Mixed Inheritance Analysis and QTL Mapping for γ-Tocopherol Content in Soybean
LIANG HuiZhen1,2, XU LanJie1, DONG Wei1, YU YongLiang1, YANG HongQi1, TAN ZhengWei1, LI Lei1, LIU XinMei1责任编辑: 李莉
收稿日期:2019-09-26接受日期:2019-12-3网络出版日期:2020-06-01
| 基金资助: |
Received:2019-09-26Accepted:2019-12-3Online:2020-06-01
作者简介 About authors
梁慧珍,Tel:0371-65751589;E-mail: lhzh66666@163.com

摘要
关键词:
Abstract
Keywords:
PDF (2208KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
梁慧珍, 许兰杰, 董薇, 余永亮, 杨红旗, 谭政委, 李磊, 刘新梅. 大豆γ-生育酚的混合遗传分析与QTL定位[J]. 中国农业科学, 2020, 53(11): 2149-2160 doi:10.3864/j.issn.0578-1752.2020.11.002
LIANG HuiZhen, XU LanJie, DONG Wei, YU YongLiang, YANG HongQi, TAN ZhengWei, LI Lei, LIU XinMei.
0 引言
【研究意义】维生素E(vitamin E,VE)是脂溶性维生素,参与生物体内信号转导、基因表达调控、抗低温胁迫等多种代谢途径,具有抗氧化、抗衰老、抗不育、预防早老性痴呆症和老年痴呆症、防癌、提高机体免疫力等生理活性功能[1,2,3]。维生素E分为两类:天然VE和合成VE。天然维生素E包括α-、β-、γ-和δ-4种生育酚以及α-、β-、γ-、δ-4种剩余三烯酚。合成维生素E是生育酚的醋酸酯,严格意义说并不算是真正的生育酚,生物活性与天然VE相差甚远。同时,由于合成品中含有杂质成分,也可能存在潜在的危害,使得人们更加倾向于使用天然VE[4]。目前,人类获得的VE,20%来源于天然VE,80%来源于合成VE,并且全球合成VE供应商主要集中在新和成(NHU)、帝斯曼(DSM)、巴斯夫(BASF)和浙江医药公司,市场对天然VE的需求日趋增大[5]。γ-生育酚(γ-Tocopherol,γ-T)是生育酚同族体,其基本结构为色原醇环与一个含有16个碳的侧链。γ-T主要存在于大豆种子中,在大豆种子VE总含量中占比高达66%,人体通过油脂摄入γ-生育酌的量较多[6]。γ-T是大豆油中的一种天然抗氧化剂,可用来保持油脂的风味、延长油脂的储藏时间以及保证种子长期储藏后的活力[7]。γ-T具有抗癌、防癌功效,其作用主要通过上调抑癌或下调促癌相关基因表达、干预细胞癌变相关信号传导通路、抑制肿瘤细胞增殖、促进肿瘤细胞凋亡等方式实现[8]。【前人研究进展】目前,γ-T的抗癌和防癌研究主要集中在γ-T的吸收和代谢[9,10]、γ-T抗癌作用及其分子机制[11,12,13]等方面;γ-T QTL定位研究主要集中在玉米[14]、向日葵[15]、油菜[16]、芥菜[17]和大麦[18],大豆γ-T的分子遗传和QTL定位较少。LI等[19,20]利用Bayfield × Hefeng 25大豆RIL群体检测到8个控制γ-T的QTL,分别定位在第2、4、6、8、10、15、16和18染色体上。SHAW等[21]利用Bayfield×Shire重组自交系群体(RIL),检测到8个控制γ-T的QTL,分别定位在第1、4、9、13、14、15、17和20染色体上。李海燕[22]利用Bayfield×合丰25大豆BIEX群体,检测到5个控制γ-T的QTL,分别定位在第2、7、18和19染色体上。张红梅等[23]利用Essex×ZDD2315大豆RIL群体分别检测到2个和2对控制大豆γ-T的加性和上位性互作QTL。【本研究切入点】国内外已检测到的大豆γ-T QTL数量较少,能够稳定表现的QTL更少,采取分子标记辅助选择进行高γ-T含量大豆新品种选育比较困难。【拟解决的关键问题】本研究以晋豆23×灰布支黑豆衍生的RIL为试验材料,采用主基因+多基因混合遗传分离分析方法[24],分析γ-T混合遗传规律;利用WinQTLCart2.5[25]复合区间模型分析方法,对大豆γ-T进行QTL分析,为高γ-T大豆品种选育提供参考依据。1 材料与方法
1.1 试验材料
供试材料为山西省农业科学院经济作物研究所配制的晋豆23×灰布支黑豆杂交组合。该组合包含233个家系的重组自交系(RIL)群体,即每一F2单株衍生成F2家系,每一F2家系再选株建成家系,至2011年是F14代。1.2 试验方法
2011年6月11日、2012年6月15日和2015年6月12日在河南省农业科学院现代农业科技试验基地(原阳)、2011年11月18日、2012年11月20日和2015年11月12日在海南省三亚南繁基地种植233个家系的RIL群体F14、F15和F16代材料及其亲本,每种材料均种植30粒种子。每个田间试验采取随机区组设计,2次重复,行长和行距分别为4.0和0.4 m,常规田间管理。四周分设保护行,一播全苗。2011年10月8日、2012年10月12日和2015年10月10日在河南省农业科学院现代农业科技试验基地(原阳)收获,2012年02月20日、2013年02月25日和2016年02月15日在海南省三亚南繁基地收获。收获后,6个环境下随机区组设计2次重复的种子,从每个家系选取15.00 g大豆籽粒,要求籽粒饱满,大小一致,用旋风磨(Retsch ZM100, Φ = 1.0 mm, Rheinische, Germany)磨粉。取2.5 g豆粉样品,加入30 mL CH3CH2OH混合均匀,再加入10 mL KOH溶液和5 mL浓度10%维生素C溶液,用恒温水浴皂化30 min后提取皂化液,采用100 mL C6H14分3次进行,弃去水层;最后,用Na2SO4过滤提取液,旋转蒸发瓶蒸发后用氮气吹干残留液,再用2.5 mL CH3OH溶液进行溶解,通过0.45 μm滤膜,4℃备用。利用高效液相色谱技术(high performance liquid chromatography,HPLC),采用外标法对γ-T进行定量分析。色谱柱:DIKMA;柱规格:250.0 mm×4.6 mm;柱填料:symmetry;柱温:40℃;钻石:C18,5 μm;荧光检测器激发波长:290 nm;发射波长:330 nm;流动相:CH3OH;流速:1.5 mL·min-1。进样量:20 μL;检测时间:10 min。将γ-T峰面积代入回归方程进行定量分析,计算出γ-T含量。γ-T计量单位:μg·g-1。1.3 数据分析
利用SPSS 19.0软件对γ-T进行描述统计和联合方差分析。遗传变异系数(genetic coefficient of variation,GCV)(100%)=σg/μ×100。其中,σg为遗传标准差,μ为群体平均数。遗传率h2 = σg2/[σg2+(σge2/n)+(σ2/rn)],其中,σg2为遗传方差;σge2为基因型×环境方差,n为年份数,σ2为误差方差,r为重复数。采用主基因+多基因混合遗传分离分析法[24]对晋豆23×灰布支黑豆及其衍生的RIL群体γ-T含量进行混合遗传分析。主基因遗传率hmg2 (%) = σmg2/σp2,多基因遗传率hpg2 (%) = σpg2/σp2,环境遗传率he2 (%) = σe2/σp2。各遗传方差间的关系为σp2 = σmg2+σpg2+σe2。其中,σp2为群体表型方差;σmg2为主基因遗传方差;σpg2为多基因遗传方差;σe2为多环境遗传方差。
1.4 遗传连锁图谱的构建与QTL分析
王珍[26]使用晋豆23×灰布支黑豆杂交衍生的RIL群体为材料,构建了一个含有227个SSR标记的大豆遗传连锁图谱。以该图谱和大豆公共图谱中的标记为基础,梁慧珍[27]重新整合,整合后图谱全长2 047.6 cM,包括27个连锁群,平均图距8.8 cM,232个标记位点。该图谱在K(Chr.9)、F(Chr.13)和J(Chr.16)3条染色体上均出现2个间隙,在H(Chr.12)染色体出现了1个间隙,因而形成了27个连锁群。所得图谱与SONG等[28]整合的图谱排序相对一致。采用梁慧珍[27]整合后的大豆分子遗传图谱对大豆γ-T含量进行QTL定位分析。把不同年份作为环境因子处理,根据6个环境的表型数据,利用WinQTLCart 2.5软件中复合区间作图法(composite interval mapping,CIM)Zmapqtl方法的Model 6,在每个被测标记区间两侧设置10 cM的窗口(Window),同时设置该窗口之外的5个标记作为余因子,步移速度为2 cM,向前回归法。以1 000次排列测验(permutation test)、全基因组P=0.05的LOD值作为QTL存在的阈值。当QTL效应P≤0.05时,认为QTL存在。参照MCCOUCH等[29]方式命名QTL。
