Comparative Analysis of miRNA Expression Profiles in the Hearts of Tibetan Cattle and Xuanhan Cattle
CHEN LuLu, WANG Hui, WANG JiKun, WANG JiaBo, CHAI ZhiXin, CHEN ZhiHua,, ZHONG JinCheng,Institute of Tibetan Plateau Research, Southwest Minzu University/Key Laboratory of Qinghai-Tibetan Plateau Animal Genetic Resource Reservation and Utilization, Sichuan Province and Ministry of Education, Chengdu 610041
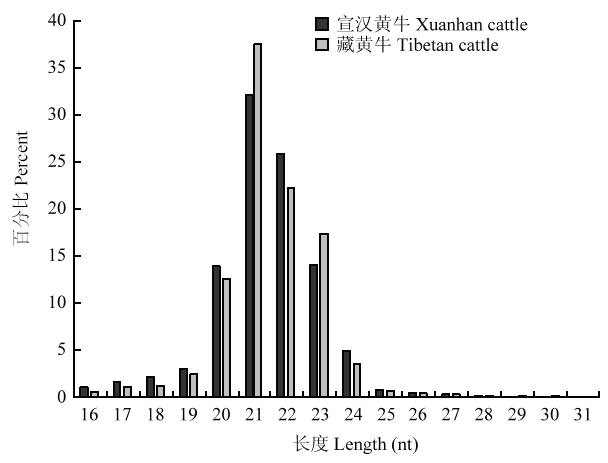
Abstract 【Objective】As a kind of non-coding RNA, miRNA is widely involved in various life activities of the organism. This study was aimed to explore the differential expression profiles of miRNA in the heart tissues between Tibetan cattle and Xuanhan cattle, so as to provide the basic data for further study on molecular mechanism of hypoxia adaptation in Tibetan cattle. 【Method】Each three healthy Tibetan and Xuanhan cattle were randomly selected for heart tissue sampling. RNA was extracted from tissues using the Trizol method. An 18 to 30nt fragment was selected by agarose gel electrophoresis, and 3' connector and 5' liner was ligated and then the fragment was enlarged. After gel electrophoresis, three Tibetan cattle and Xuanhan cattle libraries were established, respectively. High-throughput sequencing was performed by using the Illumina HiSeq4000 sequencing platform. The sequence was then filtered and the differentially expressed miRNA of Tibetan cattle and Xuanhan cattle were screened by comparing GenBank and Rfam databases. Functional annotation and signal pathway enrichment analysis of differentially expressed miRNA in Tibetan cattle and Xuanhan cattle. Finally, in order to verify the accuracy of the sequencing data, 8 miRNAs were randomly selected and the expression level of miRNA was detected by RT-qPCR. 【Result】The results showed that Tibetan cattle and Xuanhan cattle had high-quality reads of 17 463 446 and 13 662 812, respectively, while the clean reads were 16 552 296 and 12 055 304, respectively. The highest enrichment of high-quality nucleic acid sequences in Tibetan cattle and Xuanhan cattle were 21 nt, which were 37.5% and 32.1%, respectively. A total of 219 differential expressed miRNAs (48 up-regulated and 171 down-regulated) were obtained. There were 22 terms in the GO function annotation that significantly enriched in the molecular function of differentially expressed miRNAs target genes, such as GO: 0005488 (binding), GO: 0005515 (protein binding) and GO: 0043167 (ion binding). GO: 0005623 (cell), GO: 0044464 (cell component) and GO: 0005622 (cell) were among the 