 ,1, 王壮壮,1, 谷丰序1, 牟海萌1, 王宇1, 段剑钊1, 冯伟1,2, 王永华,1,2, 郭天财,1,2
,1, 王壮壮,1, 谷丰序1, 牟海萌1, 王宇1, 段剑钊1, 冯伟1,2, 王永华,1,2, 郭天财,1,2Effects of Nitrogen Fertilizer and Plant Density on Carbon Metabolism, Nitrogen Metabolism and Grain Yield of Two Winter Wheat Varieties
WANG JinFeng,1, WANG ZhuangZhuang,1, GU FengXu1, MOU HaiMeng1, WANG Yu1, DUAN JianZhao1, FENG Wei1,2, WANG YongHua,1,2, GUO TianCai,1,2通讯作者:
责任编辑: 杨鑫浩
收稿日期:2020-11-25接受日期:2021-02-7
| 基金资助: |
Received:2020-11-25Accepted:2021-02-7
作者简介 About authors
王金凤,E-mail:
王壮壮,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (612KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
王金凤, 王壮壮, 谷丰序, 牟海萌, 王宇, 段剑钊, 冯伟, 王永华, 郭天财. 氮密调控对两个冬小麦品种碳氮代谢及产量的影响. 中国农业科学, 2021, 54(19): 4070-4083 doi:10.3864/j.issn.0578-1752.2021.19.004
WANG JinFeng, WANG ZhuangZhuang, GU FengXu, MOU HaiMeng, WANG Yu, DUAN JianZhao, FENG Wei, WANG YongHua, GUO TianCai.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】碳代谢和氮代谢是小麦生长发育的两大基础代谢,二者既相互促进又激烈竞争,协同调控小麦植株的生长发育,同时二者又受基因型、栽培技术和环境的影响[1,2]。目前生产实践中,大穗型、多穗型和中间型小麦依靠不同的栽培措施均可实现超高产[3,4]。氮素是构成核酸、叶绿体、蛋白质等重要结构的基础元素,也是影响作物生长的最主要限制因子之一。合理施氮可调控土壤溶解性有机碳和有机氮,也可调节植株的碳氮平衡[2, 5-6]。另有研究表明,栽培措施亦能调控植株的碳氮营养,例如适宜种植密度可通过改善作物对水、肥的利用状况而影响冬小麦碳、氮代谢进程 [7,8]。然而在小麦实际生产中,多数农户存在盲目选种、过量施肥和大播量种植等问题,直接影响植株碳氮代谢平衡,造成植株旺长、群体郁闭,不利于产量形成和提高[9]。因此,合理调整施氮量和种植密度可以调控冬小麦群体生长结构,改善植株碳、氮代谢平衡,对提高冬小麦综合生产能力和实现可持续生产具有重要意义。【前人研究进展】已有研究表明,合理施氮有利于促进作物植株地上部器官和地下部根系的生长发育,有利于提高氮源对籽粒灌浆的供氮能力,增加灌浆期籽粒可溶性糖和蛋白质的含量,降低膜脂过氧化程度,增强氮素和光合产物向籽粒中的转移,进而提高氮肥利用率[10,11,12],籽粒产量与氮素吸收和氮素收获指数呈显著正相关[13]。对不同小麦生态区适宜施氮量的研究发现,245、300、227和212 kg·hm-2施氮量可分别作为豫东地区、新疆干旱地区、黄土高原东南部和西北部的最佳施氮量[12, 14-15]。研究表明,增加种植密度可通过增加优势茎蘖数来提高产量[16],高密度种植对作物生长发育的影响主要与植株间相互遮荫及养分竞争有关,前者的影响效应大于后者[17],且高密度种植有利于促进小麦籽粒中氮素的积累、增强氮同化酶活性,进而提高籽粒蛋白质含量[18,19,20];另有研究表明,低密度种植更有利于冬小麦光合作用[21,22],尤其在播期、行距、光能、分蘖成穗等生长条件均适宜时,极低种植密度亦能获得高产[23]。【本研究切入点】品种、施氮量和种植密度均会影响小麦植株的生长发育,进而影响植株的碳氮代谢,而碳氮代谢在生殖生长期又影响籽粒灌浆结实,最终决定产量和品质。鉴于碳、氮代谢对籽粒灌浆的重要性,深入研究氮密调控在小麦生殖生长期对植株碳、氮代谢的影响,有利于促进小麦高产高效生产。【拟解决的关键问题】本研究以分蘖力中等、成穗率较高的冬小麦鑫华麦818和分蘖力强、成穗率中等的冬小麦百农207为供试材料,探讨施氮量和种植密度对2个冬小麦品种碳、氮代谢及籽粒产量的影响,旨在明确豫东南黏壤潮土区冬小麦适宜的氮密调控栽培管理措施,以期为冬小麦高产高效栽培提供技术支撑。1 材料与方法
1.1 试验地概况
本试验于2018—2020年在河南省周口市商水县亿港农场(33°31′N,114°42′E)进行。该区属暖温带大陆性季风气候,年平均气温15.0℃,常年平均降雨量751.7 mm(数据由1990—2020年连续30年的商水县气象资料统计所得)。2018—2020 两年度冬小麦生长季的月平均降水量及月平均温度变化如图1所示,2018、2019年冬小麦冬前积温分别为641.2℃和562.2℃。该区域土壤属于黏壤潮土,2018、2019年冬小麦播种前0—20 cm和20—40 cm土层土壤基础养分结果如表1所示。图1
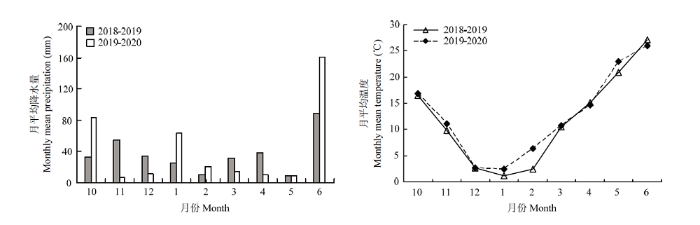 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图12018—2020年冬小麦生长季降水量及温度变化
