Identification and Virulence Analysis of CAP Superfamily Genes in Valsa mali
WANG ChengLi,, YIN ZhiYuan, NIE JiaJun, LIN YongHui, HUANG LiLi,College of Plant Protection, Northwest A&F University/State Key Laboratory of Crop Stress Biology for Arid Areas, Yangling 712100, Shaanxi
Received:2020-11-8Accepted:2020-12-17 作者简介 About authors 王程利,E-mail: wein599@163.com。
摘要 【目的】CAP(Cysteine-rich secretory protein,Antigen 5 and Pathogenesis related protein 1)超家族蛋白广泛存在于真菌、细菌、动植物等物种,并且参与病原菌的致病过程。本研究旨在鉴定苹果树腐烂病致病菌——苹果黑腐皮壳菌(Valsa mali)的CAP超家族基因并明确CAP超家族基因在病菌致病方面的作用。【方法】通过BLASTP在苹果黑腐皮壳菌全基因组中检索具有CAP保守结构域的基因;利用特异性引物对鉴定到的CAP基因进行PCR扩增和凝胶电泳检测;使用生物信息学软件和在线数据库进行蛋白序列特征和系统发育分析;利用RT-qPCR分析基因在病菌侵染过程中的表达模式;使用特异性引物扩增CAP基因上、下游片段并从PDL2质粒上扩增筛选标记基因;利用Double-joint PCR构建基因敲除盒并通过PEG介导的原生质转化技术进行基因敲除和基因回补;以遗传霉素(G418)抗性为筛选标记并利用4对引物PCR检测获得基因敲除突变体;以潮霉素(HPH)抗性为筛选标记获得基因回补菌株;通过对菌株进行培养皿内生长试验明确基因对病菌营养生长的影响;通过离体苹果枝条接种试验分析该病菌CAP超家族基因的毒性功能。【结果】在苹果黑腐皮壳菌中鉴定到3个具有CAP保守结构域的基因,分别命名为VmPR1a、VmPR1b和VmPR1c。序列特征分析发现,3个CAP蛋白均包含4个保守区:N端信号肽、N端延伸区(NTE)、CAP保守功能域和C端延伸区(CTE)。系统发育分析显示,3个CAP蛋白聚于不同的进化支,VmPR1a聚于clade2进化支并与粗糙链孢霉(Neurospora crassa)CAP蛋白进化关系较近;VmPR1b聚于clade3且与镰孢菌属(Fusarium spp.)CAP蛋白的进化关系更近;VmPR1c聚于clade1,并且也与粗糙链孢霉的CAP蛋白进化关系较近。RT-qPCR分析结果显示,VmPR1a、VmPR1b和VmPR1c在病菌侵染早期(6 h和12 h)均显著上调表达。利用PEG遗传转化技术获得VmPR1a、VmPR1b和VmPR1c敲除突变体(ΔVmPR1a-7/23、ΔVmPR1b-20/31和ΔVmPR1c-26/40);营养生长观察发现,所有敲除突变体生长表型与野生型菌株03-8均无明显差异;致病力检测发现,ΔVmPR1b-20/31致病力较野生型无明显变化,而ΔVmPR1a-7/23和ΔVmPR1c-26/40致病力较野生型显著下降。将VmPR1a和VmPR1c分别回补至ΔVmPR1a和ΔVmPR1c,回补菌株(VmPR1a/C和VmPR1c/C)致病力恢复至野生型水平。【结论】苹果黑腐皮壳菌中存在3个CAP超家族基因(VmPR1a、VmPR1b和VmPR1c),其中VmPR1a和VmPR1c是苹果黑腐皮壳菌重要的毒性因子。 关键词:苹果黑腐皮壳菌;CAP 蛋白;基因鉴定;基因敲除;毒性功能
Abstract 【Objective】CAP (Cysteine-rich secretory protein, Antigen 5 and Pathogenesis related protein 1) superfamily proteins widely exist in fungi, bacteria, animal and plant. This kind of proteins participates in the pathogenic process of the pathogen. The purpose of this study was to identify the CAP superfamily genes in Valsa mali and clarify their virulence roles.【Method】BLASTP was used to retrieve genes with the conserved CAP domains in the whole genome of V. mali. PCR amplification and gel electrophoresis detection were carried out with specific primers. Bioinformatics software and online databases were used for protein sequence characterization and phylogenetic analysis. RT-qPCR was used to analyze the gene expression profiles. Double-joint PCR was used to construct gene knocking-out cassettes, PEG-mediated protoplast