落组成58.99%的变化,并且可显著区分不同耕作方式下土壤线虫群落组成。DNT优先促进了威尔斯属(Wilsonema,Ba2)和新萨达属(Neothada,PP2)的分布,抑制了狭咽属(Discolaimium,OP5)、矮化属(Amplimerlinius,PP3)、丝尾垫刃属(Filenchus,Fu2)和表矛线属(Epidorylaimus,OP4)等属的分布。图3
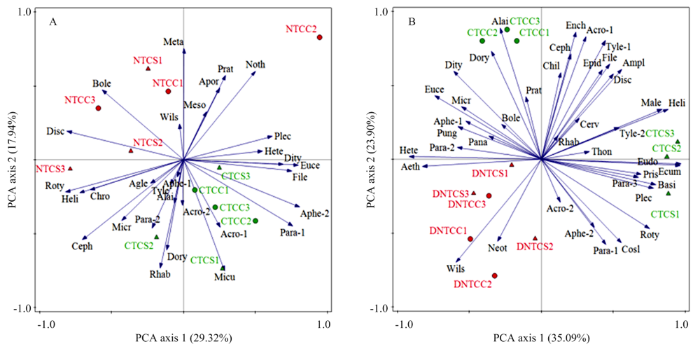 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3土壤线虫属与样方之间相关性的主成分分析(PCA)排序图
A:长期免耕阶段;B:长期免耕扰动后阶段;线虫属名用全名的前4个字母代替,若前4个字母相同,则按在电子附表1中顺序标上编号,全名见电子附表1。
CTCC:常规耕作玉米连作;CTCS:常规耕作玉米-大豆轮作;NTCC:免耕玉米连作;NTCS:免耕玉米-大豆轮作;DNTCC:免耕后扰动玉米连作;DNTCS:免耕后扰动玉米-大豆轮作
Fig. 3Principal component analysis (PCA) of the relationship between nematode genera and samples
A: Long-term no-tillage stage; B: Long-term no-tillage stage after disturbance. The name of the nematode genus is replaced by the first four letters of the full name. If the first four letters are the same, they are numbered in the order of Attached
CTCC: Conventional tillage with corn continuous cropping; CTCS: Conventional tillage with corn-soybean rotation; NTCC: No-till with corn continuous cropping; NTCS: No-till with corn-soybean rotation; DNTCC: Disturbed no-tillage with corn continuous cropping; DNTCS: Disturbed no-tillage with corn-soybean rotation
冗余分析(RDA)发现,长期免耕阶段(图4-A),第一个典型轴和所有典型轴的特征值分别为0.1266和0.3231,所有环境变量可以解释线虫群落组成32.3%的变异,其中10.5%的变异由全氮含量解释。全氮含量与真滑刃属(Aphelenchus,Fu2)、滑刃属(Aphelenchoides,Fu2)、伪垫刃属(Nothotylenchus,Fu2)、连胃属(Chronogaster,Ba2)、头叶属(Cephalobus,Ba2)具有正相关关系。
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4土壤线虫属与土壤因子之间相关性的冗余分析(RDA)排序图
BD:容重;SOC:土壤有机碳;TN:全氮
Fig. 4Redundancy analysis (RDA) of the relationship between nematode genera and soil physiochemical properties
BD: Bulk density; SOC: Soil organic carbon; TN: Total nitrogen
长期免耕后扰动阶段(图4-B),冗余分析(RDA)发现,第一个典型轴和所有典型轴的特征值分别为0.1456和0.3171,所有环境变量可以解释线虫群落组成31.7%的变异,其中11.4%的变异由容重解释。容重与大矛属(Enchodelus,OP4)、螯属(Pungentus,OP4)、狭咽属(Discolaimium,OP5)、无孔小咽属(Alaimus,Ba4)、真头叶属(Eucephalobus,Ba2)、滑刃属(Aphelenchoides,Fu2)等都具有正相关关系。