20 terms, which were significantly enriched in the cellular components. While there were 13 terms, which were significantly enriched in biological processes, such as GO: 0035556 (intracellular signal transduction), GO: 0032774 (RNA biosynthesis process) and GO: 0006351 (transcription, DNA templated). Analysis of KEGG signaling pathways revealed that miRNA target genes were significantly enriched to 232 signaling pathways, including the insulin signaling pathway (139 target genes), the mTOR signaling pathway (38 target genes) and the HIF-1 signaling pathway (92 target genes). Among them, 12 miRNA target genes worked together on these three signaling pathways. These results suggested that the differentially expressed miRNAs might participate in the regulation of hypoxia adaptation in Tibetan cattle through these three signaling pathways. Eight miRNAs were randomly selected for RT-qPCR, and the expression profiles were consistent with the sequencing data, indicating that the high-throughput sequencing data was reliable. 【Conclusion】Taken together, the expression profiles of miRNAs in the heart tissues of Tibetan and Xuanhan cattle were obtained in the present study, which laid a foundation for further research on the hypoxia adaptation mechanism of Tibetan cattle. Keywords:Tibetan cattle;Xuanhan cattle;miRNA;heart;high-throughput sequencing;hypoxia adaptability
PDF (1271KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 陈露露, 王会, 王吉坤, 王嘉博, 柴志欣, 陈智华, 钟金城. 藏黄牛与宣汉黄牛心脏miRNA表达谱比较[J]. 中国农业科学, 2020, 53(8): 1677-1687 doi:10.3864/j.issn.0578-1752.2020.08.016 CHEN LuLu, WANG Hui, WANG JiKun, WANG JiaBo, CHAI ZhiXin, CHEN ZhiHua, ZHONG JinCheng. Comparative Analysis of miRNA Expression Profiles in the Hearts of Tibetan Cattle and Xuanhan Cattle[J]. Scientia Acricultura Sinica, 2020, 53(8): 1677-1687 doi:10.3864/j.issn.0578-1752.2020.08.016
Fig. 3Quantitative histogram of GO enrichment of miRNA target genes in Tibetan cattle and Xuanhan cattle(A :Biological process B: Molecular function C: Cellular component)
PANDEY RK, SUNDARS, PRAJAPAATI VK . Differential expression of miRNA regulates T Cell differentiation and plasticity during visceral leishmaniasis infection Frontiers in Microbiology, 2016, 7(25):206-215. [本文引用: 1]
FULLAONDOA, LEE SY . Identification of putative miRNA involved in Drosophila melanogaster immune response Developmental and Comparative Immunology, 2012,36(2):267-273. [本文引用: 1]
FUX, MENGZ, LIANGW, TIANY, WANGX, HANW, LOUG, WANGX, LOUF, YENY, YUH, JOVER, HUANGW . MiR-26a enhances miRNA biogenesis by targeting Lin28B and Zcchc11 to suppress tumor growth and metastasis Oncogene, 2014,33(34):4296-4306. [本文引用: 1]