transformation was used to obtain knocking-out mutants and complementation strains. To obtain gene knockout mutants, geneticin (G418) was used as a selection marker, and transformants were validated by four pairs of primers. To obtain gene complementation transformants, hygromycin (HPH) was used as a selection marker. The vegetative growth of these strains was determined by cultivation on PDA medium, and the virulence of these strains was verified by inoculation on apple twigs.【Result】Three CAP superfamily genes were identified in V. mali, named VmPR1a, VmPR1b and VmPR1c, respectively. The three CAP proteins all contain four conserved regions, including a N-terminal signal peptide, a N-terminal extension region (NTE), a CAP domain and a C-terminal extension region (CTE). Phylogenetic analysis showed that the three proteins belonged to different clades. VmPR1a was clustered in clade2 and closely related to Neurospora crassa CAP protein. VmPR1b was clustered in clade3 and closely related to CAP proteins of Fusarium spp.. VmPR1c was clustered in clade1 and also closely related to N. crassa CAP protein. RT-qPCR analysis showed that VmPR1a, VmPR1b and VmPR1c were significantly up-regulated during early stages of infection (6 h and 12 h). VmPR1a, VmPR1b and VmPR1c knockout mutants (ΔVmPR1a-7/23, ΔVmPR1b-20/31 and ΔVmPR1c-26/40, respectively) were obtained, and all mutants showed no apparent alteration in filamentous growth compared with that of the wild-type strain. The virulence of ΔVmPR1b-20/31 was not obviously influenced. However, the virulence of ΔVmPR1a-7/23 and ΔVmPR1c-26/40 was significantly reduced compared with that of the wild-type strain. Moreover, the virulence of gene complementation strains (VmPR1a/C and VmPR1c/C) was restored to comparable level as that of the wild-type. 【Conclusion】Three CAP super-family genes were identified in V. mali (VmPR1a, VmPR1b and VmPR1c), and VmPR1a and VmPR1c are virulence factors of V. mali. Keywords:Valsa mali;CAP protein;gene identification;gene knockout;virulence
PDF (3096KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 王程利, 尹志远, 聂嘉俊, 林永辉, 黄丽丽. 苹果黑腐皮壳菌CAP超家族蛋白基因鉴定及毒性功能分析[J]. 中国农业科学, 2021, 54(16): 3440-3450 doi:10.3864/j.issn.0578-1752.2021.16.007 WANG ChengLi, YIN ZhiYuan, NIE JiaJun, LIN YongHui, HUANG LiLi. Identification and Virulence Analysis of CAP Superfamily Genes in Valsa mali[J]. Scientia Acricultura Sinica, 2021, 54(16): 3440-3450 doi:10.3864/j.issn.0578-1752.2021.16.007
