Response of Liriomyza trifolii to Cold Acclimation and Differences of Cold Tolerance Among Different Populations
ZHANG QiKai,1, XING ZhenLong2, WU ShengYong1, XU RuiRui1, LEI ZhongRen,11State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing 100193 2State Key Laboratory of Crop Stress Adaptation and Improvement, School of Life Sciences, Henan University, Kaifeng 475004, Henan
Abstract 【Objective】The response of the major invasive pest Liriomyza trifolii to cold acclimation and the cold tolerance of three geographical populations (Wuhan City, Hubei Province; Haikou City, Hainan Province; Anqing City, Anhui Province) were studied to provide references for the potential colonization of L. trifolii in high latitudes of China.【Method】The pupae and adults of L. trifolii population in Hainan were exposed to low temperatures of -12, -14, -16, -18, -20, -22 and -24℃ for 30 min, and the temperature with a survival rate of 15%-30% was selected as the recognition temperature. The L. trifolii pupae and adults were put at 5℃ for 1-6 h or 12℃ for 1-6 d, and then put at the recognition temperature for 30 min to detect the survival rate. The undomesticated pupae and adults were also placed at the recognition temperature for 30 min as a control, and the difference in low-temperature survival rate after cold acclimation was compared. Besides, five target temperatures were set at 0, -5, -10, -15, and -20℃, respectively, and the pupae of three geographical populations were exposed to the target temperatures for 2 h, the difference of survival rate was studied. Finally, the supercooling point (SCP) of pupae of three geographical populations was measured by supercooling point tester.【Result】When theL. trifolii pupae and adults were exposed to -20℃ for 30 min, the survival rates were 15.0%, 19.6%, and 21.0%, respectively. Therefore, -20℃ was determined as the cold acclimation recognition temperature of L. trifolii. The cold tolerance ofL. trifolii pupae and adults was improved after rapid cold acclimation at 5℃, and the adults responded more positively to rapid cold acclimation. The effect of cold acclimation for 1 h and 2 h was the best, and the effect of rapid cold acclimation decreased gradually until disappeared with the extension of exposure time. Within 6 days, the cold tolerance ofL. trifolii pupae and adults was improved by low-temperature exposure at 12℃ for different periods, and there was little difference in cold tolerance after cold acclimation for different periods. Besides, under the low-temperature exposure of 5, 0 and -5℃, there was no significant difference in the survival rate of pupae between the Hainan population and Anhui, Hubei populations, but under the low-temperature exposure of -10, -15 and -20℃, the pupa survival rate of Hainan population was significantly lower than that of Anhui and Hubei populations. The SCP of pupae in the Anhui and Hubei populations was significantly lower than that in the Hainan population.【Conclusion】The cold tolerance of L. trifolii can be enhanced by cold acclimation, which may be one of the reasons why L. trifolii gradually spread to high latitudes in China. The cold tolerance of the Hubei and Anhui populations is stronger than that of the Hainan population. The results are helpful to predict the overwintering distribution area of L. trifolii in China and guide its monitoring, early warning, and prevention and control. Keywords:Liriomyza trifolii;cold acclimation;supercooling point;cold tolerance
