Biologic and Transcriptomic Analysis of Citrus hystrix Responses to ‘Candidatus Liberibacter asiaticus’ at Different Infection Stages
TENG CaiLing1, ZHONG Xi1, WU HaoDi1, HU Yan2, ZHOU ChangYong1, WANG XueFeng11. National Citrus Engineering Research Center, Citrus Research Institute, Chinese Academy of Agricultural Sciences, Chongqing 400712 2. Ganzhou Bureau of Fruit Industry, Ganzhou 341000, Jiangxi
Abstract 【Objective】 Citrus Huanglongbing (HLB), associated with phloem-colonized ‘Candidatus Liberibacter asiaticus’ (CLas), severely impedes worldwide citrus production. The objective of this study is to analyze the biological symptoms, microstructures and transcriptomes of Citrus hystrix in response to CLas infection at different stages, to reveal the tolerance mechanism of C. hystrix, and to provide a basis for further screening of resistance genes and HLB-tolerant/resistant citrus breeding. 【Method】| The budwoods of C. hystrix grafted on two-year-old Carrizo citrange rootstocks (C. sinensis× P. trifoliata) used in this study were graft-inoculated with budwoods from a CLas (strain GZBJT)-infected Guanximiyou pummelo maintained in a greenhouse at Citrus Research Institute. The budwoods used as inoculum were tested CLas positive and free of other potential phloem-limited pathogens, such as Citrus tristeza virus (CTV) or Citrus tatter leaf virus (CTLV) by PCR before grafting. Inoculated plants were kept in greenhouse along with mock-inoculated healthy control plants. Real-time quantitative PCR (qPCR) was performed every 15 days after inoculation. Four months after inoculation (the earliest establishment of CLas by qPCR) and 14 months after inoculation were defined as the early infection stage and the late infection stage, respectively. The biological symptoms and microstructures were observed to analyze the structural changes of different infection stages. Combined with comparative transcriptome and RT-qPCR validation, the response mechanism of C. hystrix against HLB was explored.【Result】 No typical symptom was observed in C. hystrix at the early and late stages of infection. Light microscopy observation from the midribs of HLB-affected and uninfected C. hystrix revealed that no significant structure change was found at the early infection stage and only a few sieves in the phloem were blocked at the late stage. By comparing the RNA-seq data, 181 and 1 384 genes were found to be differentially expressed at the early stage and late stage, respectively. Differentially expressed genes (DEGs) mainly involved in cell wall metabolism, host defense response, starch and sucrose metabolism, callose synthesis and signal transduction. Comparative transcriptome analysis showed that the expression of related genes in starch and sucrose metabolism and cell wall metabolism was down-regulated at the early infection stage, and the expression of related genes in salicylic acid metabolism, salicylic acid signal transduction pathway, pathogenesis-related protein and glutathione-S-transferases was up-regulated at the late infection stage.