 ,1,2,3, 张春梅1, 吴迪1, 陆成彬1, 董亚超3, 程晓明1, 张勇,1, 高德荣1,2
,1,2,3, 张春梅1, 吴迪1, 陆成彬1, 董亚超3, 程晓明1, 张勇,1, 高德荣1,2Screening for Resistance to Fusarium Head Blight and Agronomic Traits of Wheat Germplasms from Yangtze River Region
HU WenJing,1,2,3, ZHANG ChunMei1, WU Di1, LU ChengBin1, DONG YaChao3, CHENG XiaoMing1, ZHANG Yong,1, GAO DeRong1,2通讯作者:
责任编辑: 李莉
收稿日期:2020-01-6接受日期:2020-04-20网络出版日期:2020-11-01
| 基金资助: |
Received:2020-01-6Accepted:2020-04-20Online:2020-11-01
作者简介 About authors
胡文静,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (457KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
胡文静, 张春梅, 吴迪, 陆成彬, 董亚超, 程晓明, 张勇, 高德荣. 长江中下游小麦抗赤霉病品种的筛选与部分农艺性状分析[J]. 中国农业科学, 2020, 53(21): 4313-4321 doi:10.3864/j.issn.0578-1752.2020.21.001
HU WenJing, ZHANG ChunMei, WU Di, LU ChengBin, DONG YaChao, CHENG XiaoMing, ZHANG Yong, GAO DeRong.
0 引言
【研究意义】小麦是世界上最重要的粮食作物之一,是全世界1/3以上人口的主食[1]。中国是小麦第一大生产国、消费国和贸易国,小麦生产状况对全国社会经济发展、人民生活水平提高、国家粮食安全和社会稳定具有极其重要的影响。目前,中国乃至世界小麦生产面临的主要挑战是各种病害、虫害和非生物胁迫等威胁,其中,小麦赤霉病危害尤为严峻。小麦赤霉病(Fusarium head blight,FHB)主要由禾谷镰刀菌(Fusarium graminearum)等引起,于小麦开花期侵染穗部小花,在小麦籽粒灌浆成熟过程中不断繁殖,产生和积累各种毒素,例如脱氧雪腐镰孢菌烯醇(deoxynivalenol,DON)、雪腐镰孢菌烯醇(nivalenol,NIV)和玉米赤霉烯酮(zearalenol,ZEN),严重影响小麦产量和品质,并对人、畜健康造成巨大伤害,成为粮食安全的主要威胁[1]。黄淮冬麦区和长江中下游冬麦区是中国两大小麦主产区,年种植面积约占全国小麦总面积70%,后者是小麦赤霉病高发区和常发区。据报道,2012—2015年江苏省年均赤霉病发生面积约120万hm2[2]。近年来,随着气候变暖和玉米秸秆还田量的增加,小麦赤霉病正迅速扩展到黄淮麦区南部。据统计,近10年河南省小麦赤霉病年均发生面积约110万hm2,其中2012年达333万hm2[3]。目前,培育和推广种植抗赤霉病品种是小麦生产上有效防控赤霉病的重要途径之一[3],而从遗传背景丰富的地方品种和农艺性状优良的栽培品种中筛选出稳定抗赤霉病种质资源,是提高小麦抗赤霉病育种效率的基础。广泛搜集并鉴定筛选综合农艺性状优良、丰产性好的抗赤霉病新种质资源,拓展抗源的遗传基础,对于小麦抗赤霉病育种和生产具有重要意义。【前人研究进展】国内外科学家一直致力于开展小麦抗赤霉病种质资源的鉴定发掘[4,5,6]。中国的小麦赤霉病抗源主要有两类:一是现在应用较广泛的早期改良品种和地方品种,如中国的苏麦3号、荆州1号、武汉1号、望水白、白三月黄、海盐种[6]和它们的衍生系、巴西的Frontana、Maringa等以及欧洲的Funo等[7];二是从小麦近缘种属中发掘的抗赤霉病新种质,如大赖草属、偃麦草属、鹅观草属、山羊草属中发现的抗赤霉病种质[8,9,10,11,12,13,14]。上述两类抗源的农艺性状欠佳,如苏麦3号、望水白[15,16,17],虽利用它们选育出一批抗性好的品种(宁7840、鄂恩1号[18]等),但因丰产性差未被大面积推广应用。【本研究切入点】长江中下游麦区有很多农艺性状较好的品种对赤霉病抗性表现中抗(moderately resistance,MR)和中感(moderately susceptible,MS),如阿夫、台湾小麦和南大2419等,实践证明,育种中可以选择抗性来源不同的抗源进行聚合杂交,实现抗性基因累加[4]。本研究供试小麦品种来自长江中下游麦区,其中改良品种是通过江苏省、安徽省、湖北省等地审定或者通过国家审定的主栽品种。近年来,绝大多数通过江苏省淮南麦区审定的小麦品种的亲本中包含扬麦158或宁麦9号,说明农艺性状优良的抗赤霉病品种更易被广泛应用于育种实践。之前很多小麦抗赤霉病种质资源筛选研究往往仅注重抗性鉴定,忽视与之关联的农艺性状的分析,不能给育种家提供全面的选择信息,无法解决当前生产上抗赤霉病与产量协调提高的问题。【拟解决的关键问题】本研究选取历年来长江中下游大面积种植的小麦品种,利用国内外公认的抗赤霉病主效基因Fhb1进行分子检测,通过单花滴注接种禾谷镰刀菌鉴定筛选稳定的抗赤霉病品种,调查其株高、小穗数、穗长、穗粒数等性状,筛选农艺性状优良的抗赤霉病品种作为小麦抗赤霉病育种的亲本使用,为小麦抗赤霉病育种提供参考。1 材料与方法
1.1 试验材料
供试材料来源于江苏省、湖北省、安徽省等地的93个小麦品种,其中,改良品种75个、地方品种18个,抗赤霉病对照品种为苏麦3号,感赤霉病对照品种为扬麦13。所有材料的来源及系谱见电子附表1和电子附表2。Supplemental table 1
附表1
附表1供试材料中小麦地方品种的来源
Supplemental table 1
| 序号 | 品种名称 | 来源地 |
|---|---|---|
| No. | Landrace name | District |
| 1 | 白慈麦 Baicimai | 无锡 Wuxi |
| 2 | 白火麦 Baihuomai | 不详 Unknown |
| 3 | 白蒲 Baipu | 金华 Jinhua |
| 4 | 和蒲头 Heputou | 南陵 Nanling |
| 5 | 和尚头 Heshangtou | 兰州 Lanzhou |
| 6 | 红和尚头 Hongheshangtou | 镇江 Zhenjiang |
| 7 | 红卷芒 Hongjuanmang | 南阳 Nanyang |
| 8 | 红壳酱 Hongkejiang | 剑河 Jianhe |
| 9 | 红袖子 Hongxiuzi | 淅川 Xichuan |
| 10 | 胡须麦 Huxumai | 商县 Shangxian |
| 11 | 鲫鱼麦 Jiyumai | 东阳 Dongyang |
| 12 | 江东门 Jiangdongmen | 南京 Nanjing |
| 13 | 六柱头 Liuzhutou | 上海 Shanghai |
| 14 | 糯麦 Nuomai | 不详 Unknown |
| 15 | 四方麦 Sifangmai | 丰城 Fengcheng |
| 16 | 望水白 Wangshuibai | 溧阳 Liyang |
| 17 | 蚰子头 Youzitou | 汝南 Runan |
| 18 | 鱼鳅麦 Yuqiumai | 思南 Sinan |
新窗口打开|下载CSV
Supplemental table 2
附表2
附表2供试材料中国内改良品种的系谱和审定年份
Supplemental table 2
| 序号 | 品种名称 | 系谱 | 审定年份 |
|---|---|---|---|
| No. | Cultivars name | Pedigree | Released year |
| 1 | 扬麦1号 Yangmai 1 | 阿夫 Funo | 1967 |
| 2 | 苏麦3号 Suami 3 | 阿夫/台湾小麦 Funo/Taiwan | 1970 |
| 3 | 宁麦3号 Ningmai 3 | 郑引1号/扬麦1号 (St1472/506)/Yangmai 1 | 1973 |
| 4 | 扬麦2号 Yangmai 2 | 阿夫 Funo | 1976 |
| 5 | 扬麦3号 Yangmai 3 | 阿夫 Funo | 1976 |
| 6 | 扬麦4号 Yangmai 4 | 南大2419/胜利麦//阿夫 Nada2419/Triumph//Funo | 1980 |
| 7 | 鄂麦9号 Emai 9 | 鄂麦6号 Emai 6 | 1982 |
| 8 | 鄂麦6号 Emai 6 | 南大2419 Nanda2419 | 1983 |