PDF (423KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 张起恺, 邢振龙, 吴圣勇, 徐瑞瑞, 雷仲仁. 三叶斑潜蝇对冷驯化的响应及不同种群耐寒性差异[J]. 中国农业科学, 2021, 54(13): 2781-2788 doi:10.3864/j.issn.0578-1752.2021.13.008 ZHANG QiKai, XING ZhenLong, WU ShengYong, XU RuiRui, LEI ZhongRen. Response of Liriomyza trifolii to Cold Acclimation and Differences of Cold Tolerance Among Different Populations [J]. Scientia Acricultura Sinica, 2021, 54(13): 2781-2788 doi:10.3864/j.issn.0578-1752.2021.13.008
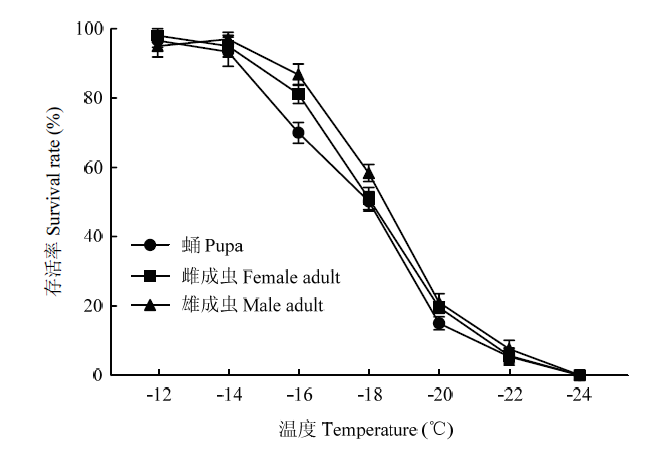
柱上不同小写字母表示差异显著(单因素方差分析,Tukey’s HSD 多重比较,P<0.05)。图3、图5同 Fig. 2Effects of rapid cold acclimation on the low-temperature survival rate of the Hainan L. trifolii at different insect stages
Different lowercases on the bars indicate significant differences (one-way ANOVA, Tukey’s HSD,P<0.05). The same asFig. 3, Fig. 5
柱上不同小写字母表示差异显著(双因素方差分析,Tukey’s HSD 多重比较,P<0.05) Fig. 4The low-temperature survival rate of L. trifolii pupae of different geographical populations under different temperatures
Different lowercases on the bars indicate significant differences (two-way ANOVA, Tukey’s HSD,P<0.05)
LEI ZR, YAO JM, ZHU CJ, WANG HH. Prediction of suitable areas for Liriomyza trifolii (Burgess) in China Plant Protection, 2007, 33(5):100-103. (in Chinese) [本文引用: 2]
JOHNSON MW, WELTER SC, TOSCANO NC, TING IP, TRUMBLE JT. Reduction of tomato leaflet photosynthesis rates by mining activity ofLiriomyza sativae(Diptera: Agromyzidae) Journal of Economic Entomology, 1983, 76(5):1061-1063. DOI:10.1093/jee/76.5.1061URL [本文引用: 1]
PARRELLA MP, JONES VP, YOUNGMAN RR, LEBECK LM. Effect of leaf mining and leaf stippling ofLiriomyzaspp.on photosynthetic rates ofChrysanthemum Annals of the Entomological Society of America, 1985, 78(1):90-93. DOI:10.1093/aesa/78.1.90URL [本文引用: 1]
REITZ SR, TRUMBLE JT. Interspecific and intraspecific differences in twoLiriomyza leafminer species in California Entomologia Experimentalis et Applicata, 2002, 102(2):101-113. DOI:10.1046/j.1570-7458.2002.00930.xURL [本文引用: 1]
WANG XH, QI XL, KANGL. The phenomenon of rapid cold acclimation of insects and its significance of ecological adaptation Progress in Natural Science, 2003, 13(11):1128-1133. (in Chinese) [本文引用: 1]
OVERGAARDJ, MALMENDALA, SØRENSENJ G, BUNDYJ G, LOESCHCKEV, NIELSENN C, HOLMSTRUPM. Metabolomic profiling of rapid cold hardening and cold shock in Drosophila melanogaster Journal of Insect Physiology, 2007, 53(12):1218-1232. DOI:10.1016/j.jinsphys.2007.06.012URL [本文引用: 1]
WANG XH, KANGL. Rapid cold hardening in young hoppers of the migratory locustLocusta migratoriaL. (Orthoptera: Acridiidae) Cryo Letters, 2003, 24(5):331-340. [本文引用: 1]