Fig. 1Dynamics of precipitation and temperature during winter wheat growing season in 2018-2020
Table 1
表1
表1播前土壤基础养分
Table 1
| 年份 Year | 品种 Variety | 土层 Soil layers (cm) | 有机质 Organic matter (g·kg-1) | 全氮 Total N (g·kg-1) | 碱解氮 Alkaline N (mg·kg-1) | 速效钾 Available K (mg·kg-1) | 有效磷 Available P (mg·kg-1) | pH |
|---|---|---|---|---|---|---|---|---|
| 2018-2019 | 鑫华麦818 Xinhua 818 | 0-20 | 17.82 | 1.27 | 102.97 | 263.67 | 4.17 | 8.19 |
| 20-40 | 9.44 | 0.86 | 78.64 | 192.44 | 2.99 | 8.31 | ||
| 百农207 Bainong 207 | 0-20 | 16.54 | 1.21 | 102.47 | 299.47 | 3.90 | 8.12 | |
| 20-40 | 13.98 | 1.13 | 88.04 | 246.16 | 3.34 | 8.21 | ||
| 2019-2020 | 鑫华麦818 Xinhua 818 | 0-20 | 17.60 | 1.17 | 94.38 | 157.92 | 6.49 | 8.16 |
| 20-40 | 7.97 | 0.75 | 74.39 | 112.95 | 3.50 | 8.24 | ||
| 百农207 Bainong 207 | 0-20 | 17.78 | 1.16 | 96.52 | 211.90 | 6.13 | 8.07 | |
| 20-40 | 8.54 | 0.89 | 76.96 | 156.68 | 3.77 | 8.16 |
新窗口打开|下载CSV
1.2 试验设计
试验地前茬为玉米,秸秆全部粉碎还田。试验采用裂裂区设计,以品种为主区,施氮量为副区,种植密度为副副区,各处理重复3次,每小区面积为40 m2(4 m×10 m)。设置品种、种植密度和施氮量3个因子,供试品种为分蘖力中等、成穗率较高的冬小麦鑫华麦818(A)和分蘖力强、成穗率中等的冬小麦百农207(B);施氮量设3个水平,分别为0、240、360 kg·hm-2,依次标记为N0、N240、N360;种植密度设3个水平,分别为225、375和525 万株/hm2,依次标记为M1、M2和M3。播前50%的氮肥底施、120 kg·hm-2的P2O5、90 kg·hm-2的K2O一次性全部底施,剩余50%氮肥于拔节期追施。2018年和2019年的播种日期分别为10月15日和10月24日,收获日期分别为6月10日和5月29日。小麦生长季灌水和病虫草害防治按照一般大田进行。1.3 测定项目与方法
1.3.1 取样方法 于孕穗期每处理小区随机选取60个叶片大小、着生态位、长势基本一致的代表性单茎,其中50片小麦旗叶用锡纸包裹之后,用液氮浸泡运回实验室置于-80℃冰箱待测,剩余10片旗叶取样后在烘箱内105℃杀青30 min,80℃烘干至恒重待测。开花期选取代表性单茎挂牌标记,分别于开花期、花后10 d、花后20 d和花后30 d取挂牌标记的小麦旗叶,取样数量和处理方式与孕穗期一致。测定碳代谢、氮代谢指标时均使用混合样。1.3.2 碳代谢指标 可溶性糖含量的测定使用旗叶干样,采用蒽酮比色法测定。磷酸蔗糖合成酶(SPS)活性和蔗糖合成酶(SS)活性的测定使用旗叶鲜样,测定方法参考DOEHLERT等[24]方法。
1.3.3 氮代谢指标 氮代谢指标的测定均使用旗叶鲜样。可溶性蛋白质含量的测定采用考马斯亮蓝法。谷氨酰胺合成酶(GS)活性的测定参考王小纯等[25]的测定方法,将1 g鲜样组织在1 mL反应体系中1 min使540 nm下的吸光值变化0.01定义为一个酶活力单位。硝酸还原酶(NR)活性的测定参照武丽等[26]测定方法。
1.3.4 产量 各处理小区计产面积为6 m2,每小区重复3次。小麦成熟期各处理选取有代表性的1 m双行固定样点并调查其穗数。收获期各种植小区均选取代表性植株单茎50个,装入纱网袋,做好标记,带回室内测定各处理穗粒数,并脱粒风干后测定其千粒重,按含水量13%折算成实际产量。
1.4 数据计算与分析
采用Excel 2013进行数据处理和图表绘制,通过SPSS 23.0统计分析软件进行相关分析(Pearson相关)和差异显著性检验(Duncan法)。2 结果
2.1 品种、密度和氮肥及其互作对冬小麦碳氮代谢与产量构成因素的方差分析
方差分析结果表明,两年度品种、密度和氮肥对不同时期冬小麦可溶性糖含量、SPS酶活性和SS酶活性调控的主效应及其互作效应均达显著水平(表2)。从单因素来看,品种、密度和氮肥均显著影响冬小麦灌浆中后期可溶性蛋白质含量、NR酶活性和GS酶活性。从双因素来看,二因素交互作用对灌浆中后期可溶性蛋白质含量有显著影响,而两年度氮代谢酶活性方差分析结果存在差异,2018—2019年灌浆中期GS酶活性和2019—2020年灌浆中期NR酶活性受品种×氮肥的影响不显著,品种×密度和氮肥×密度均显著影响灌浆中后期氮代谢酶活性。品种×密度×氮肥三因素交互作用显著影响灌浆中后期氮代谢。2个品种不同施氮量的产量及其构成因素存在显著差异,种植密度对穗数和穗粒数的影响亦达显著水平,但对千粒重和产量的影响2年结果不一致。由此可见,氮肥是2个栽培措施中的主效应,种植密度主要通过调控穗数和穗粒数而影响产量。品种×密度显著影响穗粒数和千粒重,但对产量的影响不显著;品种×氮肥和氮肥×密度显著影响穗粒数,进而显著调控产量。品种×密度×氮肥三因素交互作用仅显著影响穗粒数,而对穗数、千粒重和产量的调控效果不显著。碳代谢生理参数年际间差异较为显著,氮代谢生理参数年际间差异在不同生育时期无明显规律,两年度穗数、穗粒数和产量间差异均达极显著水平,但千粒重间差异不显著。Table 2