CHENGY, XIANGG, MENGY, DONGR . MiRNA-183-5p promotes cell proliferation and inhibits apoptosis in human breast cancer by targeting the PDCD4 Reproductive Biology, 2016, 16(3): 225-233. [本文引用: 1]
HESSAMS, SANDM, SKRYGANM, GAMBICHLERT, BECHARA FG . Expression of miRNA-155, miRNA-223, miRNA-31, miRNA-21, miRNA-125b, and miRNA-146a in the Inflammatory Pathway of hidradenitis suppurativa Inflammation, 2017,40(2):464-472. [本文引用: 1]
MALC, AFTABUDDINM, KUNDUS . IIKmTA: Inter and intra kingdom miRNA-target analyzer Interdisciplinary Sciences, Computational Life Sciences, 2018,10(3):538-543. [本文引用: 1]
JONASS, LZAURRALDEE . Towards a molecular understanding of microRNA-mediated gene silencing Nature Reviews Genetics, 2015,16(7):421-433. [本文引用: 1]
CIMMINOA, CALINGA, FABBRIM, LORIO MV, FERRACINM, SHIMIZUM, WOJCIK SE, AQEILAN RI, ZUPOS, DONOM, RASSENTIL, ALDERH, VOLINIAS, LIU CG, KIPPS TJ, NEGRINIM, CROCE CM . MiR-15 and miR-16 induce apoptosis by targeting BCL2 Proceedings of the National Academy of Sciences of the United States of America, 2005, 102(39):13944-13949. [本文引用: 1]
NALLAMSHETTYS, CHAN SY, LOSCALZOJ . Hypoxia: a master regulator of microRNA biogenesis and activity Free Radical Biology Medicine, 2013,64:20-30. [本文引用: 1]
YAOM, WANGX, TANGY, ZHANGW, CuiB, LiuQ, XingL . Dicer mediating the expression of miR-143 and miR-155 regulates hexokinase II associated cellular response to hypoxia American Journal of Physiology-Lung Cellular Physiology, 2014,307(11):829-837. [本文引用: 1]
XUX, LIUC, BAOJ . Hypoxia-induced hsa-miR-101 promotes glycolysis by targeting TIGAR mRNA in clear cell renal cell carcinoma Molecular Medicine Reports, 2017,15(3):1373-1378. [本文引用: 1]
ZHANGG, YINS, MAOJ, LIANGF, ZHAOC, LIP, ZHOUG, CHENS, TANGZ . Integrated analysis of mRNA-seq and miRNA-seq in the liver of Pelteobagrus vachelli in response to hypoxia Scientific Reports, 2016,10(6):22907. [本文引用: 1]
ZHANG YY . Effect and mechanism of bovine miR-378 in preadipocyte differention [D]. Changchun: Jilin University, 2014. (in Chinese) [本文引用: 1]
LI HJ, LIUM, YES, YANGF . De novo assembly, gene annotation, and molecular marker development using Illumina paired-end transcriptome sequencing in the clam Saxidomus purpuratus Genes & Genomics, 2017,39(6):675-685. [本文引用: 1]
GUO SX, LIU YN . Comparative study on pulmonary vascular reactivity of plateau adapted animal Yak and common cattle Chinese Journal of Pathophysiology, 1995,11(3):230-233. (in Chinese) [本文引用: 1]