试剂:RNA提取试剂盒(华越洋)、反转录试剂盒(Thermo)、Phanta® Super Fidelity DNA Polymerase(Vazyme)、限制性内切酶(Thermo)、Clone Express® Entry One Step CloningKit(Vazyme);卡那霉素(Solarbio)、潮霉素B(Roche)、SYBR Green real-time PCR Master Mix(Genestar)、遗传霉素(MP);核酸Marker DL2000、DL5000(TaKaRa);DNA凝胶回收试剂盒、质粒小量提取试剂盒(OMEGA);大肠杆菌感受态DH5α(全式金);崩溃酶(Sigma)和溶壁酶(Sigma)。引物合成及测序由生工生物工程股份有限公司完成。
1.2 方法
1.2.1 生物信息学分析 根据CAP超家族蛋白的典型特征,使用BLASTP检索苹果黑腐皮壳菌基因组数据库,并用HMMER 3.0软件( http://hmmer.janelia.org/)筛选具有CAP结构域的氨基酸序列。初筛获得的氨基酸序列输入在线数据库NCBI CDD web server( https://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml)分析蛋白结构域特征并进行蛋白功能注释。使用DNAMAN 6.0.3.9对序列比对分析;使用Protein isoelectric point calculator web server( http://isoelectric.ovh.org/index.html)预测蛋白质分子量和等电点;SignalP 4.0在线预测工具( http://www.cbs.dtu.dk/services/SignalP-4.0/)预测蛋白信号肽;跨膜结构域的预测使用TMHMM Server v. 2.0( https://services.healthtech.dtu.dk/)在线数据库;糖基化修饰位点使用NetNGlyc 1.0 Server( http://www.cbs.dtu.dk/services/NetNGlyc/)在线预测;蛋白泛素化(Ubiquitination)修饰位点的预测使用GPS-SUMO 2.0 Online Service在线软件( http://sumosp.biocuckoo.org/online.php)。从NCBI数据库中下载苹果黑腐皮壳菌CAP家族蛋白同源序列并使用MEGA 7构建系统发育树。
野生型WT:03-8;Marker:DL2000;1—4:分别指各基因的4对检测引物(5F/6R、7F/8R、9F/10R和Neo-F/R)的PCR产物检测 PCR amplification product detection of 5F/6R, 7F/8R, 9F/10R and Neo-F/R for each gene。A:ΔVmPR1a-7和ΔVmPR1a-23突变体及回补菌株PCR检测 PCR amplification product detection of ΔVmPR1a-7, ΔVmPR1a-23 and complementation strain;引物VmPR1a-5F/6R检测回补菌株ΔVmPR1a/C The complementation strain ΔVmPR1a/C was detected by primer VmPR1a-5F/6R。B:ΔVmPR1b-20和ΔVmPR1b-31突变体PCR检测 PCR amplification product detection of ΔVmPR1b-20 and ΔVmPR1b-31。C:ΔVmPR1c-26和ΔVmPR1c-40突变体及回补菌株PCR检测 PCR amplification product detection of ΔVmPR1c-26, ΔVmPR1c-40 and complementation strain;引物VmPR1c-5F/6R检测回补菌株ΔVmPR1c/C The complementation strain ΔVmPR1c/C was detected by primer VmPR1c-5F/6R Fig. 6PCR detection of mutants and complementation strains
GIBBS GM, ROELANTSK, O’BRYAN MK. The CAP superfamily: Cysteine-rich secretory proteins, antigen 5, and pathogenesis-related 1 proteins—roles in reproduction, cancer, and immune defense Endocrine Reviews, 2008, 29(7):865-897. DOI:10.1210/er.2008-0032URL [本文引用: 1]
OLRICHS NK, HELMS JB. Novel insights into the function of the conserved domain of the CAP superfamily of proteins AIMS Biophysics, 2016, 3(2):232-246. DOI:10.3934/biophy.2016.2.232URL [本文引用: 1]
KELLEHERA, DARWICHER, REZENDE WC, FARIAS LP, LEITE L CC, SCHNEITERR, ASOJO OA. Schistosoma mansoni venom allergen-like protein 4 (SmVAL4) is a novel lipid-binding SCP/TAPS protein that lacks the prototypical CAP motifs Acta Crystallographica, 2014, 71(8):2186-2196. [本文引用: 1]