表2
表2品种、密度和氮肥对冬小麦碳代谢、氮代谢及产量构成因素的方差分析
Table 2
| 指标 Item | 时期 Time | 2018―2019 | 2019―2020 | 年份 Year | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 品种 V | 密度 D | 氮肥 N | 品种× 密度 V×D | 品种× 氮肥 V×N | 密度× 氮肥 D× N | 品种×密 度×氮肥 V×D× N | 品种 V | 密度 D | 氮肥 N | 品种× 密度 V×D | 品种× 氮肥 V×N | 密度× 氮肥 D× N | 品种×密 度×氮肥 V×D× N | |||
| 可溶性糖含量 Soluble sugar content | -X | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | NS |
| 0 | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | NS | |
| 10 | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | NS | |
| 20 | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| 30 | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| SPS 酶活性 SPS activity | -X | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| 0 | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| 10 | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| 20 | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| 30 | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| SS 酶活性 SS activity | -X | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| 0 | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| 10 | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| 20 | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| 30 | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| 可溶性蛋白质含量 Soluble protein content | -X | *** | NS | *** | *** | *** | *** | *** | *** | ** | *** | *** | *** | ** | *** | ** |
| 0 | *** | *** | *** | *** | *** | NS | NS | *** | *** | *** | *** | *** | *** | *** | *** | |
| 10 | *** | *** | *** | ** | *** | NS | NS | *** | *** | *** | *** | *** | NS | *** | *** | |
| 20 | *** | *** | *** | * | *** | ** | *** | *** | *** | *** | *** | *** | *** | *** | NS | |
| 30 | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| NR 酶活性 NR activity | -X | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | NS |
| 0 | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | NS | |
| 10 | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| 20 | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | NS | *** | *** | ** | |
| 30 | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| GS 酶活性 GS activity | -X | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| 0 | ** | *** | *** | *** | *** | ** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| 10 | NS | *** | *** | *** | *** | *** | * | *** | *** | *** | *** | NS | *** | *** | *** | |