| 9 | 鄂恩1号 Een 1 | (洛夫林10/761)/苏麦3号 (Lovrin 10/761)/Sumai 3 | 1985 |
| 10 | 扬麦5号 Yangmai 5 | 扬9-16/郑引1号 Yang9-16/(St1472/506) | 1986 |
| 11 | 扬麦6号 Yangmai 6 | 大丰1087/早熟5号 Dafeng1087/Zaoshu 5 | 1991 |
| 12 | 鄂麦11 Emai 11 | (760569//襄麦8号)//309 (760569//Xiangmai 8)//309 | 1993 |
| 13 | 镇麦1号 Zhenmai 1 | 镇麦7853/苏肯1号 Zhenmai 7853/Suken 1 | 1993 |
| 14 | 宁麦8号 Ningmai 8 | 扬麦5号/扬麦6号 Yangmai 5/Yangmai 6 | 1996 |
| 15 | 扬麦9号 Yangmai 9 | 鉴三/扬麦5号 Jian 3/Yangmai 5 | 1996 |
| 16 | 鄂麦12 Emai 12 | 750025-12/鄂麦6号 750025-12/Emai 6 | 1997 |
| 17 | 宁麦9号 Ningmai 9 | 扬麦6号/西风 Yangmai 6/Xifeng | 1997 |
| 18 | 苏麦5号 Sumai 5 | 西风/扬麦4号 Xifeng/Yangmai 4 | 1997 |
| 19 | 扬麦158 Yangmai 158 | 扬麦4号/郑引1号 Yangmai 4/(St1472/506) | 1997 |
| 20 | 扬麦10号 Yangmai 10 | 扬1582///(Yuma/Chancellor8)/扬麦5号//扬85-854 Yangmai 1582///(Yuma/Chancellor8)/Yangmai 5//Yang 85-854 | 1998 |
| 21 | 苏麦6号 Sumai 6 | 6698/扬麦5号 6698/Yangmai 5 | 1999 |
| 22 | 鄂麦15 Emai 15 | 882-852//鄂恩1号/NPPP-2///贵农11 882-852//EEN 1/NPPP-2///Guinong 11 | 2000 |
| 23 | 镇麦3号 Zhenmai 3 | K258 | 2000 |
| 24 | 扬麦11 Yangmai 11 | 扬1583///(Yuma/Chancellor8)/鉴二//扬85-854 Yangmai 1583///(Yuma/Chancellor8)Jian 2//Yang 85-854 | 2001 |
| 25 | 扬麦12 Yangmai 12 | 扬1583///TP114/扬麦5号//扬85-854 Yangmai 1583///TP114/Yangmai 5//Yang 85-854 | 2001 |
| 26 | 鄂麦16 Emai16 | 7023 | 2002 |
| 27 | 鄂麦17 Emai 17 | 鄂麦12 Emai 12 | 2002 |
| 28 | 鄂麦18 Emai 18 | SKUA/865146//鄂麦11 SKUA/865146//Emai 11 | 2002 |
| 29 | 鄂麦19 Emai 19 | 荆州12 Jingzhou 12 | 2002 |
| 30 | 生选3号 Shengxuan 3 | 扬麦158 Yangmai 158 | 2002 |
| 31 | 扬麦13 Yangmai 13 | 扬88-84//Maris Dove/扬麦3号 Yang88-84//Maris Dove/Yangmai 3 | 2002 |
| 32 | 扬辐麦2号 Yangfumai 2 | 扬麦158/1-9012 Yangmai 158/1-9012 | 2003 |
| 33 | 镇麦4号 Zhenmai 4 | 扬麦158/镇麦1号 Yangmai 158/Zhenmai 1 | 2003 |
| 34 | 宁麦11 Ningmai 11 | 宁麦8号 Ningmai 8 | 2004 |
| 35 | 宁麦12 Ningmai 12 | 宁9170/扬麦158 Ning 9170/Yangmai 158 | 2004 |
| 36 | 扬麦14 Yangmai 14 | 扬麦158/扬麦6号 Yangmai 158/Yangmai 6 | 2004 |
| 37 | 扬麦16 Yangmai 16 | 扬91F138/扬90-30 Yang91F138/Yang90-30 | 2004 |
| 38 | 镇麦5号 Zhenmai 5 | 扬麦158/宁麦9号 Yangmai 158/Ningmai 9 | 2004 |
| 39 | 扬麦15 Yangmai 15 | 扬麦4号/鉴三//川育21526 Yangmai 4/Jian 3//Chuanyu 21526 | 2005 |
| 40 | 扬麦17 Yangmai 17 | 扬92F101/川育21526 Yang 92F101/Chuanyu21526 | 2005 |
| 41 | 镇麦6号 Zhenmai 6 | 扬麦158/镇麦1号 Yangmai 158/Zhenmai 1 | 2005 |
| 42 | 宁麦13 Ningmai 13 | 宁麦9号 Ningmai 9 | 2006 |
| 43 | 宁麦14 Ningmai 24 | 宁麦9号 Ningmai 9 | 2006 |
| 44 | 镇麦168 Zhenmai 168 | 苏麦6号/扬97G59 Sumai 6/Yang 97G59 | 2007 |
| 45 | 宁麦15 Ningmai 15 | 宁9144 Ning 9144 | 2008 |
| 46 | 宁糯麦1号 Ningnuomai 1 | EH-5/扬麦158 EH-5/Yangmai 158 | 2008 |
| 47 | 扬辐麦4号 Yangfumai 4 | 宁麦8号/宁麦9号 Ningmai 8/Ningmai 9 | 2008 |
| 48 | 扬麦18 Yangmai18 | 宁94///扬麦1586//扬88-128/P045 Ningmai 94///Yangmai 1586//Yang 88-128/P045 | 2008 |
| 49 | 扬麦19 Yangmai 19 | 扬麦9号6////扬1583///Yuma/Chancellor8/扬麦5号//扬85-854 Yangmai 96////Yangmai 1583///Yuma/Chancellor8/Yangmai 5//Yang85-854 | 2008 |
| 50 | 宁麦16 Ningmai 16 | 宁麦8号/宁麦9号 Ningmai 8/Ningmai 9 | 2009 |
| 51 | 扬麦20 Yangmai 20 | 扬麦10号/扬麦9号 Yangmai 10/Yangmai 9 | 2010 |
| 52 | 扬糯麦1号 Yangnuomai 1 | 扬麦15///扬麦9号//扬麦9号/CAW Yangmai 15///Yangmai 9//Yangmai 9/CAW | 2010 |
| 53 | 镇麦9号 Zhenmai 9 | 苏麦6号/扬97G59 Sumai 6/Yang 97G59 | 2010 |
| 54 | 宁麦18 Ningmai 18 | 宁93123/扬93-111 Ning 93123/Yang93-111 | 2012 |
| 55 | 宁麦19 Ningmai 19 | 宁9144 Ning 9144 | 2012 |
| 56 | 扬麦22 Yangmai 22 | 扬麦9号3/宁97033-2 Yangmai 93/Ning 97033 | 2012 |
| 57 | 扬麦21 Yangmai 21 | 宁麦9号6/红卷芒 Ningmai 96/Hongjuanmang | 2013 |
| 58 | 扬麦23 Yangmai 23 | 扬麦16/扬辐麦93-11 Yangmai 16/Yangfumai 93-11 | 2013 |
| 59 | 宁麦21 Ningmai 21 | 宁9312/扬麦158//宁9312 Ningmai 9312/Yangmai 158//Ningmai 9312 | 2013 |
| 60 | 宁麦22 Ningmai 22 | 宁麦12 Ningmai 12 | 2014 |
| 61 | 宁麦24 Ningmai 24 | 宁麦9号 Ningmai 9 | 2015 |
| 62 | 扬麦24 Yangmai 24 | 扬麦172//扬11/豫麦18 Yangmai 172//Yangmai 11/Yumai 18 | 2015 |
| 63 | 宁麦26 Ningmai 26 | 宁9531/宁麦9号 Ning9531/Ningmai 9 | 2016 |