SINCLAIR BJ, CHOWN SL. Rapid cold-hardening in a Karoo beetle,Afrinussp Physiological Entomology, 2006, 31(1):98-101. DOI:10.1111/pen.2006.31.issue-1URL [本文引用: 1]
KANGL, CHENB, WEI JN, LIU TX. Roles of thermal adaptation and chemical ecology in Liriomyza distribution and control Annual Review of Entomology, 2009, 54:127-145. DOI:10.1146/annurev.ento.54.110807.090507URL [本文引用: 1]
SHINTANIY, ISHIKAWAY. Relationship between rapid cold- hardening and cold acclimation in the eggs of the yellow-spotted longicorn beetle, Psacothea hilaris. Journal of Insect Physiology, 2007, 53(10):1055-1062. DOI:10.1016/j.jinsphys.2007.05.012URL [本文引用: 2]
KONGL, GUO JY, ZHOU ZS, WAN FH. Progress in research on cold hardening in insects Chinese Journal of Applied Entomology, 2012, 49(6):1664-1669. (in Chinese) [本文引用: 2]
TURNOCK WJ, BOIVING, RING RA. Interpopulation differences in the coldhardiness of Delia radicum (Diptera: Anthomyiidae) The Canadian Entomologist, 1998, 130(2):119-129. DOI:10.4039/Ent130119-2URL [本文引用: 1]
SHINTANIY, ISHIKAWAY. Geographic variation in cold hardiness of eggs and neonate larvae of the yellow-spotted longicorn beetle Psacothea hilaris Physiological Entomology, 1999, 24(2):158-164. DOI:10.1046/j.1365-3032.1999.00126.xURL [本文引用: 1]
TANAKAK. Cold tolerance in the house spider,Achaearanea tepidariorum (Araneae: Theridiidae) Entomological Science, 1999, 2(4):597-604. [本文引用: 1]
JING XH, KANGL. Geographical variation in egg cold hardiness: A study on the adaptation strategies of the migratory locust Locusta migratoriaL Ecological Entomology, 2003, 28(2):151-158. DOI:10.1046/j.1365-2311.2003.00497.xURL [本文引用: 2]
OVERGAARDJ, HOFFMANN AA, KRISTENSEN TN. Assessing population and environmental effects on thermal resistance in Drosophila melanogaster using ecologically relevant assays Journal of Thermal Biology, 2011, 36(7):409-416. DOI:10.1016/j.jtherbio.2011.07.005URL [本文引用: 1]
CHENB, KANGL. Variation in cold hardiness of Liriomyza huidobrensis (Diptera: Agromyzidae) along latitudinal gradients Environmental Entomology, 2004, 33(2):155-164. DOI:10.1603/0046-225X-33.2.155URL [本文引用: 2]
CHENB, KANGL. Can greenhouses eliminate the development of cold resistance of the leafminers? Oecologia, 2005, 144(2):187-195. DOI:10.1007/s00442-005-0051-2URL [本文引用: 4]
XIANG JC, LEI ZR, WANG HH, GAO YL. Interspecific competition among three invasiveLiriomyzaspecies Acta Ecologica Sinica, 2012, 32(5):1616-1622. (in Chinese) [本文引用: 1]
XING ZL, LIU YQ, CAI WZ, HUANG XZ, WU SY, LEI ZR. Efficiency of trichome-based plant defense in Phaseolus vulgaris depends on insect behavior, plant ontogeny, and structure Frontiers in Plant Science, 2017, 8:2006. DOI:10.3389/fpls.2017.02006URL [本文引用: 1]
ZHAO YX, KANGL. Cold tolerance of the leafminerLiriomyza sativae (Dipt., Agromyzidae) Journal of Applied Entomology, 2000, 124(3/4):185-189. DOI:10.1046/j.1439-0418.2000.00463.xURL [本文引用: 1]
WANG HH, LEI ZR, LIX, OETTING RD. Rapid cold hardening and expression of heat shock protein genes in the B-biotypeBemisia tabaci Environmental Entomology, 2011, 40(1):132-139. DOI:10.1603/EN09357URL [本文引用: 2]