| 20 | *** | *** | *** | *** | NS | *** | *** | *** | *** | *** | *** | *** | ** | *** | *** | |
| 30 | *** | *** | *** | *** | *** | *** | *** | * | *** | *** | *** | *** | *** | *** | NS | |
| 穗数 Spike number | *** | *** | *** | NS | NS | NS | NS | *** | *** | *** | NS | ** | *** | NS | *** | |
| 穗粒数 Grain per spike | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | * | *** | * | *** | |
| 千粒重 1000-grain weight | *** | NS | *** | * | *** | * | NS | *** | *** | *** | *** | NS | NS | NS | NS | |
| 产量Yield | *** | * | *** | NS | *** | ** | NS | *** | NS | *** | NS | *** | *** | NS | *** | |
新窗口打开|下载CSV
2.2 氮密调控对2个冬小麦品种碳代谢的影响
碳代谢对氮肥和密度的响应随品种和生育时期的变化有所不同,两年度趋势基本一致(图2)。相同种植密度下,不同施氮处理的鑫华麦818孕穗期、开花期均以不施氮的可溶性糖含量和碳代谢酶活性最高,而在开花期之后施氮显著提高了各碳代谢指标,花后10 d和花后30 d的可溶性糖含量、SPS、SS活性大小均表现为N240>N360>N0,花后20 d却表现为N360>N240>N0。与N0相比,灌浆期N240、N360的碳代谢生理参数分别增加48.58%、28.59%。百农207孕穗至花后20 d各碳代谢指标均随施氮量的增加而降低,其可溶性糖含量、SPS、SS活性大小均表现为N0>N240>N360,但花后30 d却表现为N240>N360>N0,与N0相比,N240、N360的碳代谢生理参数分别增加63.48%、31.46%。相同施氮下,不同种植密度的鑫华麦818孕穗期和花后30 d的可溶性糖含量、SPS、SS活性分别表现为M2>M3>M1和M2>M1>M3,其开花期和花后10 d、20 d的各碳代谢指标以M3最高。而百农207孕穗期至花后10 d的各碳代谢指标以M1或M3的均值最大,但其花后20 d、30 d则表现为M2>M1>M3。综合考虑品种、氮肥和密度三因素试验结果,两年度2个品种花后30 d各碳代谢生理参数均以M2N240处理最大,由此表明,适宜的氮密组合有利于增强灌浆后期的碳代谢生理功能,促进光合物质生产。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2氮肥和密度对2个冬小麦品种碳代谢的影响
不同小写字母表示同一时期下不同处理间差异显著(P<0.05)。下同
Fig. 2Effects of nitrogen fertilizer and density on carbon metabolism in two winter wheat varieties
Different lowercase letters indicate significant differences between different treatments under the same stage (P<0.05). The same as below
2.3 氮密调控对2个冬小麦品种氮代谢的影响
3种种植密度条件下,施氮均可显著提高可溶性蛋白质含量、NR酶活性和GS酶活性,且其不同氮代谢生理指标的增加比例在2个品种间存在差异(图3)。鑫华麦818整个灌浆期的可溶性蛋白质含量均以N240最高,NR、GS氮代谢酶活性仅在灌浆中期表现为N240>N360>N0,而在孕穗、开花、花后10 d和30 d则以N360最高。从灌浆中期来看,与N0相比,N240、N360的氮代谢生理参数分别增加104.12%、66.59%。两年度百农207各氮代谢生理指标参数除花后10 d外,其余测定时期均以N240最高。与N0相比,灌浆中期N240、N360的氮代谢生理参数分别增加97.61%、47.83%。各施氮处理下种植密度对2个品种冬小麦氮代谢的调控亦存在显著差异。不同种植密度间相比,鑫华麦818除孕穗期的可溶性蛋白质含量表现为M1>M2>M3外,其余各测定时期的氮代谢生理指标参数均以M2的数值最大,而百农207的可溶性蛋白质含量、NR和GS氮代谢酶活性仅在灌浆中期表现为M2>M3>M1或M2>M1>M3。综合考虑品种、氮肥和密度三因素试验结果,两年度2个品种花后20 d各氮代谢生理参数均以M2N240处理最大,由此表明,适宜的氮密组合有利于增强灌浆中期的氮代谢生理功能,促进氮素高效吸收利用。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3氮肥和密度对2个冬小麦品种氮代谢的影响
Fig. 3Effects of nitrogen fertilizer and density on nitrogen metabolism in two winter wheat varieties
2.4 氮密调控对2个冬小麦品种籽粒产量及其构成因素的影响