【Conclusion】 The early response of C. hystrix to CLas infection is mainly characterized by stable physical structure, undisturbed pathways such as starch synthesis and photosynthesis. Salicylic acid-mediated resistance signals, effector-triggered immunity (ETI), and glutathione-S-transferases mediated detoxification contribute to the tolerance of C. hystrix against CLas at the late infection stage. Keywords:citrus Huanglongbing;‘Candidatus Liberibacter asiaticus’ (CLas);;Citrus hystrix;disease tolerance;transcriptome
PDF (2873KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 滕彩玲, 钟晰, 吴昊娣, 胡燕, 周常勇, 王雪峰. 马蜂柑响应黄龙病菌不同侵染时期的生物学和转录组学分析[J]. 中国农业科学, 2020, 53(7): 1368-1380 doi:10.3864/j.issn.0578-1752.2020.07.007 TENG CaiLing, ZHONG Xi, WU HaoDi, HU Yan, ZHOU ChangYong, WANG XueFeng. Biologic and Transcriptomic Analysis of Citrus hystrix Responses to ‘Candidatus Liberibacter asiaticus’ at Different Infection Stages[J]. Scientia Acricultura Sinica, 2020, 53(7): 1368-1380 doi:10.3864/j.issn.0578-1752.2020.07.007
A:健康植株Mock plants;B:感病前期CLas-infection at the early stage;C:感病后期CLas-infection at the late stage;D:健康对照叶中脉Leaf midrib of healthy control;E:感病前期叶中脉CLas-infected leaf midrib at the early stage;F:感病后期的叶中脉CLas-infected leaf midrib at the late stage;G:D局部放大照片Magnification of D;H:E局部放大照Magnification of E;I:F局部放大照片Magnification of F;Co:皮层细胞Cortex;Fi:纤维细胞Fiber;Ph:韧皮部Phloem;Pi:髓心细胞Pith;X:木质部Xylem;Sc:分泌腔Secretory cavity;图中箭头所指的蓝色斑点代表堵塞的筛管The blue spot indicated by arrow represents phloem plugging Fig. 1Phenotypic identification and microstructure observation of C. hystrix infected by CLas at different stages
CH-M-VS-CH-HLB-E:马蜂柑染病前期与健康对照相比DEGs in CLas-infected C. hystrix at the early stage compared with healthy control; CH-M-VS-CH-HLB-L:马蜂柑染病后期与健康对照相比DEGs in CLas-infected C. hystrix at the late stage compared with healthy control。下同The same as below红色表示上调,绿色表示下调 Red and green indicate up-regulated and down-regulated, respectively Fig. 2Comparative PageMan display of perturbed pathway of DEGs in C. hystrix infected by CLas at different stages
彩色方块代表上调或下调表达的差异基因Colored squares indicate up- or down-regulated genes。图4同The same as Fig. 4 Fig. 3DEGs involved in sucrose-starch metabolism (A) and photosynthesis (B)
ETXEBERRIAE, GONZALEZP, ACHORD, ALBRIGOG . Anatomical distribution of abnormally high levels of starch in HLB-affected Valencia orange trees Physiological and Molecular Plant Pathology, 2009,74(1):76-83. [本文引用: 2]
ALBRECHTU, BOWMAN K D. Transcriptional response of susceptible and tolerant citrus to infection with ‘Candidatus Liberibacter asiaticus’ Plant Science, 2012,185/186:118-130. [本文引用: 1]
BOAVA LP, CRISTOFANI-YALYM, MACHADO MA . Physiologic, anatomic, and gene expression changes in Citrus sunki, Poncirus trifoliata and their hybrids after ‘Candidatus Liberibacter asiaticus’ infection Phytopathology, 2017,107(5):590-599. [本文引用: 1]