2 结果
2.1 大豆γ-T含量的表型分析和相关分析
RIL群体中γ-T含量的表型变异见图1和表1。6个环境中γ-T亲本差分别为-51.12和-48.77、-49.90和-51.23、-41.10和-44.26,平均差分别为-48.43和-48.09。不同环境中,亲本在后代分离群体中差异较大,RIL群体差异显著,表现出超亲分离。γ-T呈现连续分布特点,不同环境下遗传变异系数差别较小。3年间的遗传率分别为0.509、0.437和0.498,平均遗传率0.488。联合方差分析结果(表2)表明,γ-T含量在组分间差异显著,基因型和基因型×环境的互作方差显著,表明γ-T的遗传主要受基因型和基因型与环境互作的共同影响。图1
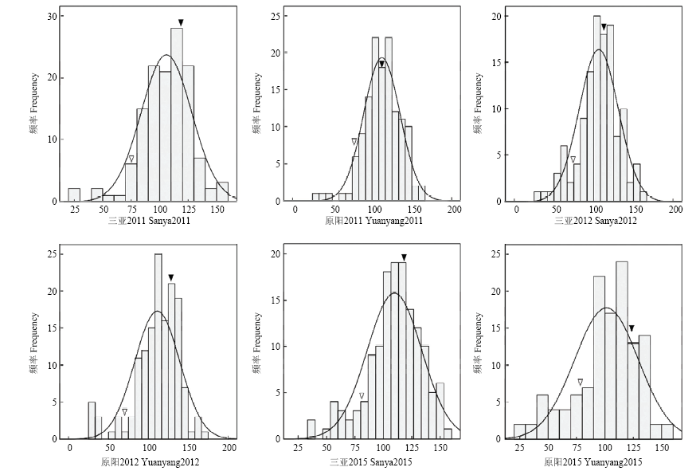 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1不同环境RIL群体γ-T含量分布图

Fig. 1Distribution of γ-T among the two parents and the RIL population across six environments
Table 1
表1
表1RIL群体大豆γ-T含量表型变异
Table 1
| 年份 Year | 平均值Mean (μg·g-1) | 亲本差 P2—P1 (μg·g-1) | RIL变幅RIL range (μg·g-1) | GCV (%) | 遗传率 h2 | ||||
|---|---|---|---|---|---|---|---|---|---|
| 原阳 Yuanyang | 三亚 Sanya | 原阳 Yuanyang | 三亚 Sanya | 原阳 Yuanyang | 三亚 Sanya | 原阳 Yuanyang | 三亚 Sanya | ||
| 2011 | 111.61 | 100.83 | -51.12 | -48.77 | 25.59—164.15 | 22.85—158.31 | 22.66 | 20.08 | 0.509 |
| 2012 | 109.92 | 101.18 | -49.90 | -51.23 | 33.65—152.19 | 27.65—161.02 | 20.07 | 16.21 | 0.437 |
| 2015 | 110.26 | 100.66 | -41.10 | -44.26 | 20.01—158.03 | 31.03—165.06 | 24.61 | 22.12 | 0.498 |
| 平均Mean | 110.60 | 100.88 | -48.43 | -48.09 | 20.01—164.15 | 22.85—165.06 | 22.59 | 19.65 | 0.488 |
新窗口打开|下载CSV
Table 2
表2
表2大豆种子γ-T含量方差分析
Table 2
| 性状 Trait | 变异来源 Variation | ||||||
|---|---|---|---|---|---|---|---|
| 基因型Genotypes | 环境Environment | 基因型×环境 Genotypes×Environment | 误差Error | ||||
| MS | F | MS | F | MS | F | MS | |
| γ-T | 14.1624 | 2.6972* | 0.0358 | 0.0068 | 16.6445 | 3.1698** | 5.2509 |
新窗口打开|下载CSV
相关性分析表明(表3),γ-T含量在2011年原阳和2011年三亚2个环境表现出显著相关,2012年原阳和2012年三亚2个环境表现出极显著相关,其他环境中均没有表现出显著相关性。说明γ-T含量受到基因型和环境的共同影响。相关性分析结果与γ-T含量在RIL群体中的分离及遗传分析结果基本一致。
Table 3
表3
表3大豆种子γ-T含量相关性分析
Table 3
| 环境 Environment | 原阳2011 Yuanyang2011 | 原阳2012 Yuanyang2012 | 原阳2015 Yuanyang2015 | 三亚2011 Sanya2011 | 三亚2012 Sanya2012 | 三亚2015 Sanya2015 |
|---|---|---|---|---|---|---|
| 原阳2011 Yuanyang2011 | 1 | |||||
| 原阳2012 Yuanyang2012 | -0.050 | 1 | ||||
| 原阳2015 Yuanyang2015 | 0.137 | 0.043 | 1 | |||
| 三亚2011 Sanya2011 | 0.417* | 0.023 | -0.006 | 1 | ||
| 三亚2012 Sanya2012 | 0.307 | 0.542** | 0.303 | 0.238 | 1 | |
| 三亚2015 Sanya2015 | 0.307 | -0.243 | 0.047 | 0.356 | 0.361 | 1 |
新窗口打开|下载CSV
2.2 γ-生育酚的主基因+多基因混合遗传分析
通过对2011年、2012年和2015年原阳和三亚共6个环境下试验数据的平均值进行主基因+多基因混合遗传分析,依据AIC值,结合均匀检验性检验、Kolmogorov试验和Smirnov检验选定最适遗传模型,并用最小二乘法估计γ-生育酚含量的遗传参数(表4)。γ-T最适遗传模型符合MX2-Duplicate-A,即2对重叠作用主基因×加性多基因模型。因为2对主基因间的重叠作用,三亚地点检测到2对主基因间上位性效应值0.4010—0.5169和多基因的加性效应值为0.1797—0.2146,主基因遗传率为11.27%—13.05%,多基因遗传率为82.51%—86.55%,说明γ-T存在多基因效应,且多基因效应大于主基因效应。但是原阳地点只检测到2对主基因间上位性效应0.9646—1.8455,主基因遗传率为39.51%—58.96%,没有检测出多基因效应。Table 4
表4
表4γ-生育酚最适模型及遗传参数估计结果
Table 4
| 参数 Parameter | γ-生育酚γ-T | 参数 Parameter | γ-生育酚 γ-T | |||||
|---|---|---|---|---|---|---|---|---|
| 原阳Yuanyang | 三亚Sanya | 原阳Yuanyang | 三亚Sanya | |||||
| 最适模型Optimal model | MX2-Duplicate-A | MX2-Duplicate-A | ||||||
| 一阶参数1st order parameter | 二阶参数2nd order parameter | |||||||
| M | 2011 | 8.6329 | 11.3909 | σp2 | 2011 | 6.3918 | 2.8311 | |
| 2012 | 10.3476 | 10.4479 | 2012 | 3.8596 | 3.2818 | |||
| 2015 | 7.0225 | 12.4529 | 2015 | 9.3853 | 3.0353 | |||
| iab(i*) | 2011 | 1.6711 | 0.4329 | σmg2 | 2011 | 3.4311 | 0.3695 | |
| 2012 | 0.9646 | 0.5169 | 2012 | 1.5250 | 0.4158 | |||
| 2015 | 1.8455 | 0.4010 | 2015 | 5.5337 | 0.3420 | |||
| [d] | 2011 | 0.1797 | σpg2 | 2011 | 2.3360 | |||
| 2012 | 0.2146 | 2012 | 2.7899 | |||||
| 2015 | 0.1921 | 2015 | 2.6270 | |||||
| h2mg (%) | 2011 | 53.68 | 13.05 | |||||
| 2012 | 39.51 | 12.67 | ||||||
| 2015 | 58.96 | 11.27 | ||||||
| hpg2 (%) | 2011 | 82.51 | ||||||
| 2012 | 84.99 | |||||||
| 2015 | 86.55 | |||||||
新窗口打开|下载CSV