JIN CY, Lü XY, GAOW, WANGY, CHEN WH, SHEN SX, CHENL, LINJ, SUNW . Study on the relationship between the expression of candidate miRNAs and the developmental characteristics in different patterns in Hu Sheep Lambskin Scientia Agricultura Sinica, 2018,51(14):2814-2824. (in Chinese) [本文引用: 1]
NALLAMSHETTYS, CHAN SY, LOSCALZOJ . Hypoxia: a master regulator of microRNA biogenesis and activity Free Radical Biology & Medicine, 2013,64:20-30. [本文引用: 1]
BURNSIDEJ, OUYANGM, ANDERSONA, BERNBERGE, LUC, MEYERS BC, GREEN PJ, MARKISM, ISAACSG, HUANGE, MORGAN RW . Deep sequencing of chicken microRNAs BMC Genomics, 2008,9(1):185. [本文引用: 1]
LORENZO FR, HUFFC, MYLLYMAKIM, OLENCHOCKB, SWIERCZEKS, TASHIT, GORDEUKV, WURENT, RI-LIG, MCCLAIN DA, KHAN TM, KOUL PA, GUCHHAITP, SALAMA ME, XINGJ, SEMENZA GL, LIBERZONE, WILSONA, SIMONSON TS, JORDE LB, KAELIN W GJ, KOIVUNENP, PRCHAL JT . A genetic mechanism for Tibetan high-altitude adaptation Nature Genetics, 2014,46(9):951-956. [本文引用: 1]
QUY, ZHAOH, HANN, ZHOUG, SONGG, GAOB, TIANS, ZHANGJ, ZHANGR, MENGX, ZHANGY, ZHANGY, ZHUX, WANGW, LAMBERTD, ERICSON PG, SUBRAMANIANS, YEUNGC, ZHUH, JIANGZ, LIR, LEIF . Ground tit genome reveals avian adaptation to living at high altitudes in the Tibetan plateau Nature Communications, 2013,4:2071. [本文引用: 1]
VASUDEVANS, TONGY, STEITZ JA . Switching from repression to activation: microRNAs can up-regulate translation Science, 2007, 318(5858):1931-1934. [本文引用: 1]
HE DF . Identification and differential expression analysis of microRNAs in heart and lung tissues between yak and cattle [D]. Chengdu: Sichuan Agricultural University, 2017. (in Chinese) [本文引用: 1]
ZHANGQ, GOUW, WANGX, ZHANGY, MAJ, ZHANGH, ZHANGY, ZHANGH . Genome resequencing identifies unique adaptations of Tibetan Chickens to hypoxia and high-dose ultraviolet radiation in high-altitude environments Genome Biology and Evolution, 2016,8(3):765-776. [本文引用: 1]
LIUJ, LI JD . Research progress of mTOR signaling pathway effect in cardiac development and reconstruction Advances in Cardiovascular Diseases, 2018(06):911-915. (in Chinese) [本文引用: 1]
SCIARRETTAS, ZHAIP, MAEJIMAY, DEL R DP, NAGARAJANN, YEED, LIUT, MAGNUSON MA, VOLPEM, FRATIG, LIH, SADOSHIMAJ . mTORC2 regulates cardiac response to stress by inhibiting MST1 Cell Reports, 2015,11(1):125-136. [本文引用: 1]
VERMAP, SHARMAA, SODHIM, THAKURK, KATERIA RS, NIRANJAN SK, BHARTI VK, KUMARP, GIRIA, KALIAS, MUKESHM . Transcriptome analysis of circulating pbmcs to understand mechanism of high altitude adaptation in native cattle of ladakh region Scientific Reports, 2018,8(1):7681. [本文引用: 1]