FOLIMONOVA SY, ROBERTSON CJ, GARNSEY SM, GOWDAS, DAWSON WO . Examination of the responses of different genotypes of citrus to Huanglongbing (citrus greening) under different conditions Phytopathology, 2009,99(12):1346-1354. [本文引用: 2]
SHOKROLLAHH, ABDULLAH TL, SIJAMK, ABDULLAH S N A, ABDULLAH N A P. Differential reaction of citrus species in Malaysia to Huanglongbing (HLB) disease using grafting method American Journal of Agricultural and Biological Science, 2009,4(1):338. [本文引用: 1]
MARTINELLIF, URATSU SL, ALBRECHTU, REAGAN RL, PHU ML, BRITTONM, BUFFALOV, FASSJ, LEICHTE, ZHAOW, LIND, D’SOUZA R, DAVIS C E, BOWMAN K D, DANDEKAR A M. Transcriptome profiling of citrus fruit response to Huanglongbing disease PLoS ONE, 2012,7(5):e38039. [本文引用: 2]
ALBRECHTU, BOWMAN KD . Gene expression in Citrus sinensis(L.) Osbeck following infection with the bacterial pathogen ‘Candidatus Liberibacter asiaticus’ causing Huanglongbing in Florida Plant Science, 2008,175(3):291-306. [本文引用: 1]
KIM JS, SAGARAM US, BURNS JK, LI JL, WANGN . Response of sweet orange ( Citrus sinensis) to ‘Candidatus Liberibacter asiaticus’ infection: Microscopy and microarray analyses Phytopathology, 2009,99(1):50-57. [本文引用: 1]
FANJ, CHENC, BRLANSKY RH, GMITTER FG, LI ZG . Changes in carbohydrate metabolism in Citrus sinensis infected with ‘Candidatus Liberibacter asiaticus’ Plant Pathology, 2010,59(6):1037-1043. [本文引用: 2]
FANJ, CHENC, YUQ, BRLANSKY RH, LI ZG, GMITTER FG . Comparative iTRAQ proteome and transcriptome analyses of sweet orange infected by ‘ Candidatus Liberibacter asiaticus’ Physiologia Plantarum, 2011,143(3):235-245. [本文引用: 1]
ALBRECHTU, FIEHNO, BOWMAN KD . Metabolic variations in different citrus root stock cultivars associated with different responses to Huanglongbing Plant Physiology and Biochemistry, 2016,107:33-44. [本文引用: 1]
ARITUAV, ACHORD, GMITTER FG, ALBRIGOG, WANGN . Transcriptional and microscopic analyses of citrus stem and root responses to ‘ Candidatus Liberibacter asiaticus’ infection PLoS ONE, 2013,8(9):e73742. [本文引用: 2]
ZHONGY, CHENG CZ, JIANGB, JIANG NH, ZHANG YY, HU ML, ZHONG GY . Digital gene expression analysis of Ponkan mandarin (Citrus reticulata Blanco) in response to Asia citrus psyllid-vectored Huanglongbing infection International Journal of Molecular Sciences, 2016,17(7):1063. [本文引用: 4]
FANJ, CHENC, YUQ, KHALAFA, ACHOR DS, BRLANSKY RH, MOORE GA, LI ZG, GMITTER FG . Comparative transcriptional and anatomical analyses of tolerant rough lemon and susceptible sweet orange in response to ‘ Candidatus Liberibacter asiaticus’ infection Molecular Plant-Microbe Interactions, 2012,25(11):1396-1407. [本文引用: 4]
ZHONGY, CHENG CZ, JIANG NH, JIANGB, ZHANG YY, WUB, HU ML, ZENG JW, YAN HX, YI GJ, ZHONG GY . Comparative transcriptome and iTRAQ proteome analyses of citrus root responses to ‘ Candidatus Liberibacter asiaticus’ infection PLoS ONE, 2015,10(6):e0126973. [本文引用: 2]