2.3 大豆γ-T的QTL定位
根据WinQTLCart 2.5的定位结果,共在6个连锁群上检测到9个γ-生育酚的QTL(表5和图2),分别位于A1(Chr.5)、A2(Chr.8)、C1(Chr.4)、G(Chr.18)、K_2(Chr.9)和M(Chr.7)染色体上,解释的表型变异范围为7.29%—29.55%。加性效应值正负表现不一,加性效应值为正,说明等位基因来自于母本晋豆23,该QTL对大豆γ-生育酚含量起正向作用;加性效应值为负,说明等位基因来自于父本灰布支黑豆,该QTL对大豆γ-生育酚含量起负向作用。位于A1(Chr.5)染色体Satt364—Satt382标记区间的qγ-A1-2,表型贡献率为29.55%,解释的表型变异最大。其中,qγ-G-1同时在2011年原阳、2012年三亚、2015年三亚3个环境下检测到,且均定位在G(Chr.18)染色体Satt275—Satt038标记区间0.01 cM处,解释的表型变异分别为8.97%、8.12%和7.91%,说明这是一个能够稳定遗传的QTL。加性效应值-0.801—-0.535,增加γ-T含量的等位基因来自父本灰布支黑豆;qγ-A1-1同时在2011年原阳和2015年原阳2个环境下检测到,且均定位在A1(Chr.5)染色体Satt276—Satt364标记区间0.01 cM处,解释的表型变异分别为29.54%和28.23%,说明这可能是一个能够稳定遗传的QTL。加性效应值0.622和0.613,增加γ-T含量的等位基因来自母本晋豆23。同时,qγ-A1-2和qγ-A1-3均被定位在A1(Chr.5)染色体上,加性效应分别为0.621和-0.212,且2个QTL位置相距较远,推测可能不是同一个QTL,说明D1a(Chr.1)染色体上可能存在多个与γ-T相关的基因。Table 5
表5
表5γ-生育酚QTL位置及其参数
Table 5
| 年份 Year | 地点 Location | QTL | 染色体 Chr. | 标记区间 Marker interval | 位置 Position (cM) | 置信区间 Confidence interval | LOD | 加性效应 Additive | R2 (%) |
|---|---|---|---|---|---|---|---|---|---|
| 2011 | 原阳Yuanyang | qγ-A1-1 | A1(5) | Satt276—Satt364 | 12.01 | 7.01—14.35 | 3.71 | 0.622 | 29.54 |
| qγ-G-1 | G(18) | Satt275—Satt038 | 0.01 | 0.00—5.15 | 2.50 | -0.590 | 8.97 | ||
| 三亚Sanya | qγ-A2-1 | A2(8) | Satt315—Satt632 | 0.01 | 0.00—4.78 | 2.54 | -1.235 | 9.45 | |
| qγ-A1-2 | A1(5) | Satt364—Satt382 | 22.24 | 18.35—27.11 | 2.63 | 0.621 | 29.55 | ||
| 2012 | 原阳Yuanyang | qγ-K_2-1 | K_2(9) | Satt264—Sat_044 | 37.68 | 35.40—37.70 | 3.14 | -1.448 | 12.15 |
| qγ-A1-3 | A1(5) | Satt382—Satt593 | 55.77 | 48.12—56.69 | 3.18 | -0.212 | 14.20 | ||
| 三亚Sanya | qγ-A2-2 | A2(8) | Satt315—Satt632 | 2.01 | 0.67—6.17 | 2.73 | -0.711 | 14.31 | |
| qγ-G-1 | G(18) | Satt275—Satt038 | 0.01 | 0.00—4.73 | 2.56 | -0.535 | 8.12 | ||
| qγ-M-1 | M(7) | Satt220—Satt323 | 75.57 | 73.27—75.60 | 2.73 | -0.658 | 10.04 | ||
| 2015 | 原阳Yuanyang | qγ-C1-1 | C1(4) | Satt565—Satt578 | 18.82 | 18.80—28.97 | 2.99 | 0.355 | 7.29 |
| qγ-A1-1 | A1(5) | Satt276—Satt364 | 12.01 | 8.38—15.03 | 3.81 | 0.613 | 28.23 | ||
| 三亚Sanya | qγ-G-1 | G(18) | Satt275—Satt038 | 0.01 | 0.00—7.02 | 2.60 | -0.801 | 7.91 |
新窗口打开|下载CSV
图2
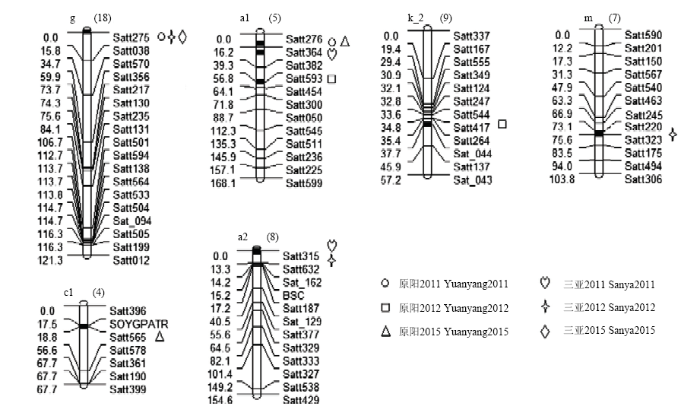 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2检测到的QTL及加性效应QTL在连锁群上的分布
Fig. 2Distribution of main QTLs and additive QTLs on linkage groups
3 讨论
3.1 主基因+多基因混合遗传分析和QTL定位结果的比较
本研究采用主基因+多基因混合遗传模型分离分析方法研究大豆γ-T在河南省和海南省2个地点3个年份共6个环境中的遗传;采用WinQTLCart 2.5软件中CIM方法进行QTL定位分析。2种分析结果相比较,发现QTL定位数目与遗传模型预测的数目并不完全一致。其中,2011年原阳和三亚、2012年原阳和2015年原阳环境下检测出2个QTL,与γ-T最适遗传模型MX2-Duplicate-A相吻合;在2012年三亚和2015年三亚2个环境下分别检测出3个和1个QTL,与遗传模型预测的数目不统一。说明2种分析方法既具有科学的统一性,又具有各自的局限性。分析其研究结果不完全统一的可能原因有:1)与混合遗传模型有关。本研究选用的是4对主基因+多基因混合遗传模型[30],该模型简化了主基因间的连锁作用。只分析了主基因间无连锁的情况。同时,分析过程中成分分布数和待估参数比较多,分析过程受到限制,估算结果的准确性有待提高;2)与QTL定位图谱有关。与标准图谱相比较,本研究选用的图谱还需要进一步完善;3)控制γ-T含量的表达在时空上存在差异。因此,对于γ-T含量的遗传机理研究,在本研究的基础上尚需进一步深化。3.2 大豆γ-T含量的QTL分析及与前人定位结果的比较
为提高QTL定位精度,得到表现稳定的QTL,采用多年多点数据分析方法是最有效的途径之一[31]。本研究采用的晋豆23×灰布支黑豆及其衍生的RIL群体γ-T定位的QTL中,qγ-G-1在3个环境下均被检测到,且均位于G(Chr.18)染色体Satt275—Satt038标记区间0.01 cM处,加性效应分别为-0.590、-0.535和-0.801,增加γ-T含量的等位基因来自父本灰布支黑豆;qγ-A1-1在2个环境下均被检测到定位在A1(Chr.5)染色体Satt276—Satt364标记区间0.01 cM处,加性效应分别为0.622和0.613,增加γ-T含量的等位基因来自母本晋豆23。可见qγ-G-1和qγ-A1-1可能是能够稳定表达的主效QTL,Satt275—Satt038和Satt276—Satt364 2个标记区间准确性较高。增加γ-T含量的2个QTLqγ-G-1和qγ-A1-1的等位基因分别来自父本和母本,与最适遗传模型符合MX2-Duplicate-A的分析结果相一致。下一步研究将针对本次γ-T基因定位的结果,在Satt275—Satt038和Satt276—Satt364连锁标记之间的序列上,寻找新的SSR标记,分析其在亲本间的多态性,进一步寻找与目的基因连锁更为紧密的SSR标记,进而实现γ-T基因的精细定位,为高γ-生育酚含量分子标记辅助育种奠定基础。生物的遗传和进化过程中,广泛存在多因一效和一因多效现象。这两种基因作用方式从不同角度共同促进生命活动的发现[32]。多因一效促成了生物多样性。不同的生物过程参与到同一种表型上,降低了单因素所带来的局限性,出现了多态性。一因多效将多种过程联系起来,共同参与基因表达过程,形成一系列生物轴,一方面促进行为的统一性,一方面形成正负反馈,避免生理生化过程的失稳。本研究发现,在6个连锁群上共检测到9个γ-T的QTL,加性效应值正负均有表现,这些等位基因一方面来自于母本晋豆23,一方面来自于父本灰布支黑豆,推测多因一效可能存在于γ-T的形成过程。同时也发现,本研究中稳定表达的标记Satt275,被认为与大豆孢囊线虫[33,34]和大豆染料木素形成[35]有关;Satt038与亚麻酸油合成[36]有关;Satt276与豆荚在灌浆期的相对磷浓度[37]和大豆结荚高度[38]有关;Satt364与产量、脂肪含量与根重有关[39];Satt382与侧根数有关[40]。说明控制这些性状的基因可能存在一因多效现象[41]。在大豆种子发育和营养生长不同时期,可以采取相应的措施调控γ-T的合成与积累。