| 64 | 华麦8号 Humai 8 | 扬麦158/小偃8788 Yangmai 158/Xiaoyan 8788 | 2016 |
| 65 | 扬麦25 Yangmai 25 | 扬麦172//扬11/豫麦18 Yangmai 172//Yangmai 11/Yumai 18 | 2016 |
| 66 | 扬麦27 Yangmai 27 | 扬麦19/扬07纹5418 Yangmai 19/Yang07wen5418 | 2017 |
| 67 | 扬麦28 Yangmai 28 | 扬麦16/扬麦18//扬麦16 Yangmai 16/Yangmai 18//Yangmai 16 | 2018 |
| 68 | 扬麦29 Yangmai 29 | 镇麦9号//扬麦175/pm17 Zhenmai 9//Yangmai175/pm17 | 2019 |
| 69 | 安徽11 Anhui 11 | 弗兰尼/早洋麦 Franny/Early Premium | Unkonwn |
| 70 | 荆州66 Jingzhou 66 | 阿夫/硬粒小麦//南大2419/黑麦 Funo/Durum//Nanda 2419/Rye | Unkonwn |
| 71 | 马场2号 Machang 2 | 郑引1号 St1472/506 | Unkonwn |
| 72 | 毛颖阿夫 Maoying Funo | 阿夫 Funo | Unkonwn |
| 73 | 南大2419 Nanda2419 | Mentana | Unkonwn |
| 74 | 万年2号 Wannian 2 | 南大2419 Nanda 2419 | Unkonwn |
| 75 | 浙麦1号 Zhemai1 | 临浦早/太和小麦 Linpuzao/Taihe | Unkonwn |
新窗口打开|下载CSV
抗病性鉴定所用禾谷镰刀菌菌株(F0301、F0609、F0980和F1312)由江苏省农业科学院植物保护研究所陈怀谷研究员惠赠。
1.2 试验地点及抗病鉴定圃设置
试验于2015—2016、2016—2017、2017—2018年3个小麦生长期(以下简称2016、2017和2018)在江苏里下河地区农业科学研究所湾头试验基地赤霉病鉴定圃内进行。3年播种日期均为10月28日,每品种种植2行,行长150 cm,行距25 cm,每行均匀播种30粒,土质为沙壤土,前茬为水稻,肥力中等,田间管理如施肥、除草、灌溉及虫害防治同常规育种田。1.3 赤霉病抗性鉴定与评价
2016、2017和2018年在田间分别进行接种鉴定。参照YU等[18]方法制备赤霉菌孢子悬浮液(1×105—5×105个/mL),采用单花滴注法接种,于小麦开花初期,用注射器吸取10 μL孢子液注入麦穗(自顶部小穗开始自上而下第6个小穗的任意一个小花中),每个品种接种30个穗子。接种后采用人工弥雾保湿(每半小时喷弥雾5 min)。接种21 d后调查接种穗的病小穗数,计算病小穗率。病小穗率(the percentage of symptomatic spikelets,PSS)=发病小穗数(symptomatic spikelets)/总小穗数(total spikelets)×100%[19]。按照《中华人民共和国农业行业标准NY/T 2954-2016:小麦区域试验品种抗赤霉病鉴定技术规程》[20]标准调查和记载抗病情况,参考张晓军等[21]方法进行抗赤霉病评价。根据3年平均病小穗率和严重度分级,将供试品种分成抗病(resistant,简称R级,0<平均严重度<2.0,仅接种小穗发病,或相邻的个别小穗发病,但病斑不扩展到穗轴)、中抗(MR级,2.0≤平均严重度<3.0,穗轴发病,发病小穗占总小穗数的1/4以下)、中感(MS级,3.0≤平均严重度<3.5,穗轴发病,发病小穗占总小穗数的1/4—1/2)和感病(susceptible,简称S级,平均严重度≥3.5,穗轴发病,发病小穗占总小穗数的1/2以上)4个等级。
1.4 Fhb1的分子检测
选取每品种10粒种子,室温下发芽,取嫩叶放入液氮中速冻,采用SDS法提取基因组DNA,参考SU等[22]设计的Fhb1功能基因KASP标记引物序列,参考RASHEED等[23]检测方法,将Fhb1位点呈抗病基因型的品种简称Fhb1+、该位点呈感病基因型的品种简称Fhb1-。1.5 农艺性状调查
2018年秋,将筛选出的3年平均病小穗率≤50%的品种种植于江苏里下河地区农业科学研究所湾头试验基地产量鉴定圃,每品种种植6行,行长150 cm,行距25 cm,每行均匀播种50粒,土质为沙壤土,前茬为水稻,肥力中等,待出苗后定苗至每行40株,田间管理如施肥、除草、灌溉及虫害防治同常规育种田。2019年待各品种成熟期收获后,随机选取每品种10个单株,调查主茎株高、每穗小穗数、穗长和穗粒数(平均值),脱粒后考察千粒重(每个品种500粒称重,3次重复,取平均值,然后转化成千粒重),计算小穗着生密度(每穗小穗数/穗长)。1.6 表型统计分析
利用Microsoft Excel 2016对试验获得的病小穗数、病小穗率、农艺性状等数据进行方差分析、T检验和描述性统计分析。2 结果
2.1 小麦种质资源的赤霉病抗性鉴定与评价
3年抗赤霉病接种鉴定结果表明,地方品种只有望水白赤霉病抗性达到R级,平均病小穗率为9.7%;MR—MS级的品种有5个,平均病小穗率为15.0%—39.0%;S级的品种有12个,平均病小穗率为50.2%—78.5%,其中,红卷芒感病最重,平均病小穗率为78.5%(图1和电子附表3)。图1
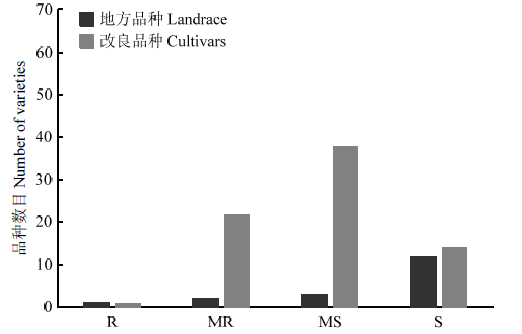 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1地方品种和改良品种中赤霉病抗性的分布
R:抗性;MR:中抗;MS:中感;S:感性
Fig. 1Distribution of resistance to FHB at four classes in wheat landrace and cultivars
R: Resistance; MR: Moderately resistance; MS: Moderately susceptible; S: Susceptible
Supplemental table 3
附表3
附表3供试小麦品种病小穗率、严重度、抗赤霉病评价和Fhb1分型结果
Supplemental table 3
| 品种名称 Name | 3年的病小穗率 PSS in three years | 平均 Average | 严重度 Severity | 抗性评价 Resistance | Fhb1 alleles | ||
|---|---|---|---|---|---|---|---|
| 2017 | 2018 | 2019 | |||||
| 望水白 Wangshuibai | 10.9 | 10.3 | 7.8 | 9.7 | 1 | R | + |
| 白慈麦 Baicimai | 15.2 | 12.5 | 17.2 | 15.0 | 2 | MR | + |
| 红壳酱 Hongkejiang | 18.9 | 16.0 | 15.2 | 16.7 | 2 | MR | - |
| 红和尚头 Hongheshangtou | 27.4 | 27.8 | 25.9 | 27.0 | 3 | MS | - |
| 白火麦 Baihuomai | 30.2 | 38.3 | 33.0 | 33.8 | 3 | MS | - |
| 和蒲头 Heputou | 32.7 | 44.9 | 39.4 | 39.0 | 3 | MS | + |
| 白蒲 Baipu | 49.2 | 52.5 | 48.9 | 50.2 | 4 | S | + |
| 红袖子 Hongxiuzi | 45.5 | 58.0 | 48.0 | 50.5 | 4 | S | - |
| 胡须麦 Huxumai | 59.4 | 50.2 | 42.9 | 50.8 | 4 | S | + |
| 糯麦 Nuomai | 50.7 | 53.2 | 48.8 | 50.9 | 4 | S | + |
| 江东门 Jiangdongmen | 51.8 | 52.7 | 48.4 | 51.0 | 4 | S | - |
| 鲫鱼麦 Jiyumai | 51.4 | 50.8 | 53.2 | 51.8 | 4 | S | - |
| 和尚头 Heshangtou | 51.3 | 56.4 | 50.1 | 52.6 | 4 | S | - |
| 四方麦 Sifangmai | 58.1 | 47.5 | 53.7 | 53.1 | 4 | S | - |
| 六柱头 Liuzhutou | 63.7 | 67.1 | 59.8 | 63.5 | 4 | S | - |
| 鱼鳅麦 Yuqiumai | 71.0 | 69.3 | 61.6 | 67.3 | 4 | S | - |
| 蚰子头 Youzitou | 78.8 | 64.7 | 64.3 | 69.3 | 4 | S | - |