ZHANGZ, ZHENGQ, ZHANG YH, LIUJ, YIN XT, TANG QB, LIJ, YUANY, LI XR, ZHUX. Cold hardiness of laboratory populations of Spodoptera frugiperda. Plant Protection, 2019, 45(6):43-49. (in Chinese) [本文引用: 1]
CHEN CP, DENLINGER DL, LEE JRR E. Cold-shock injury and rapid cold hardening in the flesh fly Sarcophaga crassipalpis Physiological Zoology, 1987, 60(3):297-304. DOI:10.1086/physzool.60.3.30162282URL [本文引用: 1]
DANKS HV. The wider integration of studies on insect cold- hardiness European Journal of Entomology, 1996, 93:383-404. [本文引用: 1]
RAKOL, HOFFMANN AA. Complexity of the cold acclimation response in Drosophila melanogaster Journal of Insect Physiology, 2006, 52(1):94-104. DOI:10.1016/j.jinsphys.2005.09.007URL [本文引用: 1]
JENSEND, OVERGAARDJ, SØRENSENJ G. The influence of developmental stage on cold shock resistance and ability to cold-harden in Drosophila melanogaster Journal of Insect Physiology, 2007, 53(2):179-186. DOI:10.1016/j.jinsphys.2006.11.008URL [本文引用: 1]
WANG XH, KANGL. Differences in egg thermotolerance between tropical and temperate populations of the migratory locust Locusta migratoria (Orthoptera: Acridiidae) Journal of Insect Physiology, 2005, 51(11):1277-1285. DOI:10.1016/j.jinsphys.2005.07.010URL [本文引用: 1]
XING ZL. Range expansion of Liriomyza trifolii (Burgess) and competitive displacement with its’ congener species,L. sativae Blanchard, in China [D]. Beijing: China Agricultural University, 2018. (in Chinese) [本文引用: 2]
CHENB, KANGL. Implication of pupal cold tolerance for the northern over-wintering range limit of the leafminer Liriomyza sativae (Diptera: Agromyzidae) in China Applied Entomology and Zoology, 2005, 40(3):437-446. DOI:10.1303/aez.2005.437URL [本文引用: 1]
MASAKIS. Geographical variation of life cycle in crickets (Ensifera: Grylloidea) European Journal of Entomology, 1996, 93:281-302. [本文引用: 1]
KIMURA MT. Cold and heat tolerance of drosophilid flies with reference to their latitudinal distributions Oecologia, 2004, 140(3):442-449. DOI:10.1007/s00442-004-1605-4URL [本文引用: 1]
MORGAN D JW, REITZ SR, ATKINSON PW, TRUMBLE JT. The resolution of Californian populations of Liriomyza huidobrensis and Liriomyza trifolii (Diptera: Agromyzidae) using PCR Heredity, 2000, 85(1):53-61. DOI:10.1046/j.1365-2540.2000.00731.xURL [本文引用: 1]
FULLER SJ, CHAVIGNYP, LAPCHINL, VANLERBERGHE- MASUTTIF. Variation in clonal diversity in glasshouse infestations of the aphid,Aphis gossypiiGlover in southern France Molecular Ecology, 1999, 8(11):1867-1877. DOI:10.1046/j.1365-294x.1999.00782.xURL [本文引用: 1]
 ,1, 邢振龙2, 吴圣勇1, 徐瑞瑞1, 雷仲仁
,1, 邢振龙2, 吴圣勇1, 徐瑞瑞1, 雷仲仁

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT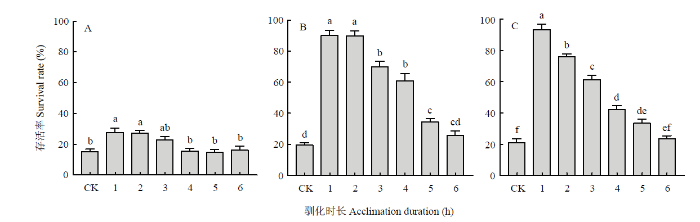 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT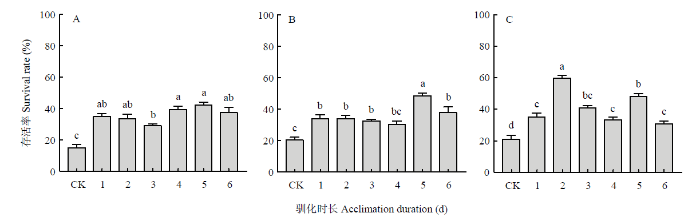 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT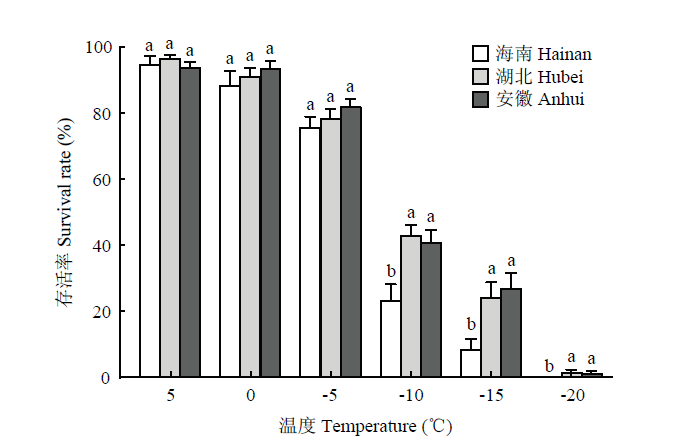 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT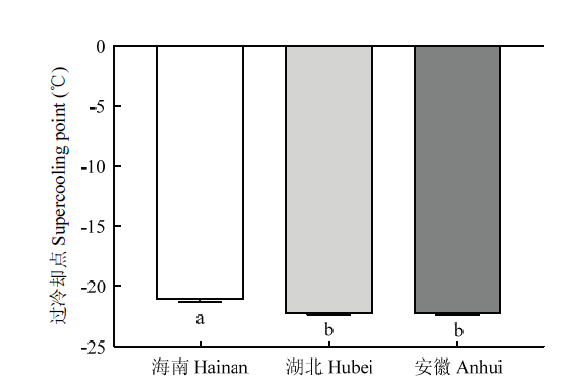 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}