QTL与环境互作、QTL与QTL上位性互作是数量性状遗传的重要组成部分,是作物杂种优势的遗传基础[42]。本研究中,在原阳和三亚2个地点均检测到上位性效应,上位性效应值最大为0.5169,加性效应最大为0.2147,上位效应的影响比加性更显著,与HAGIWARA等[43]和ZHANG等[44]认为在分析QTL定位时,上位性与上位性互作效应更有效,上位性比加性QTL更重要结论相一致。
下一步研究的重点方向之一是对这些QTL间的上位性反映机制和基因位点之间的表达调控机制进行深入研究。
与以往研究相比,尽管各研究选用的群体、图谱和环境条件不同,仍有一部分γ-T定位结果相吻合(表6)。与LI等[19,20]结果相比较,均定位在C1(Chr.4)、A2(Chr.8)和G(Chr.18)染色体上,且均定位在C1(Chr.4)染色体Satt565标记处;与SHAW等[21]结果相比较,均定位在C1(Chr.4)和K(Chr.9)染色体上;与李海燕[22]结果相比较,均定位在M(Chr.7)、K(Chr.9)和G(Chr.18)染色体上;与张红梅等[23]结果相比较,均定位在K(Chr.9)染色体上。但是,本研究中检测出的相对稳定遗传的qγ-G-1主效QTL,前人研究中虽然在这条染色体上也检测出γ-生育酚QTL,但是不在同一标记区间,定位位置是否重叠尚不能确定。分析其原因可能受到定位群体、定位图谱、遗传背景或环境差异等诸多因素影响。随着研究手段和使用图谱的不断进步和完善,逐步验证这些标记区间是否重叠。除此之外,本研究中定位出的其他QTL,均未见公开报道。
Table 6
表6
表6QTI定位结果比较
Table 6
| 亲本Parent | 性状Traits | 文献References |
|---|---|---|
| 晋豆23×灰布支黑豆Jindou23×Huibuzhi | A1(Satt276, Satt364, Satt382), A2(Satt315), C1(Satt565), G(Satt275), K(Satt264), M(Satt220) | 本研究This study |
| Bayfield×合丰25 Bayfield ×Hefeng 25 | A2(Sat_383), C1(Satt565), C2(Satt286), G(Satt199), D1b(Satt266), O(Satt576), J(Satt280) | [19] |
| Bayfield×合丰25 Bayfield ×Hefeng 25 | A2(Sat_383), C1(Satt565), C2(Satt286), G(Satt199), D1b(Satt266), O(Satt576), E(Satt355), J(Satt280) | [20] |
| Bayfield×Shire | C1(Satt646), E(Satt598), I(Satt354), F(Satt334), H(Satt279), K(Satt552) | [21] |
| Bayfield×合丰25 Bayfield ×Hefeng 25 | G(Sat_372), D1b(Satt266), L(Satt561, Satt527), M(Satt567) | [22] |
| Essex×ZDD2315 | C2(Satt489), D1a(Sat_346), D1b(AI856415), O(Satt633), K(Satt617, Satt260) | [23] |
新窗口打开|下载CSV
3.3 遗传图谱对QTL定位的影响
遗传图谱的构建在基因定位、基因的图位克隆、遗传资料多样性研究,以及遗传标记辅助选择育种等方面都具有重要意义。遗传图谱应用价值的大小,依赖于标记数量的多少或图谱的饱和度、标记在图谱上定位的准确性以及标记在图谱上分布的均匀程度等。一个基本的连锁框架图要求标记间平均间距20 cM以下。对于主基因定位,其平均距离要求在10—20 cM或更小[45]。连锁群标记较少,图谱上标记的密度较低时,影响QTL的检出效率,效应值较小的QTL有可能检测不到,进而影响定位的准确性[46]。同时,从另一角度分析,标记密度降低后仍然能够被检测到的QTL应该是效应较大的主效QTL,对分子标记辅助选择和数量性状基因的图位克隆更具实际价值。但这并不能与提高LOD的阈值注重主效QTL的检测同日而语,因为前者将付出降低QTL在染色体上定位的准确性的代价。本研究图谱是由海南省热带农业资源开发利用研究所方宣钧研究员提供(论文中致谢部分有说明),图谱总长度为2 047.6 cM,平均图距为8.8 cM[27],综合看来图谱平均间距尚能满足定位要求。下一步工作中,希望能进一步提高图谱分子标记密度和图谱饱和度,不断提高定位结果的精准度。尽管如此,利用该图谱定位了一批大豆数量性状的QTL,在国内外期刊发表,研究结果仍然具有一定的理论和现实意义。3.4 大豆γ-T遗传体系对育种的启示
通过对6个环境中大豆广义遗传力分析和γ-T方差分析研究,遗传力为43.68%—50.93%,基因型、基因型×环境均呈现出极显著差异,说明γ-T受遗传和环境因素的共同影响。主基因+多基因混合遗传模型分离分析表明,γ-T最适遗传模型符合MX2-Duplicate-A。说明在γ-T含量的遗传体系中,基因型、环境、上位性均起着重要作用,与张红梅等[23]研究结果γ-T含量的遗传主要受基因控制,环境影响程度较小的结论有所不同。WinQTLCart2.5软件QTL定位分析发现,2012年原阳、2013年原阳和三亚、2015年三亚4个环境中,每个环境下均定位出2个QTL,与主基因+多基因混合遗传模型预测的数目相吻合。但是,2012年三亚和2015年原阳分别检测出3个和1个QTL,与遗传模型MX2-Duplicate-A不一致。表明在γ-T含量的遗传中,一方面受基因型控制,表现出遗传的稳定性;一方面受环境和上位性的影响,表现出遗传的多样性和复杂性。几种不同的分析方法得出的γ-T含量的遗传结论基本一致,说明研究结果的准确性比较可靠。社会的发展带动了生活品质的提高,人们对于天然维生素E的需要越来越大。天然维生素E主要存在于植物的种子,但含量很低,提取成本较高,从育种的角度选育高含量维生素E大豆品种迫在眉睫。在各种不同形式的维生素E中,γ-T最常见,α-T生物活性最高。前人研究认为,γ-T甲基转移酶(γ-Topherol methytransferase,γ-TMT/VET4)是α-T合成途径最后一步甲基化酶,在γ-T甲基转移酶的催化作用下,以γ-T为底物转化合成α-T[47,48],通过超表达γ-T甲基转移酶基因可以实现种子中高含量γ-T向α-T的转化,从而提高种子中α-T的含量[49]。γ-T具有抗癌和防癌功效。目前,γ-T的抗癌和防癌研究已经取得重要进展,在γ-T的吸收和代谢[9,10]、γ-T抗癌作用及其分子机制[11,12,13]等方面的研究比较深入,在医学临床上得到广泛应用,取得了良好的效果。但是,实际生产中缺乏高维生素E含量特别是高γ-T含量的新品种,因此,本研究对于通过分子育种手段选育高维生素E含量大豆新品种特别是高γ-T大豆新品种,具有重要的参考价值。
作为育种工作者,进行分子标记辅助选择育种工作中,非常希望能够检测出大效应的QTL。本研究检测到的在2个环境中稳定表达的qγ-A1-1加性贡献率分别为29.54%和28.23%,但是,在3个环境中稳定表达的qγ-G-1加性贡献率均小于10%。如何聚合和转移包括微效基因在内的多基因,对育种工作者提出了新的课题。
4 结论
γ-T最适遗传模型符合MX2-Duplicate-A,即2对重叠作用主基因×加性多基因模型。其遗传同时受到基因型、环境和上位性的影响。检测到γ-T的2个稳定主效QTL,Satt275—Satt038和Satt276—Satt364是共位标记区间。致谢:
海南省热带农业资源开发利用研究所方宣钧研究员和山西省农业科学院经济作物研究所刘学义研究员提供了连锁图谱和群体材料,谨此致谢。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.3390/molecules15031746URL [本文引用: 1]
DOI:10.1104/pp.106.094771URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s00299-006-0207-5URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.1074/jbc.M201466200URL [本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