| 红卷芒 Hongjuanmang | 73.0 | 81.8 | 80.6 | 78.5 | 4 | S | - |
| 苏麦3号 Sumai 3 | 6.1 | 9.7 | 7.6 | 7.8 | 1 | R | + |
| 扬麦4号 Yangmai 4 | 13.9 | 11.7 | 14.2 | 13.2 | 2 | MR | - |
| 扬辐麦2号 Yangfumai 2 | 15.0 | 15.0 | 19.2 | 16.4 | 2 | MR | - |
| 镇麦3号 Zhenmai 3 | 17.2 | 15.2 | 17.5 | 16.6 | 2 | MR | - |
| 镇麦6号 Zhenmai 6 | 17.7 | 13.9 | 18.3 | 16.6 | 2 | MR | - |
| 扬麦11 Yangmai 11 | 18.5 | 18.3 | 16.8 | 17.9 | 2 | MR | - |
| 宁麦24 Ningmai 24 | 18.3 | 19.5 | 19.2 | 19.0 | 2 | MR | - |
| 扬麦17 Yangmai 17 | 20.7 | 21.3 | 20.3 | 20.8 | 2 | MR | - |
| 宁麦18 Ningmai 18 | 19.5 | 21.8 | 23.7 | 21.7 | 2 | MR | + |
| 扬麦18 Yangmai18 | 22.6 | 19.9 | 22.8 | 21.7 | 2 | MR | + |
| 宁麦9号 Ningmai 9 | 20.9 | 20.9 | 24.3 | 22.0 | 2 | MR | + |
| 扬麦158 Yangmai 158 | 21.4 | 20.0 | 25.6 | 22.3 | 2 | MR | - |
| 宁麦13 Ningmai 13 | 24.7 | 20.6 | 23.1 | 22.8 | 2 | MR | + |
| 宁麦19 Ningmai 19 | 21.8 | 22.0 | 25.6 | 23.1 | 2 | MR | + |
| 镇麦5号 Zhenmai 5 | 20.2 | 24.9 | 24.2 | 23.1 | 2 | MR | + |
| 镇麦4号 Zhenmai 4 | 23.2 | 20.2 | 26.1 | 23.2 | 2 | MR | - |
| 宁麦26 Ningmai 26 | 21.3 | 25.9 | 22.7 | 23.3 | 2 | MR | + |
| 扬麦21 Yangmai 21 | 23.8 | 25.4 | 21.5 | 23.5 | 2 | MR | + |
| 扬麦29 Yangmai 29 | 21.0 | 26.3 | 23.8 | 23.7 | 2 | MR | - |
| 扬麦16 Yangmai 16 | 21.4 | 28.1 | 24.5 | 24.7 | 2 | MR | - |
| 扬麦23 Yangmai 23 | 24.0 | 28.6 | 21.4 | 24.7 | 2 | MR | - |
| 苏麦6号 Sumai 6 | 29.1 | 26.7 | 18.6 | 24.8 | 2 | MR | - |
| 扬麦14 Yangmai 14 | 24.8 | 29.0 | 21.0 | 24.9 | 2 | MR | - |
| 扬麦28 Yangmai 28 | 24.1 | 28.6 | 25.4 | 26.0 | 3 | MS | + |
| 扬麦22 Yangmai 22 | 29.5 | 27.5 | 22.5 | 26.5 | 3 | MS | - |
| 宁麦14 Ningmai 24 | 25.9 | 24.5 | 29.6 | 26.7 | 3 | MS | + |
| 扬糯麦1号 Yangnuomai 1 | 27.0 | 27.7 | 25.5 | 26.7 | 3 | MS | - |
| 镇麦9号 Zhenmai 9 | 23.3 | 27.6 | 29.4 | 26.8 | 3 | MS | - |
| 镇麦1号 Zhenmai 1 | 22.9 | 30.9 | 26.8 | 26.9 | 3 | MS | - |
| 扬麦24 Yangmai 24 | 24.2 | 33.9 | 23.3 | 27.2 | 3 | MS | - |
| 苏麦5号 Sumai 5 | 28.3 | 34.1 | 23.0 | 28.5 | 3 | MS | + |
| 扬麦19 Yangmai 19 | 32.0 | 25.3 | 28.3 | 28.5 | 3 | MS | - |
| 扬麦6号 Yangmai 6 | 29.8 | 26.4 | 30.1 | 28.8 | 3 | MS | - |
| 生选3号 Shengxuan 3 | 21.6 | 32.9 | 32.6 | 29.0 | 3 | MS | - |
| 扬麦27 Yangmai 27 | 32.5 | 26.5 | 28.4 | 29.1 | 3 | MS | - |
| 扬麦20 Yangmai 20 | 33.3 | 29.7 | 25.9 | 29.6 | 3 | MS | - |
| 宁麦16 Ningmai 16 | 31.2 | 29.2 | 31.6 | 30.7 | 3 | MS | + |
| 宁麦8号 Ningmai 8 | 34.0 | 22.0 | 37.7 | 31.2 | 3 | MS | - |
| 扬麦25 Yangmai 25 | 29.9 | 30.9 | 33.2 | 31.3 | 3 | MS | - |
| 镇麦168 Zhenmai 168 | 27.3 | 28.9 | 38.3 | 31.5 | 3 | MS | - |
| 扬麦9号 Yangmai 9 | 32.5 | 38.4 | 27.6 | 32.9 | 3 | MS | - |
| 万年2号 Wannian 2 | 37.8 | 26.6 | 34.3 | 32.9 | 3 | MS | - |
| 扬麦5号 Yangmai 5 | 32.4 | 34.7 | 35.8 | 34.3 | 3 | MS | - |
| 扬麦12 Yangmai 12 | 35.6 | 37.8 | 36.5 | 36.6 | 3 | MS | - |
| 宁麦11 Ningmai 11 | 34.6 | 44.5 | 31.6 | 36.9 | 3 | MS | - |
| 扬麦10号 Yangmai 10 | 37.9 | 42.9 | 37.9 | 39.6 | 3 | MS | - |
| 鄂麦9号 Emai 9 | 39.8 | 43.3 | 36.4 | 39.8 | 3 | MS | - |
| 扬麦2号 Yangmai 2 | 40.2 | 44.1 | 38.1 | 40.8 | 3 | MS | - |
| 鄂麦11 Emai 11 | 35.3 | 49.9 | 38.9 | 41.4 | 3 | MS | - |
| 马场2号 Machang 2 | 50.0 | 35.0 | 42.5 | 42.5 | 3 | MS | - |
| 华麦8号 Humai 8 | 42.1 | 41.2 | 49.6 | 44.3 | 3 | MS | - |
| 宁麦12 Ningmai 12 | 46.4 | 46.1 | 47.6 | 46.7 | 3 | MS | - |
| 宁糯麦1号 Ningnuomai 1 | 27.2 | 32.8 | 24.0 | 28.0 | 3 | MS | - |
| 宁麦21 Ningmai 21 | 38.2 | 34.8 | 44.8 | 39.3 | 3 | MS | - |
| 宁麦22 Ningmai 22 | 40.2 | 43.2 | 43.5 | 42.3 | 3 | MS | - |
| 扬辐麦4号Yangfumai 4 | 45.6 | 41.3 | 46.0 | 44.3 | 3 | MS | + |
| 鄂麦12 Emai 12 | 46.4 | 48.3 | 47.0 | 47.2 | 3 | MS | - |
| 荆州66 Jingzhou 66 | 47.9 | 43.7 | 50.7 | 47.4 | 3 | MS | - |
| 鄂恩1号 Een 1 | 47.9 | 49.4 | 46.8 | 48.0 | 3 | MS | - |
| 宁麦15 Ningmai 15 | 49.6 | 47.7 | 50.0 | 49.1 | 3 | MS | + |
| 鄂麦18 Emai 18 | 52.3 | 51.1 | 45.2 | 49.5 | 3 | MS | - |
| 扬麦3号 Yangmai 3 | 58.0 | 52.3 | 50.3 | 53.6 | 4 | S | - |
| 鄂麦17 Emai 17 | 59.9 | 46.7 | 54.1 | 53.6 | 4 | S | - |
