Abstract 【Objective】 Fusarium wilt is one of the most important soil-borne diseases in cotton planting. Chemical methods are mainly used to control this disease, however, it has certain impact on the environment, human and animal safety. Biological control has become an important way to control cotton Fusarium wilt because of its high specificity and safety. The objective of this study is to screen an efficient antagonistic bacteria, and characterize the biocontrol mechanism of bacteria against Fusarium wilt, thus providing a technical basis for cotton Fusarium wilt control with biocontrol bacteria. 【Method】 In a previous study, an endophytic bacterium Bacillus cereus YUPP-10 was isolated from vascular of cotton, which can hydrolyze polysaccharides with β-1,4 linkage. The effects of YUPP-10 on hyphal growth, sporulation, and spore germination of F. oxysporum were tested using the confront culture method, enclosed chamber test and hanging drop method, respectively. The seeds were soaked by YUPP-10, and then, the seeding germination and the biomass of cotton were detected. The cotton were cultivated in substrate with F. oxysporum, and after a week of growth, YUPP-10 cultured with LB liquid medium was treated at different concentrations (1×10 8, 1×107and 1×106 cfu/mL, respectively), and the control efficacy against Fusarium wilt was studied in the greenhouse. The key antibacterial substances of YUPP-10 were obtained by Fosmid library, and the direct effects of recombinant cyclodextrin glycosyltransferase (CGTase) on F. oxysporum hyphal growth, sporulation, and spore germination were studied. The overexpression vector was transformed into Arabidopsis thaliana Col-0 via the floral dip method. The resistance of CGTase-overexpressing transgenic plants against Fusarium wilt was assessed with an in vitro technique. The transcriptional levels of some defense genes were analyzed under pathogen challenge. 【Result】 YUPP-10 significantly inhibited the hyphal growth, sporulation, and spore germination of F. oxysporum, the most inhibition rate of spore yield and germination was 98.41% and 51.65%, respectively. Low concentration of YUPP-10 could promote the germination rate, emergence rate and stem length of cotton seeds. After the treatment of YUPP-10, the diseased plant rate and disease index were significantly lower than those of the control group. The control efficacy was 45.11% at the concentration of 1×10 7 cfu/mL. CGTase was the key antimicrobial substance of YUPP-10, the effects of added CGTase on the transparent circles of carboxymethyl cellulose and glucomannan were measured, the results showed that CGTase could hydrolyze polysaccharides with β-1,4 linkage. CGTase also had significant inhibitory effects on the growth, sporulation and spore germination of the pathogen, the most inhibition rate of spore yield and germination was 62.63% and 30.83%, respectively. The CGTase-overexpressing A. thaliana enhanced disease resistance by enhancing the expression of defense genes. 【Conclusion】 YUPP-10 is an efficient biocontrol agent that inhibits the F. oxysporum growth, promots germination rate, emergence rate and stem length of cotton seeds, and protects cotton plant from F. oxysporum infection. CGTase can inhibit the growth of F. oxysporum, and its transgenic A. thaliana enhances the resistance to Fusarium wilt. Keywords:Bacillus cereus;cotton Fusarium wilt;Fusarium oxysporum f. sp. vasinfectum;cyclodextrin glycosyltransferase (CGTase);control efficacy