两年度氮密处理组合对2个品种籽粒产量及其构成因素均具有明显的调控效应,且趋势基本一致(表3)。相同种植密度下,施氮有利于提高2个品种穗数和穗粒数,且均以适量施氮(N240)的穗数、穗粒数最大,但千粒重随施氮量的增加而降低。在低密度M1种植条件下,2个品种籽粒产量均随施氮量的增加而增加,但中高密度M2、M3种植条件下,其籽粒产量则以N240最高。同一施氮处理下,随着种植密度的增加,穗数呈增加的趋势,但穗粒数、千粒重和产量的变化规律不一致。与百农207相比,鑫华麦818的穗数、千粒重、产量均有所增加,其平均增幅分别为10.17%、10.59%、10.98%,而其穗粒数较百农207有所减少,平均减幅为15.64%。2018—2019年试验生长季,鑫华麦818和百农207均以M2N240处理组合的产量最高,而2019—2020年试验生长季2个品种均以M1N360处理的产量最高,M2N240处理的产量次之,但二者差异不显著。与产量最低的M1N0处理相比,鑫华麦818在M1N360和M2N240处理下的产量增幅分别为56.48%、55.81%,百农207的平均增幅为136.24%、137.17%。Table 3
表3
表3氮肥与密度对两冬小麦品种产量及其构成因素的调控效应
Table 3
| 品种 Variety | 处理 Treatment | 2018―2019 | 2019―2020 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 穗数 Spike number (104·hm-2) | 穗粒数 Grain per spike | 千粒重 1000 grain weight (g) | 产量 Yield (kg·hm-2) | 穗数 Spike number (104·hm-2) | 穗粒数 Grain per spike | 千粒重 1000 grain weight (g) | 产量 Yield (kg·hm-2) | ||
| A | M1N0 | 555.84c | 36.44bc | 51.37ab | 8293.81c | 370.29e | 32.01d | 51.70a | 4504.86d |
| M1N240 | 632.50ab | 41.22a | 48.76de | 8726.52abc | 548.36abc | 36.74b | 51.06ab | 8439.39bc | |
| M1N360 | 596.67bc | 39.71a | 47.99e | 8864.79ab | 515.69d | 35.42c | 50.21ab | 9283.47a | |
| M2N0 | 613.06ab | 29.19e | 51.23ab | 8457.85bc | 514.19d | 14.52h | 49.89ab | 4581.60d | |
| M2N240 | 633.61ab | 39.55a | 49.11cde | 9074.18a | 561.19ab | 42.90a | 48.97abc | 9109.38ab | |
| M2N360 | 620.84ab | 34.24cd | 49.90bcd | 8725.23abc | 540.19bcd | 29.04e | 48.46bc | 7986.07c | |
| M3N0 | 626.67ab | 32.58d | 51.54a | 8482.79bc | 532.03cd | 19.03g | 48.49bc | 5077.76d | |
| M3N240 | 657.50a | 36.54b | 50.65abc | 8859.89ab | 573.25a | 29.26e | 46.34cd | 8666.20abc | |
| M3N360 | 657.50a | 34.89bc | 49.39cde | 8463.66bc | 552.36abc | 27.17f | 44.90d | 7944.19c | |
| B | M1N0 | 502.22c | 40.41c | 47.60a | 6717.02e | 306.27d | 41.90ab | 49.11a | 2626.90d |
| M1N240 | 541.95abc | 43.80b | 44.22b | 8109.46c | 526.28ab | 42.70ab | 46.02bc | 7934.59b | |
| M1N360 | 517.50bc | 40.14c | 41.51c | 8226.00abc | 513.78b | 42.70ab | 41.94e | 9195.11a | |
| M2N0 | 510.00bc | 34.36d | 48.35a | 7177.87d | 440.02c | 18.70c | 46.96ab | 2826.41d | |
| M2N240 | 585.28a | 46.57a | 41.33c | 8603.76a | 538.78ab | 45.80a | 44.89bcd | 9095.41a | |
| M2N360 | 539.17abc | 46.41a | 41.94c | 8322.05abc | 521.28ab | 42.50ab | 43.06de | 7661.80bc | |
| M3N0 | 526.67bc | 30.48e | 47.72a | 7410.25d | 517.53ab | 22.40c | 46.85ab | 3412.01d | |
| M3N240 | 588.61a | 42.06bc | 42.61c | 8513.11ab | 550.03a | 39.30ab | 45.95bc | 8632.98ab | |
| M3N360 | 561.67ab | 39.72c | 41.13c | 8147.44bc | 546.28ab | 38.30b | 43.86cde | 6904.84c | |
新窗口打开|下载CSV
此外,由图1和表3还可以看出,2020年3—4月降雨量较少,小麦春季生长发育的关键阶段出现较重的旱情,导致2个品种各氮密处理组合的穗数、穗粒数明显降低,其平均值较2019年分别降低15.85%、17.96%(鑫华麦818)和8.47%、8.15%(百农207)。