| 浙麦1号 Zhemai1 | 55.6 | 54.0 | 52.1 | 53.9 | 4 | S | + |
| 扬麦1号 Yangmai 1 | 48.0 | 58.9 | 54.9 | 53.9 | 4 | S | - |
| 鄂麦15 Emai 15 | 56.0 | 53.0 | 53.2 | 54.1 | 4 | S | - |
| 鄂麦6号 Emai 6 | 52.1 | 54.9 | 55.6 | 54.2 | 4 | S | - |
| 毛颖阿夫 Maoying Funo | 61.9 | 59.0 | 46.0 | 55.6 | 4 | S | - |
| 鄂麦16 Emai16 | 57.8 | 55.1 | 57.3 | 56.8 | 4 | S | - |
| 安徽11 Anhui 11 | 52.6 | 56.3 | 61.9 | 56.9 | 4 | S | - |
| 扬麦15 Yangmai 15 | 51.0 | 60.8 | 59.3 | 57.0 | 4 | S | - |
| 宁麦3号 Ningmai 3 | 55.9 | 54.5 | 61.9 | 57.5 | 4 | S | - |
| 南大2419 Nada2419 | 56.8 | 55.3 | 61.2 | 57.8 | 4 | S | - |
| 鄂麦19 Emai 19 | 59.1 | 58.9 | 58.0 | 58.7 | 4 | S | - |
| 扬麦13 Yangmai 13 | 60.8 | 63.3 | 68.8 | 64.3 | 4 | S | - |
新窗口打开|下载CSV
国内改良品种只有苏麦3号的赤霉病抗性达到R级,平均病小穗率为7.8%;MR—MS级的品种有60个,平均病小穗率变幅为13.2%—49.5%;S级的品种有14个,平均病小穗率变幅为53.6%—64.3%,其中,扬麦13感病最重,平均病小穗率为64.3%(图1和电子附表3)。
2.2 Fhb1位点分子标记检测
应用Fhb1的KASP分型标记检测93个供试品种,结果表明,地方品种望水白、白慈麦、和蒲头、白蒲、胡须麦和糯麦是Fhb1+,其他12个是Fhb1-。改良品种苏麦3号(R)、宁麦18、扬麦18、宁麦9号、宁麦13、宁麦19、镇麦5号、宁麦26、扬麦21、扬麦28、宁麦14、苏麦5号、宁麦16、扬辐麦4号、宁麦15和浙麦1号是Fhb1+,其他59个是Fhb1-。统计结果表明,Fhb1+与Fhb1-品种平均病小穗率差异显著(P<0.05)(电子附表3和图2)。总之,基因型为Fhb1+品种的赤霉病抗性显著好于Fhb1-,少数Fhb1+品种表型为感病。图2
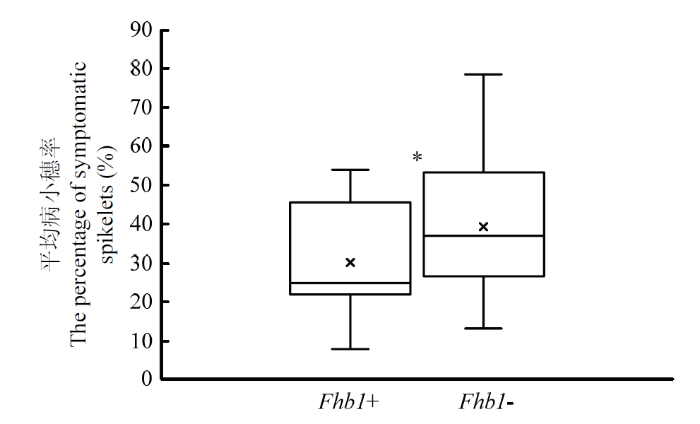 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同Fhb1基因型间的平均病小穗率
Fhb1+:Fhb1抗病等位基因;Fhb1-:Fhb1感病等位基因;*表示2种基因型间差异显著(P<0.05)
Fig. 2Comparison of the average PSS between the wheat varieties carrying different Fhb1 allele
Fhb1+: Fhb1 resistance allele: Fhb1-: Fhb1 susceptible allele; * indicates significant difference between genotypes at P<0.05
2.3 抗赤霉病品种的农艺性状分析
3年的抗赤霉病鉴定结果表明,共有67个品种的平均病小穗率≤50%,抗性达到中感及以上水平(电子附表3),其中地方品种6个,改良品种61个。2019年调查这些品种的农艺性状,利用小穗数和穗长计算小穗着生密度,结果表明,地方品种株高最大值为145.8 cm,最小值为122.6 cm,平均值为131.0 cm;穗粒数最大值为50.0粒,最小值为32.4粒,平均值为41.0粒;每穗小穗数最大为21.7个,最小为17.8个,平均值为19.1个;穗长最大值为12.7 cm,最小值为7.2 cm,平均值为10.8 cm;小穗着生密度最大值为3.0个/cm,最小值为1.4个/cm,平均值为1.9个/cm;千粒重最大值为35.5 g,最小值为26.8 g,平均值为31.5 g。改良品种株高最大值为119.6 cm,最小值为72.0 cm,平均值为86.8 cm;穗粒数最大值为50.6粒,最小值为30.9粒,平均值为43.0粒;每穗小穗数最大值为23.8个,最小值为15.7个,平均值为19.4个;穗长最大值为14.4 cm,最小值为7.9 cm,平均值为10.4 cm;小穗着生密度最大值为2.4个/cm,最小值为1.4个/cm,平均值为1.9个/cm;千粒重最大值为49.7 g,最小值为33.9 g,平均值为44.5 g(电子附表4)。Supplemental table 4
附表4
附表4平均病小穗率≤50%的小麦品种农艺性状表现
Supplemental table 4
| 品种名称 Name | 株高 Plant height(cm) | 穗粒数 Kernels per spike | 小穗数 Spikelet per spike | 穗长 Spikelet length(cm) | 穗密度(个/cm) Spikelet compactness (No./cm) | 千粒重 1000-grain weight(g) |
|---|---|---|---|---|---|---|
| 望水白 Wangshuibai | 145.8 | 33.4 | 18.7 | 12.6 | 1.5 | 35.5 |
| 白慈麦 Baicimai | 130.1 | 32.4 | 18.2 | 12.6 | 1.4 | 35.4 |
| 红壳酱 Hongkejiang | 122.6 | 43.3 | 19.8 | 11.3 | 1.8 | 26.8 |
| 红和尚头 Hongheshangtou | 123.2 | 50.0 | 21.7 | 7.2 | 3.0 | 30.7 |
| 白火麦 Baihuomai | 124.1 | 39.2 | 17.8 | 8.6 | 2.1 | 27.9 |
| 和蒲头 Heputou | 140.2 | 47.7 | 18.4 | 12.7 | 1.5 | 32.5 |
| 苏麦3号 Sumai 3 | 117.4 | 43.5 | 19.7 | 11.4 | 1.7 | 44.6 |
| 扬麦4号 Yangmai 4 | 91.8 | 43.3 | 18.6 | 8.5 | 2.2 | 49.1 |
| 扬辐麦2号 Yangfumai 2 | 86.4 | 43.1 | 20.4 | 10.7 | 1.9 | 45.9 |
| 镇麦3号 Zhenmai 3 | 86.6 | 38.7 | 18.2 | 11.8 | 1.5 | 44.6 |
| 镇麦6号 Zhenmai 6 | 86.2 | 43.1 | 18.5 | 11.0 | 1.7 | 48.1 |
| 扬麦11 Yangmai 11 | 92.6 | 39.0 | 18.3 | 10.3 | 1.8 | 49.5 |
| 宁麦24 Ningmai 24 | 72.0 | 42.5 | 17.5 | 10.1 | 1.7 | 48.0 |
| 扬麦17 Yangmai 17 | 88.6 | 42.0 | 19.0 | 10.0 | 1.9 | 45.3 |
| 宁麦18 Ningmai 18 | 81.0 | 48.1 | 17.2 | 10.1 | 1.7 | 39.8 |
| 扬麦18 Yangmai18 | 85.1 | 49.3 | 18.6 | 11.5 | 1.6 | 43.2 |
| 宁麦9号 Ningmai 9 | 78.5 | 44.0 | 18.5 | 9.6 | 1.9 | 33.9 |
| 扬麦158 Yangmai 158 | 82.4 | 35.5 | 18.3 | 9.2 | 2.0 | 47.8 |
| 宁麦13 Ningmai 13 | 80.3 | 44.0 | 17.9 | 9.6 | 1.9 | 41.1 |
| 宁麦19 Ningmai 19 | 89.0 | 45.0 | 18.5 | 9.8 | 1.9 | 46.0 |
| 镇麦5号 Zhenmai 5 | 85.5 | 35.6 | 19.0 | 7.9 | 2.4 | 39.0 |
| 镇麦4号 Zhenmai 4 | 81.6 | 43.5 | 18.0 | 10.7 | 1.7 | 46.5 |
| 宁麦26 Ningmai 26 | 88.0 | 42.7 | 17.0 | 10.1 | 1.7 | 48.5 |
| 扬麦21 Yangmai 21 | 81.9 | 43.6 | 17.1 | 10.6 | 1.6 | 44.3 |