DOI:10.1007/s10681-013-0948-5URL [本文引用: 1]

Tocopherols have several beneficial effects in plants, and are indispensable micronutrients for humans. Sweet corn is a major source of tocopherols in high concentrations. In this investigation, tocopherol compounds in sweet corn were analyzed by high performance liquid chromatography. To detect quantitative trait loci (QTL) controlling accumulation of tocopherols at the milk stage in sweet corn, a F-2 population consisting of 229 F-2:3 lines was created from the cross between a high-total tocopherols line (A6) and a low-total tocopherols line (A57). A genetic map was constructed using 136 polymorphic molecular markers including one gene-targeted marker based on the tocopherol biosynthesis pathway (HPPD). Eleven putative QTLs for tocopherol content and composition were detected by composite interval mapping and located on Chr. 1, Chr. 2, Chr. 5, Chr. 6 and Chr. 10. Phenotypic variance explained by each QTL ranged from 4.74 to 41.16 %. Eight mapped QTLs were co-localized, suggesting that the same QTL affected the amounts of more than one tocopherol compound. One candidate gene-targeted marker (HPPD) showed co-localization with the major QTL for gamma-tocopherol and total tocopherols. Only one interval (umc1177-bnlg1429) on chromosome one exhibited a QTL for alpha, delta, gamma, and total tocopherols with high LOD and R-2 values. The primary conclusion of this work is that two major QTLs located on Chr. 1 and Chr. 5 can be used for improvement of sweet corn nutrition quality by marker-assisted selection.
[本文引用: 1]
DOI:10.1007/s11032-011-9585-7URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/pbr.2016.135.issue-2URL [本文引用: 2]
DOI:10.1007/s00122-010-1264-2URL [本文引用: 2]
Vitamin E (VE) in soybean seed has value for foods, medicines, cosmetics, and animal husbandry. Selection for higher VE contents in seeds along with agronomic traits was an important goal for many soybean breeders. In order to map the loci controlling the VE content, F5-derived F6 recombinant inbred lines (RILs) were advanced through single-seed-descent (SSD) to generate a population including 144 RILs. The population was derived from a cross between ‘OAC Bayfield’, a soybean cultivar with high VE content, and ‘Hefeng 25’, a soybean cultivar with low VE content. A total of 107 polymorphic simple sequence repeat markers were used to construct a genetic linkage map. Seed VE contents were analyzed by high performance liquid chromatography for multiple years and locations (Harbin in 2007 and 2008, Hulan in 2008 and Suihua in 2008). Four QTL associated with α-Toc (on four linkage groups, LGs), eight QTL associated with γ-Toc (on eight LGs), four QTL associated with δ-Toc (on four LGs) and five QTL associated with total VE (on four LGs) were identified. A major QTL was detected by marker Satt376 on linkage group C2 and associated with α-Toc (0.0012>P>0.0001, 5.0%<R 2<17.0%, 25.1<α-Toc<30.1μgg−1), total VE (P<0.0001, 7.0%<R 2<10.0%, 118.2<total VE<478.3μgg−1). A second QTL detected by marker Satt286 on LG C2 was associated with γ-Toc (0.0003>P>0.0001, 6.0%<R 2<13.0%, 141.5<γ-Toc<342.4μgg−1) and total VE (P<0.0001, 2.0%<R 2<9.0%, 353.9<total VE<404.0μgg−1). Another major QTL was detected by marker Satt266 on LG D1b that was associated with α-Toc (0.0002>P>0.0001, 4.0%<R 2<6.0%, 27.7<α-Toc<43.7μgg−1) and γ-Toc (0.0032>P>0.0001, 3.0%<R 2<10.0%, 69.7<γ-Toc<345.7μgg−1). Since beneficial alleles were all from ‘OAC Bayfield’, it was concluded that these three QTL would have great potential value for marker assisted selection for high VE content.
DOI:10.1111/pbr.2017.136.issue-1URL [本文引用: 2]
[D].
[本文引用: 2]
[D].