PDF (2644KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 周京龙, 冯自力, 魏锋, 赵丽红, 张亚林, 周燚, 冯鸿杰, 朱荷琴. 棉花内生细菌YUPP-10及其分泌蛋白CGTase对棉花枯萎病的防治作用及机理. 中国农业科学, 2021, 54(17): 3691-3701 doi:10.3864/j.issn.0578-1752.2021.17.011 ZHOU JingLong, FENG ZiLi, WEI Feng, ZHAO LiHong, ZHANG YaLin, ZHOU Yi, FENG HongJie, ZHU HeQin. Biocontrol Effect and Mechanism of Cotton Endophytic Bacterium YUPP-10 and Its Secretory Protein CGTase Against Fusarium Wilt in Cotton. Scientia Acricultura Sinica, 2021, 54(17): 3691-3701 doi:10.3864/j.issn.0578-1752.2021.17.011
Table 4 表4 表4YUPP-10菌剂处理对棉花枯萎病病株率、病情指数的影响及防治效果 Table 4Effects of YUPP-10 bacterial fertilizer on diseased plant rate, disease index and its efficacy in control Fusarium wilt
SANOGOS, ZHANGJ. Resistance sources, resistance screening techniques and disease management for Fusarium wilt in cotton Euphytica, 2016, 207(2): 255-271. DOI:10.1007/s10681-015-1532-yURL [本文引用: 1]
ZHANGZ, DIAOH, WANGH, WANGK, ZHAOM. Use of Ganoderma lucidum polysaccharide to control cotton fusarium wilt, and the mechanism involved Pesticide Biochemistry and Physiology, 2019, 158: 149-155. DOI:10.1016/j.pestbp.2019.05.003URL [本文引用: 1]
LANGJ, HUJ, RANW, XUY, SHENQ. Control of cotton Verticillium wilt and fungal diversity of rhizosphere soils by bio-organic fertilizer Biology and Fertility of Soils, 2012, 48(2): 191-203. DOI:10.1007/s00374-011-0617-6URL [本文引用: 1]
LIS Z, LUX Y, MAP, GAOS G, LIUX Z, LIUG. Evaluation of biocontrol potential of a bacterial strain NCD-2 against cotton Verticillium wilt in field trials Acta Phytopathologica Sinica, 2005, 35(5): 451-455. (in Chinese) [本文引用: 1]
LIB, LIQ, XUZ, ZHANGN, SHENQ, ZHANGR. Responses of beneficial Bacillus amyloliquefaciens SQR9 to different soilborne fungal pathogens through the alteration of antifungal compounds production Frontiers in Microbiology, 2014, 5: 636. [本文引用: 1]
PEREGL, MCMILLANM. Scoping the potential uses of beneficial microorganisms for increasing productivity in cotton cropping systems Soil Biology and Biochemistry, 2015, 80: 349-358. DOI:10.1016/j.soilbio.2014.10.020URL [本文引用: 1]
LIH W, HANW L, WANGM Y, LUOM, HANJ, GUA X. Screening and identification of antagonistic bacteria Bacillus pumilus against cotton Fusarium wilt and biocontrol research Chinese Journal of Biological Control, 2018, 34(3): 440-448. (in Chinese) [本文引用: 1]
LIUZ Q, ZHANGB R, LIUY, GUON N, GUA X. Effect of Bacillus pumilus KX-33 seed coating agent on Verticillium wilt of cotton Journal of Xinjiang Agricultural University, 2020, 43(5): 323-329. (in Chinese) [本文引用: 1]
DAIP B, LANX J, ZHANGW W, GANL, WANGY, ZONGZ F. Identification, colonization and disease suppressive effect of strain SC11 against cotton Fusarium wilt Acta Phytopathologica Sinica, 2016, 46(2): 273-279. (in Chinese) [本文引用: 1]
PLEBANS, CHERNINL, CHETI. Chitinolytic activity of an endophytic strain of Bacillus cereus Letters in Applied Microbiology, 1997, 25(4): 284-288. DOI:10.1046/j.1472-765X.1997.00224.xURL [本文引用: 1]