2.5 氮密调控下2个冬小麦品种碳氮代谢与产量构成因素的关系
碳代谢生理参数与穗数、穗粒数、千粒重之间总体呈正相关,氮代谢生理参数与穗数呈极显著正相关,与穗粒数相关性较弱,但与千粒重呈负相关或极显著负相关(表4)。相关分析结果进一步表明,鑫华麦818的穗数、穗粒数与碳代谢生理参数呈极显著正相关。由此表明,鑫华麦818对产量三要素的协调优于百农207,增强碳代谢有利于穗数、穗粒数和千粒重的增加,且分蘖成穗特性存在差异的2个品种碳代谢与产量的相关性表现不同。增强氮代谢不利于粒重的增加,其主要通过增加穗数来提高产量。Table 4
表4
表4碳代谢、氮代谢与产量构成因素的相关性分析
Table 4
| 指标 Item | 品种Variety | 时期Time | 可溶性糖含量 Soluble sugar content | SPS酶活性 SPS activity | SS酶活性 SS activity | 可溶性蛋白质含量 Soluble protein content | NR酶活性 NR activity | GS酶活性 GS activity |
|---|---|---|---|---|---|---|---|---|
| 穗数 Spike number | A | -X | -0.34* | 0.31* | 0.45** | 0.46** | 0.26 | 0.48** |
| 0 | 0.57** | 0.51** | 0.48** | 0.59** | 0.40** | 0.67** | ||
| 10 | 0.76** | 0.75** | 0.77** | 0.70** | -0.15 | 0.60** | ||
| 20 | -0.01 | 0.58** | 0.56** | 0.20 | 0.15 | 0.60** | ||
| 30 | 0.73** | 0.74** | 0.75** | 0.79** | -0.03 | 0.40** | ||
| B | -X | -0.24 | 0.11 | 0.20 | 0.83** | 0.40** | 0.73** | |
| 0 | -0.60** | -0.21 | 0.01 | 0.68** | 0.45** | 0.69** | ||
| 10 | -0.47** | 0.02 | 0.14 | 0.73** | 0.10 | 0.67** | ||
| 20 | -0.58** | 0.08 | -0.02 | 0.76** | 0.21 | 0.47** | ||
| 30 | 0.31* | 0.43** | 0.45** | 0.59** | 0.11 | 0.30* | ||
| 穗粒数 Grain per spike | A | -X | -0.09 | 0.36** | 0.36** | 0.19 | 0.04 | 0.22 |
| 0 | 0.07 | 0.27* | 0.35* | 0.04 | 0.10 | 0.42** | ||
| 10 | 0.15 | 0.39** | 0.40** | 0.33* | -0.08 | 0.28* | ||
| 20 | 0.04 | 0.07 | 0.00 | 0.01 | -0.05 | 0.36** | ||
| 30 | 0.50** | 0.41** | 0.42** | 0.45** | -0.24 | -0.01 | ||
| B | -X | 0.16 | 0.14 | 0.14 | 0.05 | -0.03 | 0.05 | |
| 0 | 0.03 | 0.08 | 0.09 | 0.14 | -0.04 | 0.02 | ||
| 10 | -0.08 | 0.05 | 0.06 | 0.21 | -0.09 | 0.04 | ||
| 20 | -0.14 | 0.00 | -0.02 | -0.03 | -0.24 | -0.01 | ||
| 30 | -0.03 | 0.06 | 0.06 | 0.22 | -0.26 | 0.10 | ||
| 千粒重 1000 grain weight | A | -X | 0.04 | 0.39** | 0.40** | -0.19 | -0.22 | -0.14 |
| 0 | -0.10 | 0.30* | 0.35** | -0.31* | -0.25 | 0.05 | ||
| 10 | -0.36** | 0.04 | 0.10 | -0.16 | -0.30* | 0.09 | ||
| 20 | -0.27* | -0.26 | -0.25 | -0.36** | -0.21 | 0.00 | ||
| 30 | 0.18 | 0.20 | 0.18 | 0.07 | -0.48** | -0.12 | ||
| B | -X | 0.62** | 0.30* | 0.18 | -0.56** | -0.32* | -0.46** | |
| 0 | 0.70** | 0.54** | 0.30* | -0.59** | -0.28* | -0.44** | ||
| 10 | 0.85** | 0.34* | 0.16 | -0.63** | -0.33* | -0.47** | ||
| 20 | 0.68** | 0.31* | 0.39** | -0.60** | -0.19 | -0.48** | ||
| 30 | -0.40** | -0.44** | -0.44** | -0.56** | -0.14 | -0.22 |
新窗口打开|下载CSV
2.6 氮密调控下2个冬小麦品种碳氮代谢与籽粒产量的关系