VERMAP, SHARMAA, SODHIM, THAKURK, BHARTI VK, KUMARP, GIRIA, KALIAS, SWAMI SK, MUKESHM . Overexpression of genes associated with hypoxia in cattle adapted to trans himalayan region of Ladakh Cell Biology International, 2018,42(9):1141-1148. [本文引用: 1]
TAGUCHIA, YANAGISAWAK, TANAKAM, CAOK, MATSUYAMAY, GOTOH, TAKAHASHIT . Identification of hypoxia-inducible factor-1 alpha as a novel target for miR-17-92 microRNA cluster Cancer Research, 2008, 68(14):5540-5545. [本文引用: 1]
BEFANI CD, VLACHOSTERRGIOS PJ, HATZIDAKIE, PATRIKIDOUA, BONANOUS, SIMOSG, PAPANDREOU CN, LIAKOSP . Bortezomib represses HIF-1α protein expression and nuclear accumulation by inhibiting both PI3K/Akt/TOR and MAPK pathways in prostate cancer cells Journal of Molecular Medicine, 2012,90(1):45-54. [本文引用: 1]
MAJMUNDARR AJ, WONG WJ, SIMON MC . Hypoxia-inducible factors and the response to hypoxic stress Molecular Cell, 2010,40(2):294-309. [本文引用: 1]
TAGUCHIA, YANAGISAWAK, TANAK AM, CAOK, MATSUYAMAY, GOTOH, TAKAHASHIT . Identification of hypoxia-inducible factor-1 alpha as a novel target for miR-17-92 microRNA cluster Cancer Research, 2008,68(14):5540-5545. [本文引用: 1]
CASCIOS, D'ANDREAA, FERLAR, SURMACZE, GULOTTAE, AMODEOV, BAZANV, GEBBIAN, RUSSOA . MiR-20b modulates VEGF expression by targeting HIF-1 alpha and STAT3 in MCF-7 breast cancer cells Journal of Cellular Physiology,2010,224(1):242-249. [本文引用: 1]
YUEY, Lü FH, CHEN YL, WANGZ, SI AY . Effect of miR-499 on apoptosis of primary cardiomyocytes induced by anoxia-reoxygenation Journal of Zhengzhou University(Medical Sciences), 2018,53(04):503-507. (in Chinese) [本文引用: 1]
ZHAOX . MicroRNA-101a inhibits hypoxia-induced myocardial fibrosis by targeting regulation of cardiac fibroblast TGFβRI expression [D]. Wuhan:Huazhong University of Science and Technology, 2015. (in Chinese) [本文引用: 1]
XI TY, JINF, ZHUY, WANGJ, TANGL, WANGY, LIEBESKKIND DS, HEZ . MicroRNA-126-3p attenuates blood-brain barrier disruption, cerebral edema and neuronal injury following intracerebral hemorrhage by regulating PIK3R2 and Akt Biochemical and Biophysical Research Communications, 2017,494(1-2):144-151. [本文引用: 1]
 ,, 钟金城
,, 钟金城
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT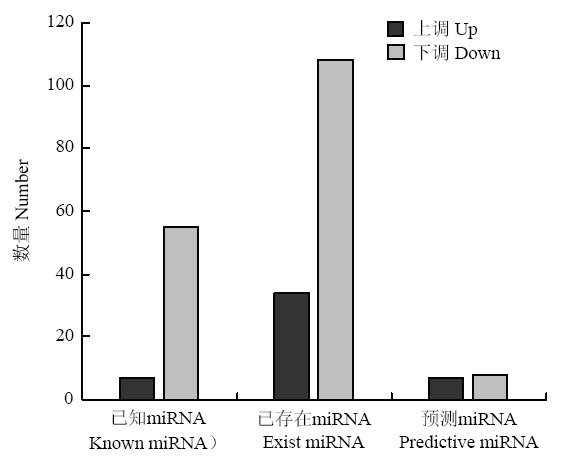 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT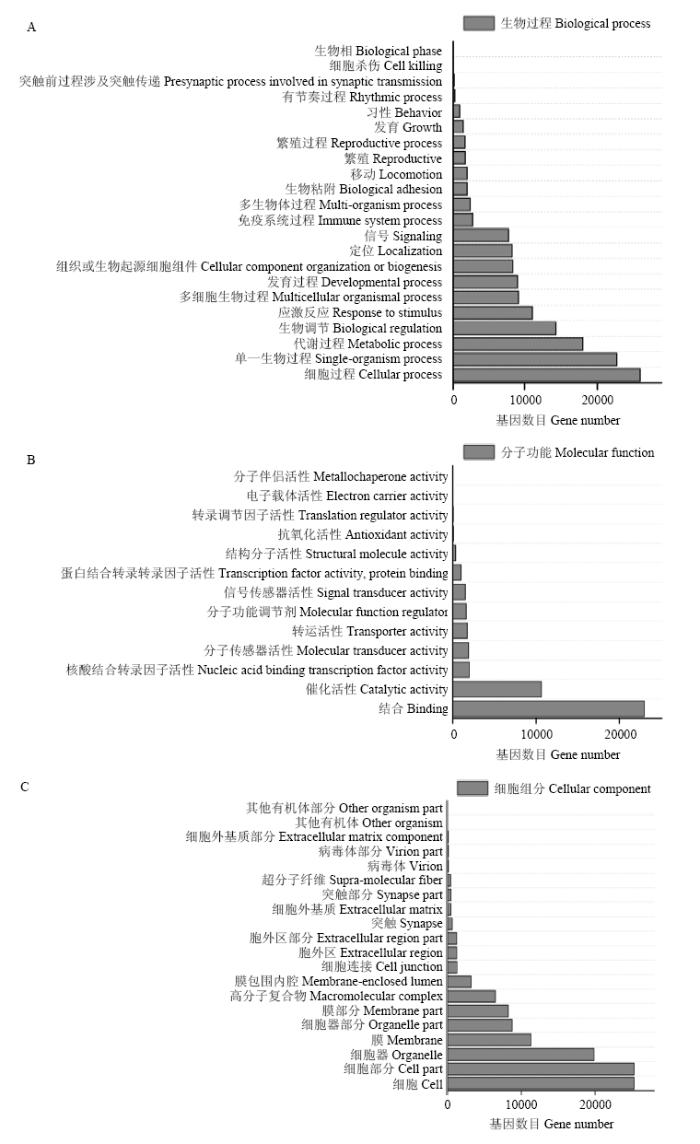 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT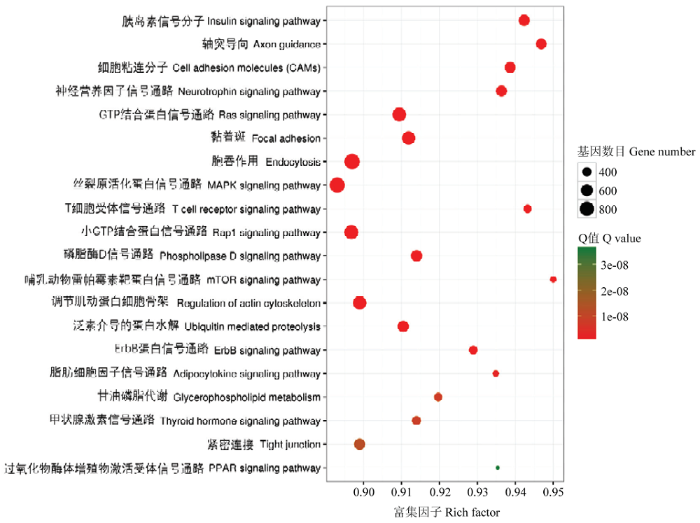 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT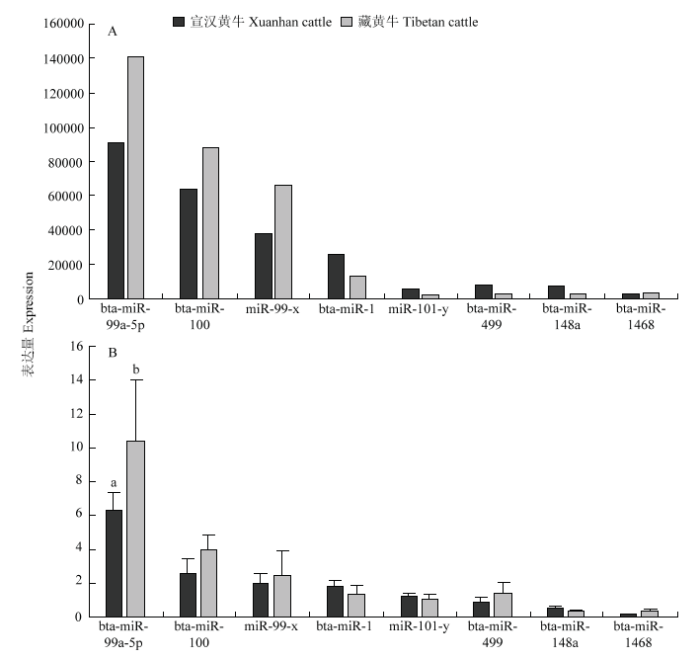 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}