LIJ G, JIANGZ Q, XUL P, SUNF F, GUOJ H. Characterization of chitinase secreted by Bacillus cereus strain CH2 and evaluation of its efficacy against Verticillium wilt of eggplant BioControl, 2008, 53(6): 931-944. DOI:10.1007/s10526-007-9144-7URL [本文引用: 1]
KISHOREG K, PANDES. Chitin-supplemented foliar application of chitinolytic Bacillus cereus reduces severity of Botrytis gray mold disease in chickpea under controlled conditions Letters in Applied Microbiology, 2007, 44(1): 98-105. DOI:10.1111/lam.2007.44.issue-1URL [本文引用: 1]
HAMMAMII, SIALAR, JRIDIM, KTARIN, NASRIM, TRIKIM A. Partial purification and characterization of chiIO8, a novel antifungal chitinase produced by Bacillus cereus IO8 Journal of Applied Microbiology, 2013, 115(2): 358-366. DOI:10.1111/jam.12242URL [本文引用: 1]
ZHOUJ L, FENGZ L, FENGH J, LIY Q, YUANY, LIZ F, WEIF, SHIY Q, ZHAOL H, SUNZ X, ZHUH Q, ZHOUY. Biocontrol effect and mechanism of cotton endophytic bacterium Bacillus cereus YUPP-10 against Verticillium wilt in Gossypium hirsutum Scientia Agricultura Sinica, 2017, 50(14): 2717-2727. (in Chinese) [本文引用: 2]
LIS K, JIZ Q, ZHANGJ W, GUOZ Y, WUW J. Synthesis of 1-acyl-3-isopropenylbenzimidazolone derivatives and their activity against Botrytis cinerea Journal of Agricultural and Food Chemistry, 2010, 58(5): 2668-2672. DOI:10.1021/jf903855yURL [本文引用: 1]
ZHUH Q, FENGZ L, LIZ F, ZHAOL H, SHIY Q. Dip quantitative fungal solution method by using vermiculite sand without bottom paper bowl to identify cotton varieties (lines) of resistance to Verticillium wilt China Cotton, 2010, 37(12): 15-17. (in Chinese) [本文引用: 1]
CLOUGHS J, BENTA F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana The Plant Journal, 1998, 16(6): 735-743. DOI:10.1046/j.1365-313x.1998.00343.xURL [本文引用: 1]
MARASCOR, ROLLIE, ETTOUMIB, VIGANIG, MAPELLIF, BORINS, ABOU-HADIDA F, EL-BEHAIRYU A, SORLINIC, CHERIFA, ZOCCHIG, DAFFONCHIOD. A drought resistance- promoting microbiome is selected by root system under desert farming PLoS ONE, 2012, 7(10): e48479. DOI:10.1371/journal.pone.0048479URL [本文引用: 1]
RASHIDS, CHARLEST C, GLICKB R. Isolation and characterization of new plant growth-promoting bacterial endophytes Applied Soil Ecology, 2011, 61(5): 217-224. DOI:10.1016/j.apsoil.2011.09.011URL [本文引用: 1]
HARDOIMP R, VANOVERBEEK L S, VANELSAS J D. Properties of bacterial endophytes and their proposed role in plant growth Trends in Microbiology, 2008, 16(10): 463-471. DOI:10.1016/j.tim.2008.07.008URL [本文引用: 1]
ALIS, CHARLEST C, GLICKB R. Delay of flower senescence by bacterial endophytes expressing 1-aminocyclopropane-1-carboxylate deaminase Journal of Applied Microbiology, 2012, 113(5): 1139-1144. DOI:10.1111/jam.2012.113.issue-5URL [本文引用: 1]
KATSURAYAK, OKUYAMAK, HATANAKAK, OSHIMAR, SATOT, MATSUZAKIK. Constitution of konjac glucomannan: Chemical analysis and 13C NMR spectroscopy Carbohydrate Polymers, 2003, 53(2): 183-189. DOI:10.1016/S0144-8617(03)00039-0URL [本文引用: 1]
FANGW, WUP. Variations of konjac glucomannan (KGM) from Amorphophallus konjac and its refined powder in China Food Hydrocolloids, 2004, 18(1): 167-170. DOI:10.1016/S0268-005X(03)00044-4URL [本文引用: 1]
