2.
3.
Effects of Heat Stress on Physiological Indexes and Ultrastructure of Grapevines
LIU Min1,2, FANG Yulin1,31. 2.
3.
责任编辑: 赵伶俐
收稿日期:2019-08-27接受日期:2019-10-30网络出版日期:2020-04-01
| 基金资助: |
Editor:
Received:2019-08-27Accepted:2019-10-30Online:2020-04-01
作者简介 About authors
刘敏,E-mail:liumin272@163.com。

摘要
关键词:
Abstract
Keywords:
PDF (5774KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
刘敏, 房玉林. 高温胁迫对葡萄幼树生理指标和超显微结构的影响[J]. 中国农业科学, 2020, 53(7): 1444-1458 doi:10.3864/j.issn.0578-1752.2020.07.013
LIU Min, FANG Yulin.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】随着全球气候变暖和夏季极端高温天气频发,高温胁迫已经成为制约农业生产的主要逆境之一。葡萄是世界上广泛种植且具有较高经济价值的果树,在高温胁迫下葡萄植株生长受到抑制,叶片发生卷曲干枯,果实发生日灼现象,严重影响果实的品质和产量[1]。因此,研究葡萄高温响应机制对于提高葡萄植株耐热性,保证夏季植株生长和果实发育具有重要意义。【前人研究进展】在高温胁迫下,植物体内会产生大量对细胞有害的活性氧(Reactive oxygen species,ROS),比如超氧阴离子($O_{2^{\frac{}{.}}}$)、过氧化氢(H2O2)、羟自由基(·OH)等,从而引起氧化胁迫[2]。ROS的增加对细胞结构和功能造成很大伤害,可以使叶绿体和线粒体发生膨胀,蛋白和核酸等大分子变性,还能引起DNA断裂[3]。植物在长期的进化过程中形成了完善而复杂的抗氧化系统来清除ROS,以降低其对细胞的伤害。常见的抗氧化酶包括过氧化氢酶(CAT)、过氧化物酶(POD)、超氧化物歧化酶(SOD)、抗坏血酸过氧化物酶(APX)等。高温胁迫对葡萄叶片中抗氧化酶活性的影响不尽相同。张俊环等[4]对‘京秀’葡萄幼苗进行45℃高温处理,发现叶片中CAT、POD和SOD 3种酶的活性有不同程度的降低,其中SOD活性的降低幅度最大。查倩等[5]对‘夏黑’葡萄幼苗进行45℃高温处理,在处理6 h后SOD活性显著降低,在处理150 h后恢复至正常水平;而POD和CAT活性在处理6 h和150 h后均显著高于对照。一般认为,适度的高温胁迫会增加葡萄叶片中抗氧化酶的活性,当温度超过一定范围时,抗氧化酶的蛋白结构受到破坏,活性显著降低。高温对植物体中抗氧化酶活性的影响因品种、器官、发育阶段、处理时间、处理温度、抗氧化酶种类的不同而有所差异[6]。光合作用是植物物质转化和能量代谢的关键,其对于高温非常敏感,这是由于光系统II(PS II)的活性容易受到高温的抑制[7,8,9]。高温胁迫会导致植物吸收的光能超过CO2同化所需的量,引起植物体内ROS的积累,造成PSII的损伤[10,11,12,13]。有研究表明,葡萄在受到高温胁迫后,供体侧放氧复合体比受体侧的电子传递对高温更加敏感[14,15]。【本研究切入点】刺葡萄是我国特有的野生葡萄种质资源,离体叶片的高温处理结果显示,刺葡萄的耐热性显著高于其他野生品种和栽培品种[16],但是刺葡萄盆栽苗在控制条件下的高温响应机制尚不明确。【拟解决的关键问题】本研究以欧亚种‘赤霞珠’和刺葡萄‘君子1号’扦插苗为试验材料,进行不同温度的高温处理,通过抗氧化酶、光合参数等指标的测定和扫描电镜、透射电镜的观察,比较两个葡萄品种的耐热性,研究葡萄在高温胁迫下的生理响应机制,为耐热品种的选育和耐热机理的研究提供参考。1 材料与方法
试验于2018年3—6月在西北农林科技大学葡萄酒学院温室进行。1.1 试验材料
将一年生欧亚种‘赤霞珠’(Vitis vinifera L. ‘Cabernet Sauvignon’)和刺葡萄‘君子1号’(Vitis davidii Foex. ‘Junzi 1’)扦插苗种植在塑料盆(直径14 cm,高度10 cm)中,营养土与珍珠岩比例为4﹕1,培养温度为22—27℃,湿度为70%—90%,每周浇2次水。移栽3个月后,植株长至10—12片叶,选择健康无病虫害、长势一致的植株进行试验处理。1.2 处理方法
将‘赤霞珠’和‘君子1号’植株移至25℃植物培养箱中适应2 d,湿度70%,光照12 000 lx,第3天将植株分别置于35℃、40℃、45℃植物培养箱中连续处理48 h,昼夜16 h/8 h,以25℃培养的植株作为对照,每个处理5盆。1.3 生理指标的测定方法