MILNE TJ, ABBENANTEG, TYNDALL J DA, HALLIDAYJ, LEWIS RJ. Isolation and characterization of a cone snail protease with homology to CRISP proteins of the pathogenesis-related protein superfamily Journal of Biological Chemistry, 2003, 278(33):31105-31110. DOI:10.1074/jbc.M304843200URL [本文引用: 1]
KING TP, MORAND, WANG DF, KOCHOUMIANL, CHAIT BT. Structural studies of a hornet venom allergen antigen 5, Dol m V and its sequence similarity with other proteins Protein Sequences and Data Analysis, 1990, 3(3):263-266. [本文引用: 1]
VAN LOON LC, VAN KAMMENA. Polyacrylamide disc electrophoresis of the soluble leaf proteins from Nicotiana tabacum var. ‘Samsun’ and ‘Samsun NN’: II Changes in protein constitution after infection with tobacco mosaic virus Virology, 1970, 40:190-211. [本文引用: 1]
CHEN YL, LEE CY, CHENG KT, CHANG WH, HUANG RN, NAM HG, CHEN YR. Quantitative peptidomics study reveals that a wound-induced peptide from PR-1 regulates immune signaling in tomato The Plant Cell, 2014, 26:4135-4148. DOI:10.1105/tpc.114.131185URL [本文引用: 2]
ROHMM, LINDEMANNE, HILLERE, ERMERTD, LEMUTHK, TRKULJAD, SOGUKPINARO, BRUNNERH, RUPPS, URBAN CF, SOHNK. A family of secreted pathogenesis-related proteins in Candida albicans Molecular Microbiology, 2013, 87(1):132-151. DOI:10.1111/mmi.2013.87.issue-1URL [本文引用: 1]
PRADOS-ROSALES RC, ROLDAN-RODRIGUEZR, SERENAC, LOPEZ-BERGES MS, GUARROJ, MARTÍNEZ-DEL-POZOÁ, DI PIETROA. A pr-1-like protein of Fusarium oxysporum functions in virulence on mammalian hosts Journal of Biological Chemistry, 2012, 287(26):21970-21979. DOI:10.1074/jbc.M112.364034URL [本文引用: 1]
TEIXEIRA P JP, THOMAZELLA D PT, VIDAL RO, DO PRADO P FV, REISO, BARONI RM, FRANCO SF, MIECZKOWSKIP, PEREIRA G AG, MONDEGO J MC. The fungal pathogen Moniliophthora perniciosa has genes similar to plant PR-1 that are highly expressed during its interaction with cacao PLoS ONE, 2012, 7(9):e45929. DOI:10.1371/journal.pone.0045929URL [本文引用: 2]
LUS, EDWARDS MC. Molecular characterization and functional analysis of PR-1-like proteins identified from the wheat head blight fungus Fusarium graminearum. Phytopathology, 2018, 108(4):510-520. DOI:10.1094/PHYTO-08-17-0268-RURL [本文引用: 2]
YIN ZY, LIU HQ, LI ZP, KE XW, DOU DL, GAO XN, SONGN, DAI QQ, WU YX, XU JR, KANG ZS, HUANG LL. Genome sequence of Valsa canker pathogens uncovers a potential adaptation of colonization of woody bark New Phytologist, 2015, 208(4):1202-1216. DOI:10.1111/nph.2015.208.issue-4URL [本文引用: 1]