LAFARGEC, CAYOTN, HORYC, GONCALVESL, CHASSEMONTC, LEBAIL P. Effect of konjac glucomannan addition on aroma release in gels containing potato starch Food Research International, 2014, 64: 412-419. DOI:10.1016/j.foodres.2014.07.008URL [本文引用: 1]
PANGBURNS H, TRESCONYP V, HELLERJ. Lysozyme degradation of partially deacetylated chitin, its films and hydrogels Biomaterials, 1982, 3(2): 105-108. DOI:10.1016/0142-9612(82)90043-6URL [本文引用: 1]
CHENJ, PENGH, WANGX, SHAOF, YUANZ, HANH. Graphene oxide exhibits broad-spectrum antimicrobial activity against bacterial phytopathogens and fungal conidia by intertwining and membrane perturbation Nanoscale, 2014, 6(3): 1879-1889. DOI:10.1039/C3NR04941HURL [本文引用: 1]
NEJADP, JOHNSONP A. Endophytic bacteria induce growth promotion and wilt disease suppression in oilseed rape and tomato Biological Control, 2000, 18(3): 208-215. DOI:10.1006/bcon.2000.0837URL [本文引用: 1]
ABDALLAHR A B, MOKNI-TLILIS, NEFZIA, JABNOUN-KHIAREDDINE H, DAAMI-REMADI M. Biocontrol of Fusarium wilt and growth promotion of tomato plants using endophytic bacteria isolated from Nicotiana glauca organs Biological Control, 2016, 97(10): 80-88. DOI:10.1016/j.biocontrol.2016.03.005URL [本文引用: 1]
AJILOGBAC F. RAPD delineation of native Bacillus spp. and analyses of their biocontrol effect on tomato Fusarium wilt [D]. Mafikeng Campus of the North-West University, 2013. [本文引用: 1]
ABDALLAHR A, NEU O, JABNOUN-KHIAREDDINEH, MOKNI-TLILIS, NEFZIA, MEDIMAGH-SAIDANAS, DAAMI-REMADI M. Endophytic Bacillus spp. from wild Solanaceae and their antifungal potential against Fusarium oxysporum f. sp. lycopersici elucidated using whole cells, filtrate cultures and organic extracts Journal of Plant Pathology and Microbiology, 2015, 6: 11. [本文引用: 1]
FUY, HUANGY Y, XIAOA Q, YOUC P. Preliminary study on the resistant-related enzymes from banana by the induction of Bacillus cereus strain 255 Guangdong Agricultural Sciences, 2011, 38(9): 75-77. (in Chinese) [本文引用: 1]
ZHOUJ, FENGZ, LIUS, WEIF, SHIY, ZHAOL, HUANGW, ZHOUY, FENGH, ZHUH. CGTase, a novel antimicrobial protein from Bacillus cereus YUPP-10, suppresses Verticillium dahliae and mediates plant defence responses Molecular Plant Pathology, 2021, 22(1): 130-144. DOI:10.1111/mpp.13014URL [本文引用: 1]
GUOX, STOTZH U. Defense against Sclerotinia sclerotiorum in Arabidopsis is dependent on jasmonic acid, salicylic acid, and ethylene signaling Molecular Plant-Microbe Interactions, 2007, 20(11): 1384-1395. DOI:10.1094/MPMI-20-11-1384URL [本文引用: 1]
GLAZEBROOKJ, CHENW, ESTESB, CHANGH S, NAWRATHC, MÉTRAUXJ P, ZHUT, KATAGIRIF. Topology of the network integrating salicylate and jasmonate signal transduction derived from global expression phenotyping The Plant Journal, 2003, 34(2): 217-228. DOI:10.1046/j.1365-313X.2003.01717.xURL [本文引用: 1]
 ,1,2, 冯自力2, 魏锋2, 赵丽红2, 张亚林2, 周燚
,1,2, 冯自力2, 魏锋2, 赵丽红2, 张亚林2, 周燚

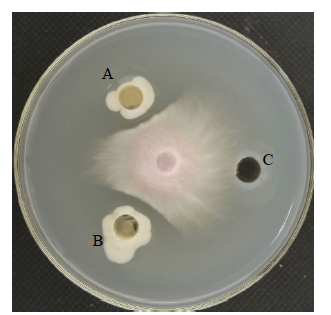 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT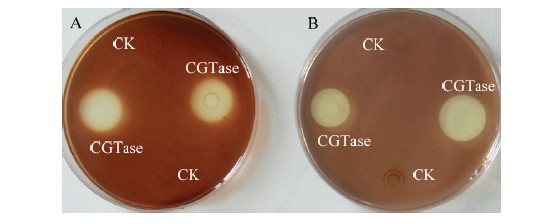 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT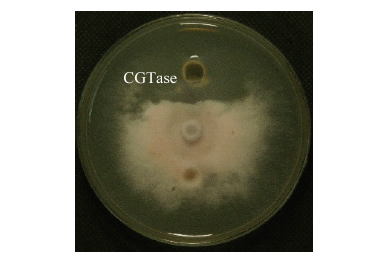 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}