2.4 线虫生态指数
从线虫生态指数来看(图5),NTCC显著(P<0.05)促进Fs(lnR=0.5141),而显著(P<0.05)抑制Fe(lnR=-0.6637)。DNTCC对所有生态指数都具有抑制效应,且显著(P<0.05)抑制H'、J、SR、Fs和Ff。其中,对Fs和Ff抑制效应最大,效应值分别为lnR=-1.3089、lnR=-1.5628。图5
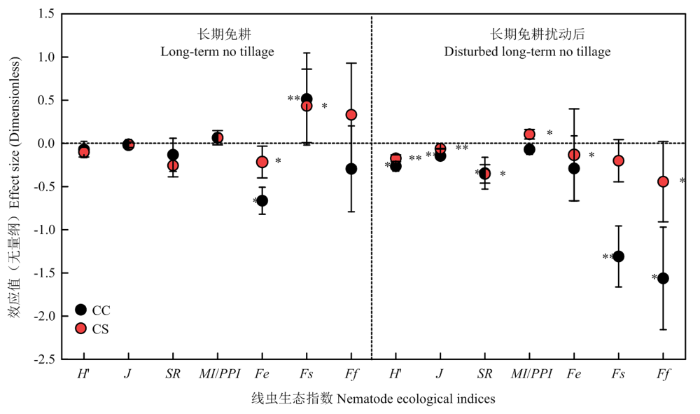 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5长期免耕或长期免耕后扰动对线虫生态指数的效应大小
CC:玉米连作;CS:玉米-大豆轮作。**和*分别表示显著性水平为P<0.01和P<0.05。CC显著性标在左侧,CS显著性标在右侧
Fig. 5Effect sizes of long-term no-tillage or disturbed no-tillage on nematode ecological indices
CC: corn continuous cropping; CS: corn-soybean rotation. **,* Indicate significance levels at P<0.01 and P<0.05, respectively. CC significance is marked on the left, and CS significance is marked on the right
NTCS显著(P<0.05)促进Fs(lnR=0.4346),而显著(P<0.05)抑制Fe(lnR=-0.2157)。DNTCS除对MI/PPI(lnR=0.1056)有显著(P<0.05)促进效应之外,对其他生态指数都有抑制效应,其中对H'、J、SR、Fe和Ff抑制效应显著(P<0.05)(图5)。
3 讨论
3.1 线虫群落结构对土壤扰动的响应
本研究发现同一轮作方式下,土壤线虫各营养类群对NT及DNT处理的响应程度并不一致。在CC下,杂食/捕食线虫比其他营养类群对长期免耕扰动前/后的响应更加明显。杂食/捕食线虫属的数量及其多度的变化趋势均由长期免耕扰动前的促进效应转变为扰动后的抑制效应(图1,图2)。这主要是因为杂食/捕食线虫对土壤扰动响应极为敏感,在遭到深翻整地的土壤扰动后其多度会迅速下降甚至一些属会消失[25],如PCA结果发现,c-p值为5的狭咽属(Discolaimium,OP),其多度与NT高度正相关(图3-A);但在遭到深翻和移除秸秆等土壤扰动后其多度迅速降低(图3-B)。杂食/捕食线虫是线虫微食物网的顶级捕食者,可以通过捕食作用“自上而下”的调控其他线虫营养类群(植物寄生线虫和食微线虫)。研究发现在CS下,NT处理对杂食/捕食线虫多度的促进效应由扰动前的lnR=0.7237下降到扰动后的lnR=0.3158(降幅为0.4079),食细菌线虫的属数及其多度、食真菌线虫的多度均由长期免耕扰动前的抑制效应转变为扰动后的促进效应。这些研究结果表明,在长期免耕土壤上实施土壤扰动后会通过削弱杂食/捕食线虫“自上而下”的级联效应[34],促进线虫微食物网中营养级较低的食微线虫多样性的增加。长期免耕扰动前或扰动后土壤线虫群落组成相对常规耕作的差异取决于其居住的土壤微环境变化。冗余分析发现,长期免耕阶段中土壤全氮含量对线虫群落组成影响最大,可解释其10.5%的变化;并且全氮含量越高越有利于c-p值为2的食细菌和食真菌线虫相对多度的增加(图4-A)。张爱林等[35]的研究结果也显示线虫类群中c-p值为2的机会主义者会随着施氮水平的增加而显著增加。在所有轮作方式下,NT土壤中全氮含量均高于CT(表1),表明NT有利于土壤养分富集,可以通过促进机会主义者“自下而上”地维持营养级较高的杂食/捕食线虫的活性和生存。在长期免耕后扰动阶段中,土壤容重对线虫群落种属组成影响最大,可解释其11.4%的变化。c-p值为4的食细菌线虫及c-p值为4、5的杂食/捕食线虫的相对多度与土壤容重具有正相关关系,表明土壤容重是限制较高c-p值线虫营养类群生存的重要因子(图4-B)。ZHANG等[15]也发现土壤容重与狭咽属(Discolaimium,OP)等c-p值较高的线虫属具有正相关关系。土壤容重是一个集成指标,可以有效反映土壤孔隙大小及土壤水分的状况[36,37]。线虫生活在土壤颗粒周围和颗粒之间的水膜中,充水孔隙的增加会促进线虫的运动和存活[38]。因此,土壤容重的适当增加有利于较高c-p值线虫营养类群的生存发展。然而,在对长期免耕土壤进行深翻和秸秆移除等土壤扰动后,其容重比扰动前呈明显减少趋势(表1),所以长期免耕土壤受到扰动后会显著降低c-p值>3线虫营养类群的多度。