[本文引用: 2]
DOI:10.3724/SP.J.1006.2015.00187URL [本文引用: 3]
维生素E (VE)具有提高人体免疫力、抗癌、预防心血管疾病等保健作用,从大豆中提取的VE安全性更高。本研究采用高效液相色谱技术(HPLC)检测大豆BIEX群体(Essex×ZDD2315)维生素E的α-生育酚、γ-生育酚和δ-生育酚含量。应用QTLNetwork 2.1软件分别检测到8个和12对控制大豆维生素E及组分含量的加性和互作QTL。α-生育酚含量加性和互作QTL累计贡献值分别为8.68% (2个)和15.57% (4对),γ-生育酚含量加性和互作QTL累计贡献值分别为8.59% (2个)和11.57% (2对),δ-生育酚含量加性和互作QTL累计贡献值分别为5.44% (1个)和17.61% (3对),维生素E总含量的加性和互作QTL累计贡献值分别为11.39% (3个)和9.48% (3对)。未检测到维生素E及组分含量和环境互作的QTL。未定位到的微效QTL累计贡献值为66.16%~75.32%,说明未定位到的微效基因的变异占2/3以上。各性状的遗传构成中,未检测出的微效QTL份额最大,加性QTL和互作QTL贡献相差不大。在育种中应考虑常规方法聚合微效QTL与标记辅助方法聚合主要QTL相结合。
DOI:10.3724/SP.J.1006.2015.00187URL [本文引用: 3]
维生素E (VE)具有提高人体免疫力、抗癌、预防心血管疾病等保健作用,从大豆中提取的VE安全性更高。本研究采用高效液相色谱技术(HPLC)检测大豆BIEX群体(Essex×ZDD2315)维生素E的α-生育酚、γ-生育酚和δ-生育酚含量。应用QTLNetwork 2.1软件分别检测到8个和12对控制大豆维生素E及组分含量的加性和互作QTL。α-生育酚含量加性和互作QTL累计贡献值分别为8.68% (2个)和15.57% (4对),γ-生育酚含量加性和互作QTL累计贡献值分别为8.59% (2个)和11.57% (2对),δ-生育酚含量加性和互作QTL累计贡献值分别为5.44% (1个)和17.61% (3对),维生素E总含量的加性和互作QTL累计贡献值分别为11.39% (3个)和9.48% (3对)。未检测到维生素E及组分含量和环境互作的QTL。未定位到的微效QTL累计贡献值为66.16%~75.32%,说明未定位到的微效基因的变异占2/3以上。各性状的遗传构成中,未检测出的微效QTL份额最大,加性QTL和互作QTL贡献相差不大。在育种中应考虑常规方法聚合微效QTL与标记辅助方法聚合主要QTL相结合。
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 3]
[本文引用: 3]
DOI:10.1007/s00122-004-1602-3URL [本文引用: 1]
A total of 391 simple sequence repeat (SSR) markers designed from genomic DNA libraries, 24 derived from existing GenBank genes or ESTs, and five derived from bacterial artificial chromosome (BAC) end sequences were developed. In contrast to SSRs derived from EST sequences, those derived from genomic libraries were a superior source of polymorphic markers, given that the mean number of tandem repeats in the former was significantly less than that of the latter (P<0.01). The 420 newly developed SSRs were mapped in one or more of five soybean mapping populations:
 Minsoy
Minsoy × Noir 1, Minsoy × Archer, Archer × Noir 1, Clark × Harosoy, and A81-356022 × PI468916. The JoinMap software package was used to combine the five maps into an integrated genetic map spanning 2,523.6cM of Kosambi map distance across 20 linkage groups that contained 1,849 markers, including 1,015 SSRs, 709 RFLPs, 73 RAPDs, 24 classical traits, six AFLPs, ten isozymes, and 12 others. The number of new SSR markers added to each linkage group ranged from 12 to 29. In the integrated map, the ratio of SSR marker number to linkage group map distance did not differ among 18 of the 20 linkage groups; however, the SSRs were not uniformly spaced over a linkage group, clusters of SSRs with very limited recombination were frequently present. These clusters of SSRs may be indicative of gene-rich regions of soybean, as has been suggested by a number of recent studies, indicating the significant association of genes and SSRs. Development of SSR markers from map-referenced BAC clones was a very effective means of targeting markers to marker-scarce positions in the genome.
× Noir 1, Minsoy × Archer, Archer × Noir 1, Clark × Harosoy, and A81-356022 × PI468916. The JoinMap software package was used to combine the five maps into an integrated genetic map spanning 2,523.6cM of Kosambi map distance across 20 linkage groups that contained 1,849 markers, including 1,015 SSRs, 709 RFLPs, 73 RAPDs, 24 classical traits, six AFLPs, ten isozymes, and 12 others. The number of new SSR markers added to each linkage group ranged from 12 to 29. In the integrated map, the ratio of SSR marker number to linkage group map distance did not differ among 18 of the 20 linkage groups; however, the SSRs were not uniformly spaced over a linkage group, clusters of SSRs with very limited recombination were frequently present. These clusters of SSRs may be indicative of gene-rich regions of soybean, as has been suggested by a number of recent studies, indicating the significant association of genes and SSRs. Development of SSR markers from map-referenced BAC clones was a very effective means of targeting markers to marker-scarce positions in the genome.[本文引用: 1]
DOI:10.3724/SP.J.1006.2010.00191URL [本文引用: 1]
主基因加多基因混合遗传模型分离分析方法是基于数量性状表型数据的统计遗传分析方法,该方法适于育种工作者利用杂种分离世代的数据对育种性状的遗传组成做出初步判断,制订相应的育种策略,也可用以校验QTL定位所揭示的性状遗传组成。重组自交家系群体(RIL)是一种永久性群体,可以进行有重复的比较试验,适合于环境影响较大的复杂性状的遗传研究。本研究以RIL群体为对象,将遗传模型拓展到4对主基因,建立相应的分离分析方法。新构建的模型共包括4对主基因和4对主基因加多基因两类共15个遗传模型,通过极大似然法和IECM算法估算各种模型的分布参数,由AIC值和一组适合性测验选取最佳遗传模型,再由最小二乘法估计模型的遗传参数。通过模拟实验对所建立的模型进行验证,模拟群体中一阶遗传参数的估计值与设定值之间有很好一致性。以大豆科丰1号×南农1138-2构成的RIL群体及其亲本棕榈酸含量的遗传(I-1模型,即4对加性-上位性主基因和加性-上位性多基因遗传模型)为例,说明该方法的应用效果。