LIAO HL, BURNS JK . Gene expression in Citrus sinensis fruit tissues harvested from Huanglongbing-infected trees: Comparison with girdled fruit Journal of Experimental Botany, 2012,63(8):3307-3319. [本文引用: 2]
DUAN YP, ZHOU LJ, HALL DG, LI WB, DODDAPANENIH, LINH, LIUL, VAHLING CM, GABRIEL DW, WILLIAMS KP, DICKERMANA, SUN YJ, COTTWALDT . Complete genome sequence of citrus Huanglongbing bacterium, ‘ Candidatus Liberibacter asiaticus’ obtained through metagenomics Molecular Plant-Microbe Interactions, 2009,22(8):1011-1020. [本文引用: 1]
LUH, ZHANGC, ALBRECHTU, SHIMIZUR, WANGG, BOWMAN KD . Overexpression of a citrus NDR1 ortholog increases disease resistance in Arabidopsis Frontiers in Plant Science, 2013,4:157. [本文引用: 1]
ROSALESR, BURNS JK . Phytohormone changes and carbohydrate status in sweet orange fruit from Huanglongbing-infected trees Journal of Plant Growth Regulation, 2011,30(3):312-321. [本文引用: 1]
NEHELAY, HIJAZF, ELZAAWELY AA, EL-ZAHABY H M, KILLINY N. Citrus phytohormonal response to ‘ Candidatus Liberibacter asiaticus’ and its vector Diaphorina citri Physiological and Molecular Plant Pathology, 2018,102:24-35. [本文引用: 1]
HUY, ZHONGX, LIU XL, LOU BH, ZHOU CY, WANG XF . Comparative transcriptome analysis unveils the tolerance mechanisms of Citrus hystrix in response to ‘Candidatus Liberibacter asiaticus’ infection PLoS ONE, 2017,12(12):e0189229. [本文引用: 2]
KOH EJ, ZHOUL, WILLIAMS DS, PARKJ, DINGN, DUAN YP, KANG BH . Callose deposition in the phloem plasmodesmata and inhibition of phloem transport in citrus leaves infected with ‘ Candidatus Liberibacter asiaticus’ Protoplasma, 2012,249(3):687-697. [本文引用: 2]
FANJ, CHENC, ACHOR DS, BRLANSKY RH, LI ZG, GMITTER FG . Differential anatomical responses of tolerant and susceptible citrus species to the infection of ‘ Candidatus Liberibacter asiaticus’ Physiological and Molecular Plant Pathology, 2013,83:69-74. [本文引用: 1]
DENGH, ACHOR DS, EXTEBERRIAE, YUQ, DUD, STANTOND, LIANGG, GMITTERFG . Phloem regeneration is a mechanism for Huanglongbing-tolerance of ‘Bearss’ lemon and ‘LB8-9’ sugar Belle? mandarin Frontiers in Plant Science, 2019,10:277. [本文引用: 1]
WANG YS, ZHOU LJ, YU XY, STOVERE, LUOF, DUAN YP . Transcriptome profiling of Huanglongbing (HLB) tolerant and susceptible citrus plants reveals the role of basal resistance in HLB tolerance Frontiers in Plant Science, 2016,7:933. [本文引用: 2]
LIONETTIV . PECTOPLATE: The simultaneous phenotyping of pectin methylesterases, pectinases, and oligogalacturonides in plants during biotic stresses Frontiers in Plant Science, 2015,6:331. [本文引用: 1]
OWENS RA, BLACKBURNM, DINGB . Possible involvement of the phloem lectin in long-distance viroid movement Molecular Plant-Microbe Interactions, 2001,14(7):905-909. [本文引用: 2]
FOLIMONOVA SY, ACHOR DS . Early events of citrus greening (Huanglongbing) disease development at the ultrastructural level Phytopathology, 2010,100(9):949-958. [本文引用: 1]
CHISHOLM ST, COAKERG, DAYB, STASKAWICA BJ . Host-microbe interactions: Shaping the evolution of the plant immune response Cell, 2006,124(4):803-814. [本文引用: 1]