WANG XL. Pathogen of apple tree valsa canker in China: A combined analysis of phenotypic characteristics and rDNA-ITS sequences [D]. Yangling: Northwest A&F University, 2007. (in Chinese) [本文引用: 1]
KEX, HUANG LL, HAN QM, GAO XN, KANG ZS. Histological and cytological investigations of the infection and colonization of apple bark by Valsa mali var. mali Australasian Plant Pathology, 2013, 42(1):85-93. DOI:10.1007/s13313-012-0158-yURL [本文引用: 1]
DU ZT, LI ZP, GAO XN, HUANG LL, HAN QM. Study on the conidia dispersal and the disease dynamics of apple tree canker caused by Valsa mali var. mali in Shaanxi Journal of Fruit Science, 2013, 30(5):819-822. (in Chinese) [本文引用: 1]
LINX, SUN CR, WANG CX, LIANS, DONG XL, LI BH. Epidemic factors affecting the infection and occurrence of Valsa mali Scientia Agricultura Sinica, 2021, 54(11):2333-2342. (in Chinese) [本文引用: 1]
BESSHOH, TSUCHIYAS, SOEJIMAJ. Screening methods of apple trees for resistance to Valsa canker Euphytica, 1994, 77(1/2):15-18. DOI:10.1007/BF02551454URL [本文引用: 1]
ABEK, KOTODAN, KATOH, SOEJIMAJ. Genetic studies on resistance to Valsa canker in apple: Genetic variance and breeding values estimated from intra- and inter-specific hybrid progeny populations Tree Genetics and Genomes, 2011, 7(2):363-372. DOI:10.1007/s11295-010-0337-3URL [本文引用: 1]
LIU XY, LÜS, WANGY, WANGK, LI TH, HAN ZH, ZHANG XH. Evaluation of resistance of Malus germplasms to apple canker (Valsa ceratosperma) Journal of Fruit Science, 2011, 28(5):843-848. (in Chinese) [本文引用: 1]
YIN ZY, KE XW, HUANG DX, GAO XN, VOEGELE RT, KANG ZS, HUANG LL. Validation of reference genes for gene expression analysis in Valsa mali var. mali using real-time quantitative PCR World Journal of Microbiology and Biotechnology, 2013, 29(9):1563-1571. DOI:10.1007/s11274-013-1320-6URL [本文引用: 1]
YUJ, HAMARIZ, HANK, SEOJ, REYES-DOMÍNGUEZY, SCAZZOCCHIOC. Double-joint PCR: A PCR-based molecular tool for gene manipulations in filamentous fungi Fungal Genetics and Biology, 2004, 43:973-981. [本文引用: 1]
GAOJ, LI YB, KE XW, KANG ZS, HUANG LL. Development of genetic transformation system of Valsa mali of apple mediated by PEG Acta Microbiologica Sinica, 2011, 51(9):1194-1199. (in Chinese) [本文引用: 1]
ZANGR, HUANG LL, KANG ZS, WANG XL. Biological characteristics and pathogenicity of different isolates of Cytospora spp. isolated from apple trees in Shaanxi Province Acta Phytopathologica Sinica, 2007, 37(4):343-351. (in Chinese) [本文引用: 1]