DOI:10.3724/SP.J.1006.2010.00191URL [本文引用: 1]
主基因加多基因混合遗传模型分离分析方法是基于数量性状表型数据的统计遗传分析方法,该方法适于育种工作者利用杂种分离世代的数据对育种性状的遗传组成做出初步判断,制订相应的育种策略,也可用以校验QTL定位所揭示的性状遗传组成。重组自交家系群体(RIL)是一种永久性群体,可以进行有重复的比较试验,适合于环境影响较大的复杂性状的遗传研究。本研究以RIL群体为对象,将遗传模型拓展到4对主基因,建立相应的分离分析方法。新构建的模型共包括4对主基因和4对主基因加多基因两类共15个遗传模型,通过极大似然法和IECM算法估算各种模型的分布参数,由AIC值和一组适合性测验选取最佳遗传模型,再由最小二乘法估计模型的遗传参数。通过模拟实验对所建立的模型进行验证,模拟群体中一阶遗传参数的估计值与设定值之间有很好一致性。以大豆科丰1号×南农1138-2构成的RIL群体及其亲本棕榈酸含量的遗传(I-1模型,即4对加性-上位性主基因和加性-上位性多基因遗传模型)为例,说明该方法的应用效果。
DOI:10.1007/BF00220855URL [本文引用: 1]
DOI:10.1007/s10681-006-9085-8URL [本文引用: 1]
[本文引用: 1]
DOI:10.1007/s00122-018-3063-0URL [本文引用: 1]
DOI:10.3724/SP.J.1006.2015.01372URL [本文引用: 1]
以栽培大豆晋豆23为母本, 以山西农家品种大豆灰布支黑豆为父本杂交衍生的447个RIL作为供试群体构建遗传图谱, 利用高效液相色谱法定性、定量测定样品中的异黄酮及其组分含量。采用主基因+多基因混合遗传分离分析法和WinQTLCart 2.5复合区间作图法, 对大豆异黄酮及其组分含量进行混合遗传分析和QTL定位。结果表明, 大豆苷、黄豆苷元、染料木素、染料木苷、大豆苷元和异黄酮总含量分别受4、4、2、3、2和2对主基因控制, 并有多基因修饰。检测到44个与大豆异黄酮及其组分含量相关的QTL, 与大豆苷、染料木素、黄豆苷元、大豆苷元、染料木苷和异黄酮总含量相关的QTL分别有10、9、4、7、8和6个。连续2年分别检测到与大豆苷、染料木苷、黄豆苷元和异黄酮关联, 分别位于标记区间satt430~satt359、satt038~satt570、satt197~sat_128和satt249~satt285的稳定表达QTL, 可尝试用于分子标记辅助育种。
DOI:10.3724/SP.J.1006.2015.01372URL [本文引用: 1]
以栽培大豆晋豆23为母本, 以山西农家品种大豆灰布支黑豆为父本杂交衍生的447个RIL作为供试群体构建遗传图谱, 利用高效液相色谱法定性、定量测定样品中的异黄酮及其组分含量。采用主基因+多基因混合遗传分离分析法和WinQTLCart 2.5复合区间作图法, 对大豆异黄酮及其组分含量进行混合遗传分析和QTL定位。结果表明, 大豆苷、黄豆苷元、染料木素、染料木苷、大豆苷元和异黄酮总含量分别受4、4、2、3、2和2对主基因控制, 并有多基因修饰。检测到44个与大豆异黄酮及其组分含量相关的QTL, 与大豆苷、染料木素、黄豆苷元、大豆苷元、染料木苷和异黄酮总含量相关的QTL分别有10、9、4、7、8和6个。连续2年分别检测到与大豆苷、染料木苷、黄豆苷元和异黄酮关联, 分别位于标记区间satt430~satt359、satt038~satt570、satt197~sat_128和satt249~satt285的稳定表达QTL, 可尝试用于分子标记辅助育种。
DOI:10.1007/s10681-014-1264-4URL [本文引用: 1]
DOI:10.1371/journal.pgen.1004061URL [本文引用: 1]
DOI:10.3724/SP.J.1006.2014.00037URL [本文引用: 1]
以栽培大豆晋豆23为母本, 半野生大豆灰布支黑豆ZDD2315为父本杂交衍生的F2:15和F2:16的447个RIL家系为遗传群体, 绘制SSR遗传图谱, 采用混合线性模型方法, 对2年大豆小区产量及主要农艺性状进行加性QTL、加性×加性上位互作及环境互作分析。结果检测到9个与小区产量、茎粗、有效分枝、主茎节数、株高、结荚高度相关的QTL, 分别位于J_2、I、M连锁群上, 其中小区产量、茎粗、株高、有效分枝和主茎节数QTL的加性效应为正值, 说明增加这些性状的等位基因来源于母本晋豆23。同时, 检测到7对影响小区产量、茎粗、株高和结荚高度的加性×加性上位互作效应及环境互作效应的QTL, 共发现14个与环境存在互作的QTL。上位效应和QE互作效应对大豆小区产量及主要农艺性状的遗传影响较大。大豆分子标记辅助育种中, 既要考虑起主要作用的QTL, 又要注重上位性QTL, 才有利于性状的稳定表达和遗传。
DOI:10.3724/SP.J.1006.2014.00037URL [本文引用: 1]
以栽培大豆晋豆23为母本, 半野生大豆灰布支黑豆ZDD2315为父本杂交衍生的F2:15和F2:16的447个RIL家系为遗传群体, 绘制SSR遗传图谱, 采用混合线性模型方法, 对2年大豆小区产量及主要农艺性状进行加性QTL、加性×加性上位互作及环境互作分析。结果检测到9个与小区产量、茎粗、有效分枝、主茎节数、株高、结荚高度相关的QTL, 分别位于J_2、I、M连锁群上, 其中小区产量、茎粗、株高、有效分枝和主茎节数QTL的加性效应为正值, 说明增加这些性状的等位基因来源于母本晋豆23。同时, 检测到7对影响小区产量、茎粗、株高和结荚高度的加性×加性上位互作效应及环境互作效应的QTL, 共发现14个与环境存在互作的QTL。上位效应和QE互作效应对大豆小区产量及主要农艺性状的遗传影响较大。大豆分子标记辅助育种中, 既要考虑起主要作用的QTL, 又要注重上位性QTL, 才有利于性状的稳定表达和遗传。
DOI:10.2135/cropsci1999.3951464xURL [本文引用: 1]
DOI:10.3864/j.issn.0578-1752.2017.18.002URL [本文引用: 1]
【目的】主根长和侧根数是重要的根系性状。通过不同氮磷钾处理,发掘大豆苗期主根长和侧根数的基因资源、了解其遗传机制,定位其主效QTL,分析QTL间的上位性和环境互作效应,对生产提供理论指导。【方法】以栽培大豆晋豆23为母本、山西农家品种灰布支黑豆(ZDD02315)为父本所衍生的447个RIL作为供试群体,取亲本及447个家系各30粒种子,用灭菌纸包裹后,2015年和2016年分别放置于CK(模拟种植不施肥)、NPK(模拟大田正常配施氮磷钾肥)和1.5NPK(模拟高肥田块)3种生长环境下进行水培试验,每组试验设置3次重复,环境温度20—28℃,幼苗长到V2期,对幼苗期相关根部性状数据进行测量。分别采用WinQTLCart 2.5和QTLNETwork 2.1 2种遗传模型检测QTL,分析QTL间的上位性和环境互作效应。【结果】基于复合区间作图(CIM)共检测到24个影响主根长和侧根数的QTL,分布于第2、3、5、6、7、8、9、10、11、12、13、14、16、17共14条染色体中,单个QTL的贡献率介于8.52%—43.62%,QTL主要表现为加性效应。基于混合线性模型(MCIM)检测到影响主根长和侧根数的QTL各1个,2个QTL均表现出加性效应和环境互作效应。另有2对主根长和2对侧根数均检测出加性×加性上位性互作QTL,主根长和侧根数各有1对表现出主效QTL与非主效QTL加性×加性上位性互作,各有1对表现出非主效QTL与非主效QTL加性×加性上位性互作,2对主根长互作QTL分别解释了1.53%和1.95%的表型变异率,2对侧根数互作QTL分别解释了2.47%和1.13%的表型变异率。2个QTL能在2种分析方法中同时检测到,9个QTL能在3种环境下同时检测到。第6染色体在2015年NPK、1.5NPK和2016年1.5NPK 3个环境下均检测到主根长QTL,第5染色体在2015年NPK和1.5NPK、2016年CK 3个环境下、第17染色体在2015年CK和NPK、2016年NPK 3个环境下均检测到侧根数QTL。【结论】苗期大豆主根长和侧根数对氮磷钾的吸收影响较少,生产中尽可能减少氮磷钾使用量。不同浓度氮磷钾处理苗期主根长和侧根数参数间既有共同的控制基因,也有各自独特的控制基因,多数QTL不能在多个环境下重复检测到,控制其表达的遗传机制较为复杂。加性效应、加性与环境互作和加性×加性上位性互作效应在主根长和侧根数的形成和遗传中发挥着重要作用。主根长和侧根数各有1个QTL能在2种分析方法中同时检测到,Satt442-Satt296和Satt521-GMABABR是共位标记区间。
DOI:10.3864/j.issn.0578-1752.2017.18.002URL [本文引用: 1]
【目的】主根长和侧根数是重要的根系性状。通过不同氮磷钾处理,发掘大豆苗期主根长和侧根数的基因资源、了解其遗传机制,定位其主效QTL,分析QTL间的上位性和环境互作效应,对生产提供理论指导。【方法】以栽培大豆晋豆23为母本、山西农家品种灰布支黑豆(ZDD02315)为父本所衍生的447个RIL作为供试群体,取亲本及447个家系各30粒种子,用灭菌纸包裹后,2015年和2016年分别放置于CK(模拟种植不施肥)、NPK(模拟大田正常配施氮磷钾肥)和1.5NPK(模拟高肥田块)3种生长环境下进行水培试验,每组试验设置3次重复,环境温度20—28℃,幼苗长到V2期,对幼苗期相关根部性状数据进行测量。分别采用WinQTLCart 2.5和QTLNETwork 2.1 2种遗传模型检测QTL,分析QTL间的上位性和环境互作效应。【结果】基于复合区间作图(CIM)共检测到24个影响主根长和侧根数的QTL,分布于第2、3、5、6、7、8、9、10、11、12、13、14、16、17共14条染色体中,单个QTL的贡献率介于8.52%—43.62%,QTL主要表现为加性效应。基于混合线性模型(MCIM)检测到影响主根长和侧根数的QTL各1个,2个QTL均表现出加性效应和环境互作效应。另有2对主根长和2对侧根数均检测出加性×加性上位性互作QTL,主根长和侧根数各有1对表现出主效QTL与非主效QTL加性×加性上位性互作,各有1对表现出非主效QTL与非主效QTL加性×加性上位性互作,2对主根长互作QTL分别解释了1.53%和1.95%的表型变异率,2对侧根数互作QTL分别解释了2.47%和1.13%的表型变异率。2个QTL能在2种分析方法中同时检测到,9个QTL能在3种环境下同时检测到。第6染色体在2015年NPK、1.5NPK和2016年1.5NPK 3个环境下均检测到主根长QTL,第5染色体在2015年NPK和1.5NPK、2016年CK 3个环境下、第17染色体在2015年CK和NPK、2016年NPK 3个环境下均检测到侧根数QTL。【结论】苗期大豆主根长和侧根数对氮磷钾的吸收影响较少,生产中尽可能减少氮磷钾使用量。不同浓度氮磷钾处理苗期主根长和侧根数参数间既有共同的控制基因,也有各自独特的控制基因,多数QTL不能在多个环境下重复检测到,控制其表达的遗传机制较为复杂。加性效应、加性与环境互作和加性×加性上位性互作效应在主根长和侧根数的形成和遗传中发挥着重要作用。主根长和侧根数各有1个QTL能在2种分析方法中同时检测到,Satt442-Satt296和Satt521-GMABABR是共位标记区间。