3.2 线虫生态指数对土壤扰动的响应
线虫生态指数可以全面、深入地反映土壤线虫群落结构在不同田间管理下的差异[39]。研究发现,在两种轮作方式下,线虫生态指数的效应值在长期免耕阶段呈现正、负效应值交互模式;然而在长期免耕后扰动阶段,几乎所有生态指数(除MI/PPI之外)的效应值都呈负响应趋势(图5)。其中变化趋势最大的是Fs,其效应值由长期免耕扰动前的促进效应转变为扰动后的抑制效应,在CC和CS下分别降低到-1.3089和-0.2006。Fs是结构代谢足迹,反映了c-p值为3—5的营养级较高的土壤线虫对资源的代谢活性[20],对食物网的稳定性具有重要的调节功能[40]。NT对Fs的促进效应已在长期实施保护性耕作技术的欧洲地区所证实[13],并指出NT通过减少耕作及增加有机物料为土壤线虫提供了干扰较少且食物资源丰富的生存环境。我们的结果表明,在长期免耕土壤实施土壤扰动,如深翻和移除秸秆等措施,会破坏土壤线虫群落结构的稳定性。但值得注意的是,在长期免耕扰动阶段中,CS对MI/PPI具有显著的促进效应。MI/PPI是自由生活线虫成熟度与植物寄生线虫成熟度比值,常用于表征外界条件的改变对土壤环境稳定性的影响[41];其值越高,说明土壤环境越稳定。对长期免耕土壤实施土壤扰动后,CS仍能维持对MI/PPI的促进效应,表明CS能在一定程度上抵抗土壤扰动带来的对线虫群落结构稳定性的破坏。这可能是由于与玉米连作相比,玉米-大豆轮作会增加土壤氮素及其他高质量作物残体的输入量,可以通过维持微生物的活性从食物资源上影响线虫群落结构,从而使土壤线虫群落结构具有一定的抗干扰能力[42]。总之,我们的研究结果表明,长期免耕后实施土壤扰动,如深翻和移除秸秆等措施,会破坏土壤线虫群落结构的稳定性,但破坏程度取决于轮作方式。与CC相比,CS能够削弱深翻和移除秸秆对土壤线虫群落结构的破坏作用。4 结论
与常规耕作(CT)相比,免耕(NT)能够在各轮作方式下形成较为稳定的土壤线虫群落结构。然而,在长期免耕土壤实施深翻和移除秸秆等扰动措施会破坏土壤线虫群落结构的稳定性,但破坏程度因轮作方式而异。与玉米连作(CC)相比,玉米-大豆轮作(CS)仍能维持对MI/PPI的促进效应,表明CS能在一定程度上抵抗土壤扰动带来的对线虫群落结构稳定性的破坏。这可能是由于大豆的种植向土壤输入了高质量的植物残体,可以维持土壤生物对碳源的需求,进而增强了土壤线虫群落结构抵抗外界扰动的能力。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/gcb.2004.10.issue-2URL [本文引用: 2]
DOI:10.1016/j.apsoil.2019.03.006URL [本文引用: 2]
DOI:10.2134/agronj14.0443URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1038/s41586-019-1418-6URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.17520/biods.2018082URL [本文引用: 1]

土壤线虫生态学主要探讨土壤线虫群落和其周围环境(包括生物和非生物)的相互关系, 包括不同生态系统中土壤线虫群落的分布和结构组成、线虫群落与土壤环境及其他土壤生物之间的相互作用等。本文回顾了我国研究者近年来在土壤线虫生态学研究领域的研究现状, 包括不同生态系统土壤线虫群落的分布、组成和多样性及其影响因素, 土壤线虫群落与全球气候环境变化的关系, 土壤线虫群落的生态功能以及土壤线虫群落生态学分析方法的发展及应用。重点评述近年来我国土壤线虫生态学的发展现状, 同时分析和比较了国内外土壤线虫生态学的发展态势, 提出建设全国范围的监测网络的重要性。未来我国土壤线虫生态学的发展方向应继续加强小尺度下土壤微食物网联通性和大尺度下全球气候变化对土壤线虫群落影响的研究以及加强相关新的研究技术方法的应用。