XU CJ. Pathogenic function of three pectinase genes in Valsa mali [D]. Yangling: Northwest A&F University, 2016. (in Chinese) [本文引用: 1]
WU YX, XU LS, YIN ZY, DAI QQ, GAO XN, FENGH, VOEGELE RT, HUANG LL. Two members of the velvet family, VmVeA and VmVelB, affect conidiation, virulence and pectinase expression in Valsa mali Molecular Plant Pathology, 2018, 19(7):1639-1651. DOI:10.1111/mpp.2018.19.issue-7URL [本文引用: 1]
FENGH, XUM, ZHENGX, ZHU TY, GAO XN, HUANG LL. MicroRNAs and their targets in apple (Malus domestica cv. “Fuji”) involved in response to infection of pathogen Valsa mali Frontiers in Plant Science, 2017, 8:2081. DOI:10.3389/fpls.2017.02081URL [本文引用: 1]
XUM, GUOY, TIAN RZ, GAOC, GUO FR, VOEGELE RT, BAO JY, LI CJ, JIA CH, FENGH, HUANG LL. Adaptive regulation of virulence genes by microRNA-like RNAs in Valsa mali New Phytologist, 2020, 227:899-913. DOI:10.1111/nph.v227.3URL [本文引用: 1]
VLEESHOUWERS VG, OLIVER RP. Effectors as tools in disease resistance breeding against biotrophic, hemibiotrophic, and necrotrophic plant pathogens Molecular Plant-Microbe Interactions, 2014, 27(3):196-206. DOI:10.1094/MPMI-10-13-0313-IAURL [本文引用: 1]
LIZ, YIN ZY, FANY, XUM, KANG ZS, HUANG LL. Candidate effector proteins of the necrotrophic apple canker pathogen Valsa mali can suppress BAX-induced PCD Frontiers in Plant Science, 2015, 6:579. [本文引用: 1]
ZHANGM, FENGH, ZHAO YH, SONG LL, GAOC, XU XM, HUANG LL. Valsa mali pathogenic effector VmPxE1 contributes to full virulence and interacts with the host peroxidase MdAPX1 as a potential target Frontiers in Microbiology, 2018, 9:821. DOI:10.3389/fmicb.2018.00821URL [本文引用: 1]
ZHANGM, XIE SC, ZHAO YH, MENGX, SONG LL, FENGH, HUANG LL. Hce2 domain-containing effectors contribute to the full virulence of Valsa mali in a redundant manner Molecular Plant Pathology, 2019, 20(6):843-856. DOI:10.1111/mpp.2019.20.issue-6URL [本文引用: 1]
 ,, 尹志远, 聂嘉俊, 林永辉, 黄丽丽
,, 尹志远, 聂嘉俊, 林永辉, 黄丽丽

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT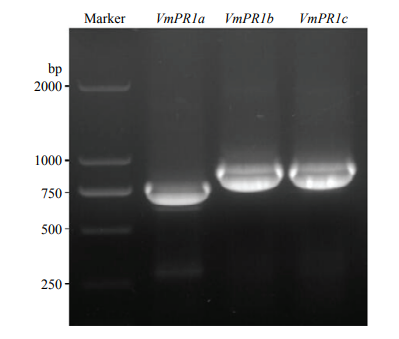 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT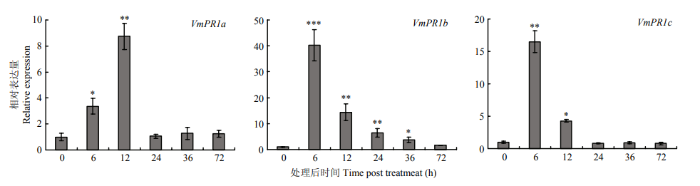 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT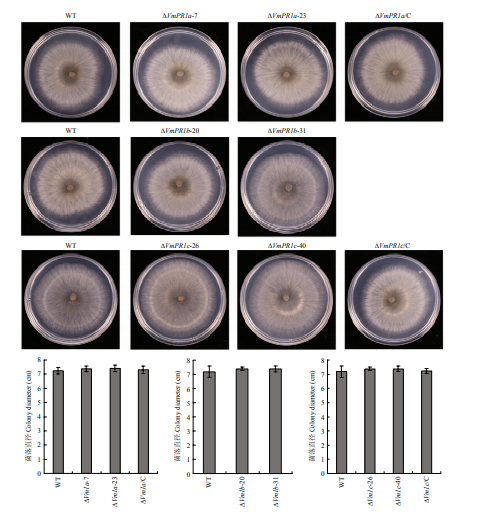 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT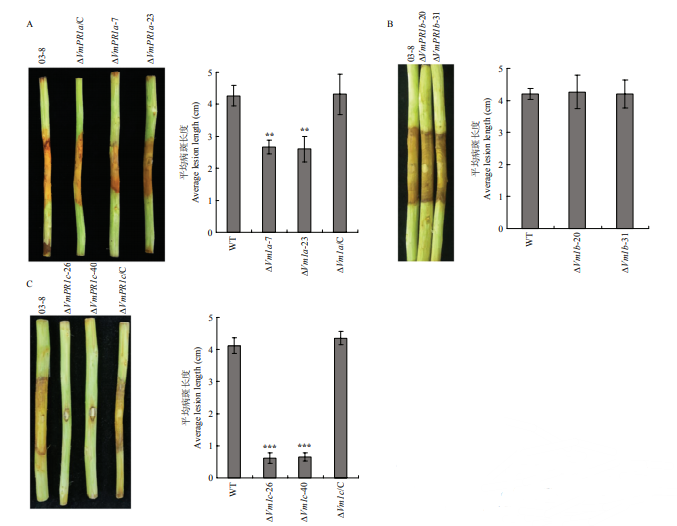 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}