| 扬麦29 Yangmai 29 | 78.0 | 46.3 | 20.0 | 10.5 | 1.9 | 47.3 |
| 扬麦16 Yangmai 16 | 86.3 | 45.1 | 18.8 | 9.8 | 1.9 | 49.5 |
| 扬麦23 Yangmai 23 | 78.9 | 46.2 | 19.1 | 9.8 | 1.9 | 45.4 |
| 苏麦6号 Sumai 6 | 74.8 | 46.0 | 20.9 | 10.3 | 2.0 | 40.7 |
| 扬麦14 Yangmai 14 | 81.9 | 30.9 | 19.2 | 9.9 | 1.9 | 46.9 |
| 扬麦28 Yangmai 28 | 90.8 | 37.7 | 18.8 | 11.0 | 1.7 | 47.1 |
| 扬麦22 Yangmai 22 | 79.1 | 42.7 | 19.8 | 10.2 | 1.9 | 46.3 |
| 宁麦14 Ningmai 24 | 80.2 | 41.3 | 17.9 | 9.5 | 1.9 | 45.5 |
| 扬糯麦1号 Yangnuomai 1 | 76.3 | 45.2 | 18.8 | 9.2 | 2.0 | 49.7 |
| 镇麦9号 Zhenmai 9 | 80.7 | 40.9 | 19.1 | 9.2 | 2.1 | 47.6 |
| 镇麦1号 Zhenmai 1 | 96.2 | 47.6 | 20.0 | 9.6 | 2.1 | 48.9 |
| 扬麦24 Yangmai 24 | 83.3 | 36.3 | 19.4 | 9.7 | 2.0 | 44.9 |
| 苏麦5号 Sumai 5 | 83.6 | 45.5 | 18.7 | 9.5 | 2.0 | 39.9 |
| 扬麦19 Yangmai 19 | 74.1 | 40.7 | 20.5 | 10.0 | 2.1 | 47.5 |
| 扬麦6号 Yangmai 6 | 75.4 | 47.7 | 21.3 | 10.1 | 2.1 | 44.8 |
| 生选3号 Shengxuan 3 | 101.9 | 43.1 | 21.7 | 14.4 | 1.5 | 45.3 |
| 扬麦27 Yangmai 27 | 75.2 | 40.0 | 21.3 | 11.3 | 1.9 | 43.1 |
| 扬麦20 Yangmai 20 | 89.9 | 45.5 | 19.0 | 10.7 | 1.8 | 45.5 |
| 宁麦16 Ningmai 16 | 90.0 | 47.4 | 19.5 | 9.7 | 2.0 | 46.4 |
| 宁麦8号 Ningmai 8 | 72.8 | 41.9 | 20.2 | 8.3 | 2.4 | 44.9 |
| 扬麦25 Yangmai 25 | 82.0 | 41.6 | 19.6 | 10.6 | 1.8 | 45.5 |
| 镇麦168 Zhenmai 168 | 80.6 | 41.3 | 19.3 | 9.6 | 2.0 | 47.4 |
| 扬麦9号 Yangmai 9 | 76.6 | 43.2 | 20.9 | 9.6 | 2.2 | 40.7 |
| 万年2号 Wannian 2 | 119.6 | 46.2 | 19.1 | 11.6 | 1.7 | 43.0 |
| 扬麦5号 Yangmai 5 | 92.3 | 44.6 | 21.4 | 11.8 | 1.8 | 49.1 |
| 扬麦12 Yangmai 12 | 86.4 | 40.0 | 19.0 | 9.2 | 2.1 | 46.2 |
| 宁麦11 Ningmai 11 | 82.8 | 46.9 | 23.0 | 12.0 | 1.9 | 39.3 |
| 扬麦10号 Yangmai 10 | 90.5 | 39.7 | 19.1 | 9.7 | 2.0 | 48.3 |
| 鄂麦9号 Emai 9 | 99.1 | 39.4 | 20.2 | 11.1 | 1.8 | 42.7 |
| 扬麦2号 Yangmai 2 | 106.1 | 46.7 | 19.5 | 9.5 | 2.1 | 35.7 |
| 鄂麦11 Emai 11 | 83.4 | 40.0 | 20.0 | 12.2 | 1.6 | 48.3 |
| 马场2号 Machang 2 | 79.6 | 50.6 | 19.2 | 9.2 | 2.1 | 36.8 |
| 华麦8号 Humai 8 | 75.7 | 34.7 | 18.4 | 12.4 | 1.5 | 36.9 |
| 宁麦12 Ningmai 12 | 89.3 | 45.1 | 22.6 | 14.1 | 1.6 | 47.6 |
| 宁糯麦1号 Ningnuomai 1 | 91.5 | 41.7 | 19.1 | 9.5 | 2.0 | 43.8 |
| 宁麦21 Ningmai 21 | 89.4 | 44.8 | 18.2 | 10.5 | 1.7 | 46.5 |
| 宁麦22 Ningmai 22 | 88.7 | 43.3 | 19.8 | 12.0 | 1.7 | 46.8 |
| 扬辐麦4号 Yangfumai 4 | 87.5 | 45.7 | 19.4 | 9.9 | 2.0 | 48.9 |
| 鄂麦12 Emai 12 | 85.6 | 42.8 | 20.5 | 9.5 | 2.2 | 39.9 |
| 荆州66 Jingzhou 66 | 110.3 | 40.9 | 21.3 | 12.0 | 1.8 | 38.5 |
| 鄂恩1号 Een 1 | 100.5 | 42.5 | 23.8 | 13.6 | 1.8 | 36.7 |
| 宁麦15 Ningmai 15 | 92.6 | 45.3 | 15.7 | 10.9 | 1.4 | 43.5 |
| 鄂麦18 Emai 18 | 100.9 | 39.4 | 20.5 | 11.0 | 1.9 | 42.0 |
新窗口打开|下载CSV
地方品种株高普遍偏高,千粒重普遍偏低,综合农艺性状差。改良品种的株高变辐范围较大,43个品种的株高小于90.0 cm,穗粒数达到或者超过45.0粒的有20个品种,其中宁麦19、扬麦29、扬麦16、扬麦23、扬糯麦1号、镇麦1号、扬麦20、宁麦12和扬辐麦4号的千粒重达到或者高于45.0 g(电子附表4),综合分析67个赤霉病中感及以上水平品种的主要农艺性状表现,认为宁麦19、扬麦29、扬麦16、扬麦23、扬糯麦1号、扬麦20、宁麦12和扬辐麦4号的综合农艺性状优良,穗粒数和千粒重表现突出(表1)。
Table 1
表1
表1综合农艺性状优良且赤霉病抗性达到中感赤霉病及以上水平的品种
Table 1
| 品种名称 Name | 株高 Plant height (cm) | 穗粒数 Kernels per spike | 小穗数 Spikelet per spike | 穗长 Spikelet length (cm) | 小穗着生密度(个/cm) Spikelet compactness (No./cm) | 千粒重 1000-grain weight (g) |
|---|---|---|---|---|---|---|
| 宁麦19 Ningmai 19 | 89.0 | 45.0 | 18.5 | 9.8 | 1.9 | 46.0 |
| 扬麦29 Yangmai 29 | 78.0 | 46.3 | 20.0 | 10.5 | 1.9 | 47.3 |
| 扬麦16 Yangmai 16 | 86.3 | 45.1 | 18.8 | 9.8 | 1.9 | 49.5 |
| 扬麦23 Yangmai 23 | 78.9 | 46.2 | 19.1 | 9.8 | 1.9 | 45.4 |
| 扬糯麦1号 Yangnuomai 1 | 76.3 | 45.2 | 18.8 | 9.2 | 2.0 | 49.7 |
| 扬麦20 Yangmai 20 | 89.9 | 45.5 | 19.0 | 10.7 | 1.8 | 45.5 |
| 宁麦12 Ningmai 12 | 89.3 | 45.1 | 22.6 | 14.1 | 1.6 | 47.6 |
| 扬辐麦4号 Yangfumai 4 | 87.5 | 45.7 | 19.4 | 9.9 | 2.0 | 48.9 |
新窗口打开|下载CSV
3 讨论
3.1 抗赤霉病优异种质资源筛选