DOI:10.17520/biods.2018082URL [本文引用: 1]
土壤线虫生态学主要探讨土壤线虫群落和其周围环境(包括生物和非生物)的相互关系, 包括不同生态系统中土壤线虫群落的分布和结构组成、线虫群落与土壤环境及其他土壤生物之间的相互作用等。本文回顾了我国研究者近年来在土壤线虫生态学研究领域的研究现状, 包括不同生态系统土壤线虫群落的分布、组成和多样性及其影响因素, 土壤线虫群落与全球气候环境变化的关系, 土壤线虫群落的生态功能以及土壤线虫群落生态学分析方法的发展及应用。重点评述近年来我国土壤线虫生态学的发展现状, 同时分析和比较了国内外土壤线虫生态学的发展态势, 提出建设全国范围的监测网络的重要性。未来我国土壤线虫生态学的发展方向应继续加强小尺度下土壤微食物网联通性和大尺度下全球气候变化对土壤线虫群落影响的研究以及加强相关新的研究技术方法的应用。
DOI:10.1016/0167-8809(93)90074-YURL [本文引用: 1]
DOI:10.1016/j.still.2006.11.008URL [本文引用: 1]
DOI:10.1016/j.soilbio.2013.03.023URL [本文引用: 1]
DOI:10.1111/mec.v28.22URL [本文引用: 2]
DOI:10.1016/j.apsoil.2006.09.008URL [本文引用: 1]
DOI:10.1016/j.ejsobi.2014.11.006URL [本文引用: 2]
DOI:10.1007/BF00324627URL [本文引用: 2]
[本文引用: 1]
DOI:10.1126/science.269.5228.1257URL [本文引用: 1]
DOI:10.2136/sssaj2006.0174URL [本文引用: 1]
DOI:10.1038/35012234URL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/S1002-0160(07)60073-3URL [本文引用: 1]
DOI:10.1016/j.tree.2016.02.016URL [本文引用: 1]
[本文引用: 3]
DOI:10.1016/S0929-1393(98)00123-1URL [本文引用: 2]
DOI:10.1007/s003740050320URL [本文引用: 1]
DOI:10.1007/s00374-003-0659-5URL [本文引用: 1]
DOI:10.1016/j.ejsobi.2010.01.003URL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s00442-011-1909-0URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1890/0012-9658(1999)080[1150:TMAORR]2.0.CO;2URL [本文引用: 1]
DOI:10.1111/oik.2004.107.issue-2URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.ejsobi.2012.02.005URL [本文引用: 1]
DOI:10.1111/geb.2014.23.issue-9URL [本文引用: 1]
DOI:10.1016/j.still.2019.05.017URL [本文引用: 1]
DOI:10.1016/S0929-1393(01)00152-4URL [本文引用: 1]
DOI:10.1016/j.apsoil.2012.04.006URL [本文引用: 1]
DOI:10.1016/S0929-1393(02)00089-6URL [本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}