2个冬小麦品种碳代谢生理参数与籽粒产量的相关性均出现了负相关到正相关的转变,且品种间相关性“负-正”的转变时间早晚存在差异(表5)。鑫华麦818在孕穗、开花2个时期的旗叶碳代谢与籽粒产量呈负相关或不显著正相关,但花后10 d至30 d二者间的相关性一直呈极显著正相关;而百农207孕穗至花后20 d旗叶可溶性糖含量与籽粒产量的相关性均呈极显著负相关,SPS、SS酶活性与籽粒产量呈负相关或不显著正相关,但其花后30 d各碳代谢指标均与产量呈极显著正相关。另外,2个品种旗叶氮代谢在各测定时期均与产量呈显著或极显著正相关。鑫华麦818整个灌浆期的旗叶碳氮代谢生理参数与籽粒产量的相关性均呈显著或极显著正相关,而百农207只在灌浆后期(花后30 d)实现碳氮代谢与产量同步正相关。由此表明,碳氮代谢平衡有利于冬小麦产量构成因素的自我调节,最终获得较高的产量,这可能是鑫华麦818最终产量高于百农207的主要生理原因。Table 5
表5
表5碳代谢、氮代谢与产量的相关性分析
Table 5
| 品种 Variety | 时期 Time | 可溶性糖含量 Soluble sugar content | SPS酶活性 SPS activity | SS酶活性 SS activity | 可溶性蛋白质含量 Soluble protein content | NR酶活性 NR activity | GS酶活性 GS activity |
|---|---|---|---|---|---|---|---|
| A | -X | -0.68** | -0.04 | 0.14 | 0.78** | 0.60** | 0.46** |
| 0 | 0.02 | 0.21 | 0.22 | 0.63** | 0.72** | 0.71** | |
| 10 | 0.50** | 0.49** | 0.50** | 0.69** | 0.28* | 0.63** | |
| 20 | 0.35** | 0.56** | 0.49** | 0.45** | 0.45** | 0.69** | |
| 30 | 0.54** | 0.53** | 0.55** | 0.62** | 0.33* | 0.30* | |
| B | -X | -0.55** | -0.03 | 0.05 | 0.78** | 0.50** | 0.68** |
| 0 | -0.67** | -0.35* | -0.10 | 0.68** | 0.59** | 0.63** | |
| 10 | -0.51** | 0.02 | 0.14 | 0.59** | 0.36** | 0.60** | |
| 20 | -0.59** | 0.02 | -0.07 | 0.74** | 0.40** | 0.57** | |
| 30 | 0.41** | 0.44** | 0.44** | 0.53** | 0.36** | 0.29* |
新窗口打开|下载CSV
3 讨论
3.1 品种、密度和氮肥与冬小麦碳代谢的关系
叶片光合产物主要以蔗糖的形式存在并向籽粒等部位运输,而磷酸蔗糖合成酶(SPS)、蔗糖合成酶(SS)、蔗糖-6-磷酸酯酶(Suc-6-Pase)和蔗糖酶(SC)共同协调蔗糖的合成与降解,其中SPS是促进蔗糖合成的关键酶,尤其在灌浆期,旗叶蔗糖含量与SPS活性一致[27,28,29,30]。适量配施氮肥有利于提高籽粒中蔗糖含量[31],在一定范围内增施氮肥有利于提高旗叶净光合速率和SPS酶活性,促进可溶性总糖、蔗糖的合成[32],而氮肥过量会降低籽粒蔗糖含量,打破碳氮平衡[9],且高氮条件下非气孔因素会影响光合产物合成与运转[32];过量施氮造成穗数增多、群体过大,群体间CO2浓度降低,进而影响旗叶碳代谢循环[33,34]。本研究结果发现,施氮量对冬小麦碳代谢的调控效应是相对的,其调控效果在不同生育时期存在差异,240 kg·hm-2适量施氮处理的碳代谢优势主要体现在灌浆后期。另外,相同种植密度条件下,分蘖力中等、成穗率较高的鑫华麦818在施氮量0—240 kg·hm-2范围内其灌浆期可溶性糖含量、SPS酶活性和SS酶活性均随施氮量的增加而增加,而分蘖力强、成穗率中等的百农207碳代谢生理参数自孕穗至灌浆中期均以不施氮处理最大,但灌浆后期的氮肥调控优势较强,花后30 d的N240仍维持较高的碳代谢生理参数,这可能是因为百农207分蘖力强、源端供应充足,施氮量对其生育前期碳代谢影响较小。本研究结果还表明,鑫华麦818在孕穗期和灌浆后期以中密度M2的碳代谢生理参数最大,百农207亦于灌浆中后期M2的碳代谢参数值最大。因此,适宜的种植密度有利于塑造群体生长起点,协调植株碳氮代谢平衡,调节分蘖与成穗两极分化的矛盾,培育高质量群体生产结构,但种植密度过高则会降低分蘖成穗率,难以实现高产。在本试验条件下,两年度2个品种在灌浆后期均以M2N240氮密处理的碳代谢参数值最大,究其原因可能是由于灌浆后期中密适氮组合具有良好的群体生产结构,田间通风透光条件较好,从而有利于促进叶绿素合成,提高光能转化效率,延缓植株衰老。3.2 品种、密度和氮肥与冬小麦氮代谢的关系
NR、GS分别是氮同化、氨同化过程的2种氮代谢关键酶,共同影响小麦蛋白质含量的高低及品质的好坏。杜少勇等[35]研究表明,增施氮肥可显著增加冬小麦营养器官和籽粒中的氮素积累量,显著提高籽粒的GS酶活性。本研究结果与其相似,即施氮可显著提高鑫华麦818和百农207 2个冬小麦旗叶的可溶性蛋白质含量、NR和GS酶活性。这可能是功能旗叶氮代谢能力的提高有利于促进茎秆、籽粒氮素的积累,进而延缓冬小麦植株衰老。有研究证实,适量施氮有利于促进冬小麦氨同化,增强叶片氮素的有序运转能力,提高氮素运转速率、氮素利用效率和对籽粒的贡献率[36,37]。本文研究亦发现,适量施氮N240的氮代谢优势主要体现在灌浆中期,且对2个品种发挥调控作用的生育时期存在差异,分蘖力强、成穗率中等的百农207除灌浆前期外,孕穗、开花、灌浆中后期的3个氮代谢生理指标参数均随施氮量的增加呈先增后降的变化趋势;而对分蘖力中等、成穗率较高的鑫华麦818,适量施氮N240主要增强了该品种灌浆中期的3个氮代谢生理参数。有研究表明,高密度较低密度种植处理有利于冬小麦氮素积累量增加和产量提高[19],但沈学善等[38]结合播期对种植密度的研究发现,不同播期种植条件下冬小麦蛋白质含量和产量均以中密度处理(225 万株/hm2)最高。本研究结果亦表明,2个品种灌浆中期的氮代谢生理参数均以中密度M2最大。由此表明,适宜的氮密组合可有效提高冬小麦灌浆中期可溶性蛋白质含量、NR和GS酶活性等氮代谢生理参数,进而增强小麦籽粒灌浆强度,提高粒重和产量。3.3 品种、密度和氮肥与冬小麦产量的关系