小麦赤霉病抗性由多基因控制,抗赤霉病QTL除具有加性效应外,也存在上位性效应[6]。本研究经过连续3年单花滴注抗赤霉病鉴定,筛选出67个赤霉病抗性达到中感及以上水平的品种,平均病小穗率为7.8%—49.5%,其中,望水白和苏麦3号分别是地方品种和改良品种中唯一的赤霉病抗性达到R级的品种,说明小麦R级抗赤霉病的种质资源十分稀少,在抗赤霉病遗传改良中搜集此类抗源较困难。已有研究[24,25]表明品种赤霉病抗性达到中抗至中感级的抗性一般受多基因控制,抗性稳定,可被广泛利用。程顺和等[26]提出抗赤霉病育种的第2条技术路线,即选用丰产、抗赤性中感至中抗品种(系)杂交,后代注重丰产,兼顾抗赤霉病等性状选择,可育成大面积推广种植的中抗赤霉病品种。地方品种株高普遍偏高、千粒重普遍较低(电子附表4),即使赤霉病抗性很好,也不利于小麦抗赤霉病育种直接利用,而改良品种的株高、千粒重等农艺性状较地方品种优良很多,利于生产上小麦抗赤霉病育种直接利用。以往研究表明,长江中下游小麦产量育种的策略是增加粒数、提高粒重为主[27],因此,每穗粒数和千粒重的选择是决定该麦区小麦产量的关键因素。本研究表明宁麦19、扬麦29、扬麦16、扬麦23、扬糯麦1号、扬麦20、宁麦12和扬辐麦4号的综合农艺性状优良,穗粒数和千粒重表现突出,可作为抗赤霉病育种的亲本选用。3.2 不同Fhb1基因型在供试品种中的分布
已知苏麦3号是国际上研究和利用最广泛的赤霉病抗源[6],其携带的抗赤霉病主效基因Fhb1已经被克隆[28,29],有研究表明[30,31],不同来源的Fhb1抗病基因(供体),导入受体后赤霉病抗性表现差异显著,即使携带同一供体Fhb1抗病基因的材料在不同遗传背景下抗性也表现不同。朱展望等[32]通过基因检测和系谱分析发现,中国小麦改良品种所含Fhb1至少有2个来源,分别为苏麦3号和宁麦9号,并以后者为主。对本研究供试材料中改良品种系谱分析表明宁麦18、扬麦18、宁麦9号、宁麦13、宁麦19、镇麦5号、宁麦26、扬麦21、扬麦28、宁麦14、苏麦5号、宁麦16、扬辐麦4号和宁麦15等14个品种中的Fhb1抗病基因来源于宁麦9号(电子附表2)。本研究筛选出67个赤霉病抗性在中感及以上水平品种,分子标记检测表明Fhb1+品种有18个,占26.9%:其中6个地方品种中共有3个携带Fhb1抗病基因,占50%,分别是望水白、和蒲头和白慈麦;61个改良品种中共有15个携带Fhb1抗病基因,占24.2%,说明有一定数量的中感及以上水平的品种并不携有Fhb1抗病基因,在这些不携带Fhb1抗病基因的品种中有17个赤霉病抗性达到中抗,这是一些非常宝贵的抗赤霉病种质资源,可通过配制双亲群体,挖掘新的抗赤霉病基因。
抗性评价为S级的品种中白蒲、胡须麦、糯麦和浙麦1号也携有来源尚不清楚的Fhb1抗病基因,推测这些感病品种中Fhb1可能与其他基因存在互作效应,导致品种赤霉病抗性改变或者降低。
3.3 扬麦系列品种的应用价值
已有报道表明,利用抗赤霉病基因/QTL进行抗病育种时,由于基因间的连锁而导入一些不利基因,从而导致某些农艺性状变差[33,34],而LI等[35]分别以宁麦9号、宁麦13和建阳84为Fhb1抗病基因供体,以黄淮冬麦区小麦品种为背景,构建6个回交群体,分别考察这6个群体中携带Fhb1纯合抗病基因型(Fhb1-R)、纯合感病基因型(Fhb1-S)和杂合基因型(Fhb1-H)的株高、穗长、每穗小穗数和千粒重等农艺性状考察,发现没有显著的差异,这可能是育种过程中不断选择的结果,也可能跟特定的遗传背景表达有关。本研究供试品种中扬麦系列品种的平均病小穗率为13.2%—64.3%,86%的扬麦系列品种赤霉病抗性达到中抗—中感水平,其中10个赤霉病抗性达到中抗,平均病小穗率为13.2%—25.0%;15个赤霉病抗性达到中感,平均病小穗率为26.0%—40.8%;4个感赤霉病,平均病小穗率为53.6%—64.3%。考察67份赤霉病达到中感及以上水平品种的主要农艺性状,结果表明,8个品种的综合农艺性状优良,穗粒数和千粒重表现突出,可作为抗赤霉病育种的亲本选用,其中扬麦系列品种有6份,且只有扬辐麦4号含有Fhb1抗病基因。蒋正宁等[36]利用已知3个抗扩展QTL(Fhb1、Fhb2和QFhb-2DL)的7个分子标记对202份扬麦品种(系)进行分子检测,结果表明,扬麦品种(系)所含赤霉病抗扩展基因至少有2个来源,其一为含有Fhb1抗病基因的品种,以扬麦18、扬麦21和扬麦28等为代表,其抗源主要来自宁麦9号或其衍生品种;其二为不含有Fhb1抗病基因的品种,以扬麦14、扬麦16、扬麦17和扬麦23等为代表,推测这类品种所含抗性基因来自扬麦158。胡文静等[3]研究表明,扬麦16中携带抗赤霉病位点QFhb.yaas-2DL、QFhb.yaas-3BL、QFhb.yaas-4DS和QFhb.yaas-6AS,表型贡献率为8.8%—15.0%,其中QFhb.yaas-2DL与已报道的Wuhan-1中检测到的2DL上抗赤霉病位点QFhs.crc-2D位置相近,ZHU等[37]利用小麦90K SNP芯片对240份国内外品种进行抗赤霉病全基因组关联分析,在19个扬麦品种(系)里检测到QFhb.hbaas-2DL位点抗性等位变异,且证实与Wuhan-1的QFhs.crc-2D位点是一致的。JIANG等[38]利用扬麦158/宁麦9号的RIL群体定位到来源于扬麦158的抗赤霉病位点QFhb-5A和来源于宁麦9号的Fhb1,且后代中同时具有2个位点的家系赤霉病抗性显著高于亲本。HU等[39]通过全基因组关联分析在染色体4A和5D上检测到来源于扬麦衍生系苏麦6号、镇麦168和镇麦9号的抗赤霉病优异单倍型。经过系谱分析推测扬麦抗赤霉病基因来源于最原始的亲本之一阿夫(电子附表2)。下一步将精细定位扬麦中携带的抗赤霉病主效QTL,开发利于抗赤霉病育种的分子标记,评鉴它们的抗赤霉病效应和对产量等性状的影响,解析在不同遗传背景下的效应,明确其最优组合及育种应用价值;同时,继续将已知抗病基因导入长江中下游大面积主栽品种中,在保持优良农艺性状的基础上,聚合多个抗赤霉病基因,进一步提升小麦品种的赤霉病抗性水平。
4 结论
除苏麦3号和望水白外,供试材料中未发现其他高抗赤霉病的种质资源,赤霉病抗性达到中感及以上水平的品种有67个,这些品种中仅26.9%携带Fhb1抗病基因。最终筛选得到的赤霉病抗性与综合农艺性状结合较好的小麦品种以扬麦品种为主,其中只有扬辐麦4号携带Fhb1抗病基因,说明利用Fhb1抗病基因并不是解决小麦赤霉病危害的唯一途径。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 4]
[本文引用: 4]
DOI:10.1007/s00122-004-1640-xURLPMID:15057418 [本文引用: 1]

Fusarium head blight (FHB) reduces grain yield and quality in common and durum wheat. Host FHB resistance is an effective control measure that is achieved by stacking multiple resistance genes into a wheat line. Therefore, breeders would benefit from knowing which resistance sources carry different resistance genes. A diverse collection of FHB-resistant and -susceptible wheat lines was characterized with microsatellite markers linked to FHB resistance quantitative trait loci (QTLs) on chromosomes 2DL, 3BS (distal to the centromere), 3BSc (proximal to the centromere), 4B, 5AS and 6BS identified in wheat lines Maringa, Sumai 3 and Wuhan 1. Putative Sumai 3 QTLs were commonly observed in advanced breeding lines, whereas putative Maringa and Wuhan 1 QTLs were relatively rare. Marker data suggested the 3BS, 3BSc and 5AS QTLs in the Brazilian cv. Maringa were derived from Asian germplasm and not from Frontana or other Brazilian lines. Haplotype diversity was reduced near the 5AS QTL, which might impact the deployment of this QTL. Finally, Brazilian germplasm was not closely related to other resistance sources and might be useful for pyramiding with Asian wheat-derived FHB resistance.