DOI:10.3864/j.issn.0578-1752.2019.01.002URL [本文引用: 1]
【目的】 通过对大豆α-生育酚进行遗传和QTL分析,研究其遗传机制,定位其主效QTL,为高α-生育酚含量的大豆品种选育奠定遗传学基础。【方法】 以栽培大豆晋豆23为母本、山西农家品种大豆灰布支黑豆(ZDD02315)为父本杂交衍生的447个RIL作为供试群体构建遗传图谱,试验群体及亲本分别于2011年、2012年和2015年夏季在河南省农业科学院原阳试验基地种植,冬季在海南省三亚南繁基地种植。田间试验采取随机区组设计,2次重复。从6个环境中每个家系选取15.00 g籽粒饱满,大小一致的大豆种子,利用高效液相色谱法定性、定量测定样品中的α-生育酚含量。采用主基因+多基因混合遗传分离分析法和WinQTLCart 2.5复合区间作图法,对大豆α-生育酚含量进行主基因+多基因混合遗传分析和QTL定位。【结果】 基于主基因+多基因混合遗传分离分析法,α-生育酚受4对主基因控制,遗传基因分布在双亲中。4对主基因间加性效应值中3对为正值,表明这些基因来源于母本晋豆23;1对为负值,表明该对基因来源于父本灰布支黑豆;4对主基因之间相互作用的上位性效应表现为正值和负值的各有3对,说明不同基因间上位性效应对α-TOC的影响方向并不完全一致。环境因素引起的变异为0.13%—4.05%。表明α-TOC主要受4对主基因影响,受环境因素影响较小。采用WinQTLCart 2.5复合区间作图(CIM)共检测到17个影响α-生育酚的QTL,分布于第1、2、5、6、8、14、16、17共8条染色体中,单个QTL的贡献率8.35%—35.78%,QTL主要表现为加性效应。qα-D1a-1同时在2011年原阳、2012年原阳和三亚、2015年原阳4个环境下检测到,且均定位在第1染色体Satt320—Satt254标记区间19.79 cM处,解释的表型变异分别为12.55%、12.01%和11.89%、12.61%,加性效应值0.119-0.132,增加α-TOC含量的等位基因来自母本晋豆23;qα-A2-1同时在2011年原阳和三亚、2015年原阳3个环境下检测到,且均定位在第8染色体Sat_129—Satt377标记区间44.53 cM处,解释的表型变异分别为23.18%和22.56%、23.01%,加性效应值-0.195—-0.180,增加α-TOC含量的等位基因来自父本灰布支黑豆。qα-D1a-1和qα-A2-1 2个QTL能够稳定遗传。【结果】 α-生育酚最适遗传模型符合4MG-AI,即4对具有加性上位性效应的主基因遗传模型。其遗传主要受4对主基因影响,受环境因素影响较小。检测到α-生育酚的2个稳定主效QTL,Satt320—Satt254和Sat_129—Satt377是共位标记区间。
DOI:10.3864/j.issn.0578-1752.2019.01.002URL [本文引用: 1]
【目的】 通过对大豆α-生育酚进行遗传和QTL分析,研究其遗传机制,定位其主效QTL,为高α-生育酚含量的大豆品种选育奠定遗传学基础。【方法】 以栽培大豆晋豆23为母本、山西农家品种大豆灰布支黑豆(ZDD02315)为父本杂交衍生的447个RIL作为供试群体构建遗传图谱,试验群体及亲本分别于2011年、2012年和2015年夏季在河南省农业科学院原阳试验基地种植,冬季在海南省三亚南繁基地种植。田间试验采取随机区组设计,2次重复。从6个环境中每个家系选取15.00 g籽粒饱满,大小一致的大豆种子,利用高效液相色谱法定性、定量测定样品中的α-生育酚含量。采用主基因+多基因混合遗传分离分析法和WinQTLCart 2.5复合区间作图法,对大豆α-生育酚含量进行主基因+多基因混合遗传分析和QTL定位。【结果】 基于主基因+多基因混合遗传分离分析法,α-生育酚受4对主基因控制,遗传基因分布在双亲中。4对主基因间加性效应值中3对为正值,表明这些基因来源于母本晋豆23;1对为负值,表明该对基因来源于父本灰布支黑豆;4对主基因之间相互作用的上位性效应表现为正值和负值的各有3对,说明不同基因间上位性效应对α-TOC的影响方向并不完全一致。环境因素引起的变异为0.13%—4.05%。表明α-TOC主要受4对主基因影响,受环境因素影响较小。采用WinQTLCart 2.5复合区间作图(CIM)共检测到17个影响α-生育酚的QTL,分布于第1、2、5、6、8、14、16、17共8条染色体中,单个QTL的贡献率8.35%—35.78%,QTL主要表现为加性效应。qα-D1a-1同时在2011年原阳、2012年原阳和三亚、2015年原阳4个环境下检测到,且均定位在第1染色体Satt320—Satt254标记区间19.79 cM处,解释的表型变异分别为12.55%、12.01%和11.89%、12.61%,加性效应值0.119-0.132,增加α-TOC含量的等位基因来自母本晋豆23;qα-A2-1同时在2011年原阳和三亚、2015年原阳3个环境下检测到,且均定位在第8染色体Sat_129—Satt377标记区间44.53 cM处,解释的表型变异分别为23.18%和22.56%、23.01%,加性效应值-0.195—-0.180,增加α-TOC含量的等位基因来自父本灰布支黑豆。qα-D1a-1和qα-A2-1 2个QTL能够稳定遗传。【结果】 α-生育酚最适遗传模型符合4MG-AI,即4对具有加性上位性效应的主基因遗传模型。其遗传主要受4对主基因影响,受环境因素影响较小。检测到α-生育酚的2个稳定主效QTL,Satt320—Satt254和Sat_129—Satt377是共位标记区间。
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s10681-006-9085-8URL [本文引用: 1]
It has been theoretically proposed that multiple linked quantitative trait loci (QTLs) play a role in the accumulation of hidden variation within and between populations. In this study, the genetic bases for grain characteristics were examined by comparing two accessions representing the two rice subspecies by QTL analysis. Grain dimensions are known to be quantitative traits and to be diagnostic between these two subspecies. To enhance the power to detect QTL with small effects, after transferring a segment of chromosome 6 from an Indica type into a Japonica type of rice by repeated backcrosses, the introgressed segment was dissected by making recombinant inbred lines (RILs) which were expected to have different sizes of the introgressed segment in the same genetic background. The resulting RILs showed distinct transgression of the grain characteristics examined. Multiple QTLs controlling each of the length and breadth of seeds were detected on the introgressed segment, and showed positive and negative additive effects as well as epistatic interactions. The present study confirmed that transgressive segregation resulted from a breakdown of linkage and that the detection of QTLs was highly dependent upon the genetic effects of the neighboring QTLs, indicating the need for caution in interpreting QTL effects.
DOI:10.1016/j.fcr.2004.06.004URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/BF00220989URL [本文引用: 1]
[本文引用: 1]
DOI:10.1046/j.1432-1033.2003.03364.xURL [本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}