种植密度过大、施氮量过多虽有利于单位面积穗数增加,但导致顶部与底部小穗的穗粒数减少和中部小穗的千粒重降低,最终影响产量形成[39]。本研究表明,提高种植密度有利于增加冬小麦穗数,同一种植密度下随施氮量的增加,各处理组合的穗数、穗粒数均呈先升后降的单峰变化趋势,但其产量变化在不同密度处理间存在差异,中高密度条件下随施氮量的增加呈先升后降的变化趋势,这与葛君等[14]的研究结果一致,但本研究在低密度种植条件下产量随施氮量的增加而增加,这可能是由于低密度条件下植株群体间竞争较弱,碳代谢差异较小,增加施氮量会显著提高植株氮代谢能力,从而显著提高产量;高密度处理加剧了植株群体和个体竞争,但适宜的氮密组合处理有利于促进碳氮代谢平衡和产量构成三因素协调发展,从而获得高产。另有研究表明,小麦旗叶光合速率、叶绿素含量、SPS酶活性等碳代谢生理参数和籽粒产量变化趋势一致,均随施氮量的增加呈先增后降的变化趋势[14],而本研究表明,碳代谢与穗数、穗粒数、千粒重呈正相关,但随着小麦生育期的推进,碳代谢与产量的相关性呈现“负相关”到“正相关”的转变;氮代谢与产量呈极显著正相关,且主要通过调控穗数来增加产量,不利于千粒重的增加;鑫华麦818在整个灌浆期呈现碳氮代谢与产量正相关,而百农207仅在灌浆后期表现碳氮代谢平衡。因此,鑫华麦818产量构成三因素协调能力高于百农207。研究表明,湿润年份下中密度种植有利于增产,干旱年份下高密度栽培则更有利于提高氮素利用效率而实现增产[20],且灌浆期干旱可加速蔗糖向淀粉的转化速率、缩短灌浆周期、促进早熟[40]。但本研究结果与前人有所差异,2018—2019年正常年份下2个品种均以M2N240处理的产量最高,且碳氮代谢对氮肥、密度的调控效应主要体现在灌浆中后期;而在小麦春季生长期间干旱发生较重的2019—2020年度2个品种均以M1N360处理的产量最高,M2N240处理其次,但2个处理的产量差异不显著。由此可见,冬小麦分蘖两极分化的关键时期出现春旱导致穗数与穗粒数降低、产量下降,但适量增施氮肥、增加种植密度可减缓干旱对产量的负面影响。
4 结论
品种、氮肥、密度及其交互作用显著影响冬小麦碳氮代谢,且各处理组合的碳氮代谢生理参数在分蘖成穗特性不同的2个冬小麦品种间存在差异。氮密调控对冬小麦碳氮代谢的调控作用主要在灌浆中后期,且施氮对小麦产量的调控效应最大。综合考虑品种、氮肥、密度及其交互作用对冬小麦碳氮代谢平衡及产量的影响,种植密度375 万株/hm2与施氮量240 kg·hm-2相配合的M2N240组合可协调植株碳氮代谢平衡,调节分蘖与成穗两极分化的矛盾,促进产量三因素协调发展,进而实现高产,可作为适宜豫东南黏壤潮土区的氮密调控组合。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/pce.v44.4URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s10705-009-9252-0URL [本文引用: 2]
DOI:10.1007/s42106-019-00071-7URL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.fcr.2018.12.011URL [本文引用: 1]
DOI:10.1104/pp.86.4.1013URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/BF00020422URL [本文引用: 1]
DOI:10.17521/cjpe.2003.0025 [本文引用: 1]

环境温度的变化影响植物的碳氮代谢过程,从而改变植物的生长发育。综述了小麦光合产物形成、叶片蔗糖合成、茎鞘中非结构碳水化合物合成与降解、籽粒淀粉合成的一般规律及其在不同温度条件下所发生的变化,指出了小麦灌浆期温度超过30 ℃后粒重和产量将会降低。小麦品质主要由胚乳中面筋蛋白的特性决定,从小麦籽粒蛋白质形成、含量、组分等方面分析了其与灌浆期温度的关系,根据面筋蛋白特性的变化,认为高温胁迫对小麦烘烤品质会产生不利影响。同时全球气候变暖可能加重小麦灌浆期的高温危害,提出了今后相关研究中将要出现的3个热点问题:1)CO2浓度升高和温度升高协同作用对小麦产量和品质的影响,2)小麦耐热性的生理机制及其栽培调控措施,3)筛选小麦耐热基因及选育耐热新品种。
DOI:10.17521/cjpe.2003.0025 [本文引用: 1]
环境温度的变化影响植物的碳氮代谢过程,从而改变植物的生长发育。综述了小麦光合产物形成、叶片蔗糖合成、茎鞘中非结构碳水化合物合成与降解、籽粒淀粉合成的一般规律及其在不同温度条件下所发生的变化,指出了小麦灌浆期温度超过30 ℃后粒重和产量将会降低。小麦品质主要由胚乳中面筋蛋白的特性决定,从小麦籽粒蛋白质形成、含量、组分等方面分析了其与灌浆期温度的关系,根据面筋蛋白特性的变化,认为高温胁迫对小麦烘烤品质会产生不利影响。同时全球气候变暖可能加重小麦灌浆期的高温危害,提出了今后相关研究中将要出现的3个热点问题:1)CO2浓度升高和温度升高协同作用对小麦产量和品质的影响,2)小麦耐热性的生理机制及其栽培调控措施,3)筛选小麦耐热基因及选育耐热新品种。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}