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s10681-005-2437-yURL [本文引用: 1]
DOI:10.2135/cropsci2009.03.0115URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:28555151 [本文引用: 1]
DOI:10.1007/s00122-014-2399-3URLPMID:25239219 [本文引用: 1]
KEY MESSAGE: Recent advances in Setaria genomics appear promising for genetic improvement of cereals and biofuel crops towards providing multiple securities to the steadily increasing global population. The prominent attributes of foxtail millet (Setaria italica, cultivated) and green foxtail (S. viridis, wild) including small genome size, short life-cycle, in-breeding nature, genetic close-relatedness to several cereals, millets and bioenergy grasses, and potential abiotic stress tolerance have accentuated these two Setaria species as novel model system for studying C4 photosynthesis, stress biology and biofuel traits. Considering this, studies have been performed on structural and functional genomics of these plants to develop genetic and genomic resources, and to delineate the physiology and molecular biology of stress tolerance, for the improvement of millets, cereals and bioenergy grasses. The release of foxtail millet genome sequence has provided a new dimension to Setaria genomics, resulting in large-scale development of genetic and genomic tools, construction of informative databases, and genome-wide association and functional genomic studies. In this context, this review discusses the advancements made in Setaria genomics, which have generated a considerable knowledge that could be used for the improvement of millets, cereals and biofuel crops. Further, this review also shows the nutritional potential of foxtail millet in providing health benefits to global population and provides a preliminary information on introgressing the nutritional properties in graminaceous species through molecular breeding and transgene-based approaches.
URLPMID:25726000 [本文引用: 1]
URLPMID:27776114 [本文引用: 1]
DOI:10.1038/s41588-019-0425-8URLPMID:31182809 [本文引用: 1]
Fusarium head blight (FHB), which is mainly caused by Fusarium graminearum, is a destructive wheat disease that threatens global wheat production. Fhb1, a quantitative trait locus discovered in Chinese germplasm, provides the most stable and the largest effect on FHB resistance in wheat. Here we show that TaHRC, a gene that encodes a putative histidine-rich calcium-binding protein, is the key determinant of Fhb1-mediated resistance to FHB. We demonstrate that TaHRC encodes a nuclear protein conferring FHB susceptibility and that a deletion spanning the start codon of this gene results in FHB resistance. Identical sequences of the TaHRC-R allele in diverse accessions indicate that Fhb1 had a single origin, and phylogenetic and haplotype analyses suggest that the TaHRC-R allele most likely originated from a line carrying the Dahongpao haplotype. This discovery opens a new avenue to improve FHB resistance in wheat, and possibly in other cereal crops, by manipulating TaHRC sequence through bioengineering approaches.
DOI:10.1038/s41588-019-0426-7URLPMID:31182810 [本文引用: 1]
Head or ear blight, mainly caused by Fusarium species, can devastate almost all staple cereal crops (particularly wheat), resulting in great economic loss and imposing health threats on both human beings and livestock(1-3). However, achievement in breeding for highly resistant cultivars is still not satisfactory. Here, we isolated the major-effect wheat quantitative trait locus, Qfhs.njau-3B, which confers head blight resistance, and showed that it is the same as the previously designated Fhb1. Fhb1 results from a rare deletion involving the 3' exon of the histidine-rich calcium-binding-protein gene on chromosome 3BS. Both wheat and Arabidopsis transformed with the Fhb1 sequence showed enhanced resistance to Fusarium graminearum spread. The translation products of this gene's homologs among plants are well conserved and might be essential for plant growth and development. Fhb1 could be useful not only for curbing Fusarium head blight in grain crops but also for improving other plants vulnerable to Fusarium species.
URLPMID:16791697 [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:30136107 [本文引用: 1]
URLPMID:27306516 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:29780395 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1038/s41588-019-0425-8URLPMID:31182809 [本文引用: 1]
Fusarium head blight (FHB), which is mainly caused by Fusarium graminearum, is a destructive wheat disease that threatens global wheat production. Fhb1, a quantitative trait locus discovered in Chinese germplasm, provides the most stable and the largest effect on FHB resistance in wheat. Here we show that TaHRC, a gene that encodes a putative histidine-rich calcium-binding protein, is the key determinant of Fhb1-mediated resistance to FHB. We demonstrate that TaHRC encodes a nuclear protein conferring FHB susceptibility and that a deletion spanning the start codon of this gene results in FHB resistance. Identical sequences of the TaHRC-R allele in diverse accessions indicate that Fhb1 had a single origin, and phylogenetic and haplotype analyses suggest that the TaHRC-R allele most likely originated from a line carrying the Dahongpao haplotype. This discovery opens a new avenue to improve FHB resistance in wheat, and possibly in other cereal crops, by manipulating TaHRC sequence through bioengineering approaches.
URLPMID:31182810 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:32174947 [本文引用: 1]
[本文引用: 1]
DOI:10.1186/s12870-019-2177-0URLPMID:31959107 [本文引用: 1]
BACKGROUND: Fusarium head blight (FHB), primarily caused by Fusarium graminearum, is a major threat to wheat production and food security worldwide. Breeding stably and durably resistant cultivars is the most effective approach for managing and controlling the disease. The success of FHB resistance breeding relies on identification of an effective resistant germplasm. We conducted a genome-wide association study (GWAS) using the high-density wheat 90 K single nucleotide polymorphism (SNP) assays to better understand the genetic basis of FHB resistance in natural population and identify associated molecular markers. RESULTS: The resistance to FHB fungal spread along the rachis (Type II resistance) was evaluated on 171 wheat cultivars in the 2016-2017 (abbr. as 2017) and 2017-2018 (abbr. as 2018) growing seasons. Using Illumina Infinum iSelect 90 K SNP genotyping data, a genome-wide association study (GWAS) identified 26 loci (88 marker-trait associations), which explained 6.65-14.18% of the phenotypic variances. The associated loci distributed across all chromosomes except 2D, 6A, 6D and 7D, with those on chromosomes 1B, 4A, 5D and 7A being detected in both years. New loci for Type II resistance were found on syntenic genomic regions of chromsome 4AL (QFhb-4AL, 621.85-622.24 Mb) and chromosome 5DL (QFhb-5DL, 546.09-547.27 Mb) which showed high collinearity in gene content and order. SNP markers wsnp_JD_c4438_5568170 and wsnp_CAP11_c209_198467 of 5D, reported previously linked to a soil-borne wheat mosaic virus (SBWMV) resistance gene, were also associated with FHB resistance in this study. CONCLUSION: The syntenic FHB resistant loci and associated SNP markers identified in this study are valuable for FHB resistance breeding via marker-assisted selection.

{kind=link}
{kind=link}
{kind=link}
{kind=link}