JONES JD, DANGL JL . The plant immune system Nature, 2006,444(7117):323-329. [本文引用: 1]
ZOUH, GOWDAS, ZHOUL, HAJERIS, CHENG, DUANY . The destructive citrus pathogen, ‘ Candidatus Liberibacter asiaticus’ encodes a functional flagellin characteristic of a pathogen-associated molecular pattern PLoS ONE, 2012,7(9):e46447. [本文引用: 1]
SHI QC, FEBRES VJ, ZHANG SJ, YU FH, MCCOLLUMG, HALL DG, MOORE GA, STOVERE . Identification of gene candidates associated with Huanglongbing tolerance, using ‘ Candidatus Liberibacter asiaticus’ flagellin 22 as a proxy to challenge citrus Molecular Plant-Microbe Interactions, 2017,31(2):200-211. [本文引用: 1]
DA GRACA JV, DOUHAN GW, HALBERT SE, KEREMANE ML, LEE RF, VIDALAKISG, ZHAOH . Huanglongbing: An overview of a complex pathosystem ravaging the world’s citrus Journal of Integrative Plant Biology, 2016,58(4):373-387. [本文引用: 1]
ZHANGY, LIX . Salicylic acid: Biosynthesis, perception, and contributions to plant immunity Current Opinion in Plant Biology, 2019,50:29-36. [本文引用: 1]
ZHUF, XI DH, YUANS, XUF, ZHANG DW, LIN HH . Salicylic acid and jasmonic acid are essential for systemic resistance against Tobacco mosaic virus in Nicotiana benthamiana Molecular Plant-Microbe Interactions, 2014,27(6):567-577. [本文引用: 1]
ZOUX, BAIX, WENQ, XIEZ, WUL, PENGA, HEY, XUL, CHENS . Comparative analysis of tolerant and susceptible citrus reveals the role of methyl salicylate signaling in the response to Huanglongbing Journal of Plant Growth Regulation, 2019,38:1516-1528. [本文引用: 1]
VAN LOON LC, REPM, PIETERSE C MJ . Significance of inducible defense-related proteins in infected plants Annual Reviews of Phytopathology, 2006,44(1):135-162. [本文引用: 1]
WUJ, ALFREZ FM, JOHNSON EG, GRAHAM JH . Up-regulation of PR 1 and less disruption of hormone and sucrose metabolism in roots is associated with lower susceptibility to ‘ Candidatus Liberibacter asiaticus’ Plant Pathology, 2018,67(6):1426-1435. [本文引用: 1]
FUS, SHAOJ, ZHOUC, HARTUNG JS . Transcriptome analysis of sweet orange trees infected with ‘ Candidatus Liberibacter asiaticus’ and two strains of Citrus tristeza virus BMC Genomics, 2016,17:349. [本文引用: 1]
NWUGO CC, DUANY, LINH . Study on citrus response to Huanglongbing highlights a down-regulation of defense-related proteins in lemon plants upon ‘ Ca. Liberibacter asiaticus’ infection PLoS ONE, 2013,8(6):e67442. [本文引用: 1]
MARTINELLIF, REAGAN RL, DOLAND, FILECCIAV, DANDEKAR AM . Proteomic analysis highlights the role of detoxification pathways in increased tolerance to Huanglongbing disease BMC Plant Biology, 2016,16(1):167. [本文引用: 1]


 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT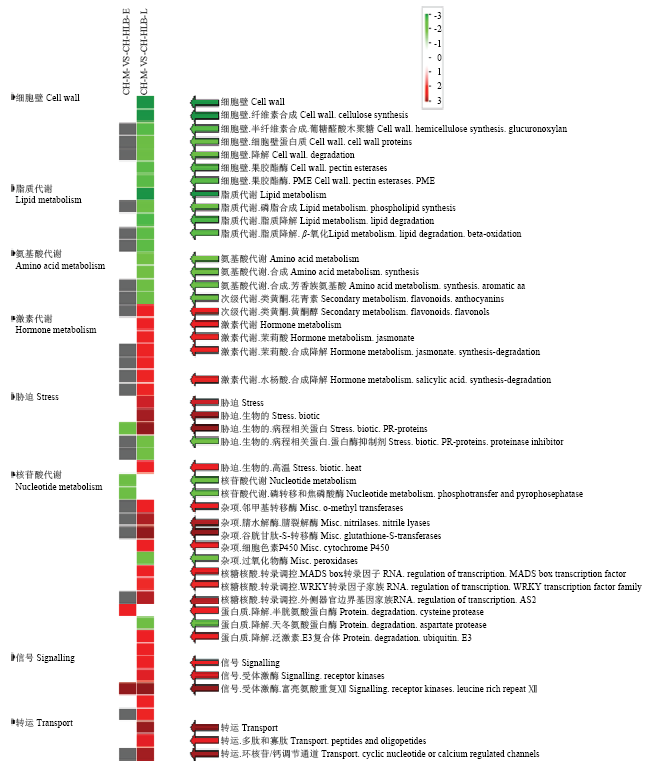 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT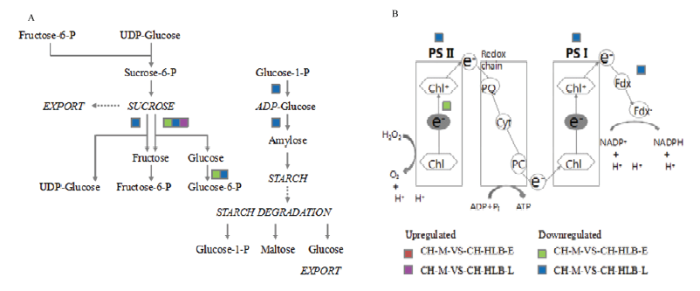 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT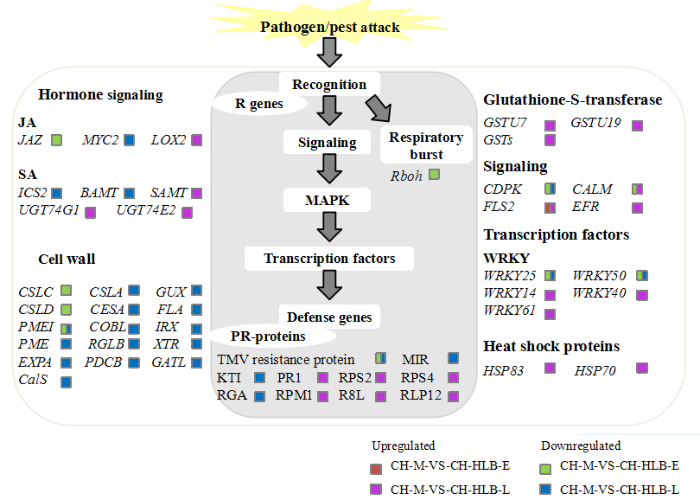 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}