1.3.1 相对电导率 采集葡萄植株从基部数第4—6片叶,用打孔器取15个直径1 cm叶圆片,分别置于3个装有10 mL超纯水的离心管中,每管5个,在150 r/min、25℃的摇床上振荡3 h,用DDS-307型电导率仪测定初始电导率(E1),然后将装有样品的离心管置于水中煮沸30 min,冷却至室温测定总电导率(E2),计算相对电导率(%)=E1/E2×100。1.3.2 相对含水量 采集葡萄植株从基部数第4—6片叶立即称重,记为鲜重(FW),再将叶片浸入蒸馏水中4 h,取出擦干称重,记为饱和鲜重(TW),然后将叶片置于80℃烘箱中烘至恒重,记为干重(DW),叶片相对含水量(%)=(FW-DW)/(TW-DW)×100。
1.3.3 叶绿素总量 采集葡萄植株从基部数第4—6片叶,用剪刀将叶片剪成细丝,置于25 mL的容量瓶中,记录叶片重量;加入0.5 mL纯丙酮和15 mL 80%丙酮,室温下避光浸提过夜;用80%的丙酮定容至25 mL,过滤后用UV-1800型紫外-可见分光光度计分别测定在663 nm和645 nm处的吸光值,计算总叶绿素含量(mg∙g-1 FW)。
1.3.4 抗氧化酶活性和MDA含量 采集葡萄植株从基部数第4—6片叶,在液氮保护下磨成粉末,装在2 mL离心管中,每管约0.1 g,在离心管中加入1 mL提取液,摇匀后于8 000×g 4℃离心15 min,取上清液,置于冰上待测。参照超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)、丙二醛(MDA)试剂盒(苏州科铭)的操作说明进行加样和反应,用UV-1800型紫外-可见分光光度计测定吸光值,并计算酶活性和MDA含量,每个处理3次重复。
1.3.5 叶绿素荧光参数 用便携式调制叶绿素荧光仪PAM2500(Walz,德国)测定叶绿素荧光参数。每个处理选取从基部数第4—6片叶进行测定,测定前将叶片用叶夹夹住,暗适应30 min,然后测定初始荧光(Fo);用饱和脉冲光(10 000 μmol·m-2·s-1)照射0.5 s,测定最大荧光(Fm);用500 μmol·m-2·s-1的活化光照射叶片,当Ft稳定后,再次打开饱和脉冲光,测定Fm′;关闭活化光同时打开远红光照射5 s,测定Fo′;记录Fv/Fm、ΦPSⅡ、qP、Fv′/Fm′等参数。
1.3.6 光合参数 每个处理选取从基部数第4—6片叶,在上午9:00—11:00用LI-6800便携式光合仪(LI-COR,美国)测定净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2(Ci)和气孔导度(Gs)。设定参数为:流速500 μmol·s-1,相对湿度70%,CO2浓度400 μmol·mol-1,风扇速度10 000 r/min,温度25℃,光强1 800 μmol·m-2·s-1。
1.4 组织化学染色法
1.4.1 超氧阴离子染色将葡萄叶片浸入到NBT溶液(1 mg·L-1,用pH 7.8的PBS配置)中,抽真空10 min,室温下放置30 min,叶片出现蓝色斑点,将其置于在脱色液(乙醇﹕乙酸﹕甘油=3﹕1﹕1)中,90℃水浴加热,直至绿色完全脱去,最后将叶片置于蒸馏水中准备拍照。
1.4.2 过氧化氢染色 将葡萄叶片浸入到DAB溶液(1 mg·L-1,pH=3.8—4.0)中,抽真空10 min,在摇床上80 r/min避光振荡4 h,叶片出现棕色斑点,脱色方法同上。
1.5 扫描电镜观察
45℃高温处理前及处理24 h后,采集植株中部的功能叶片,取主叶脉两侧的部分,用刀片切成5 mm×5 mm的小方块,将其放在2 mL离心管中,用戊二醛固定。用PBS缓冲液清洗4次后,用乙醇梯度脱水,然后用乙酸异戊酯置换。将样品进行干燥、喷金,在S4800场发射扫描电子显微镜(日立,日本)下进行观察和拍照。用300倍照片计算气孔密度,用5 000倍照片测量气孔大小和开张程度,每个处理选取10个视野,取平均值,用Photoshop软件进行测量。1.6 透射电镜观察
45℃高温处理前及处理24 h后,采集植株中部的功能叶片,取叶片主叶脉两侧的部分,用刀片切成2 mm×5 mm小方块,将其放在2 mL离心管中,用戊二醛固定。用PBS缓冲液冲洗样品,锇酸固定,再用PBS缓冲液清洗,乙醇梯度脱水,用白胶进行渗透和包埋。用超薄切片机EM UC7(徕卡,德国)切片,将样品用醋酸双氧铀和柠檬酸铅染色。用HT7700透射电子显微镜(日立,日本)进行观察和拍照。统计叶绿体和淀粉粒的数量,用Photoshop软件测量其长度和宽度,每个处理选取10个视野,取平均值。1.7 数据处理
采用SPSS 21软件对试验数据进行方差分析。2 结果
2.1 高温对葡萄植株形态的影响
‘赤霞珠’和‘君子1号’在35℃和40℃处理48 h后,植株形态没有明显变化,新梢正常生长,叶片边缘完整,未出现热害症状(图1)。而在45℃处理后,‘赤霞珠’顶部的2片幼叶呈烧焦状,其他叶片边缘有不同程度的卷曲、干枯,叶柄夹角变大,但叶柄颜色正常,未出现坏死症状。‘君子1号’在45℃处理后,顶部3片叶完全萎蔫,其余叶片在边缘和中部均出现坏死斑,值得注意的是‘君子1号’的叶柄在距离叶柄洼约1 cm处的细胞坏死,阻断了水分向叶片的运输,加重了叶片的胁迫症状。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1高温处理对葡萄植株形态的影响
A:赤霞珠;B:君子1号
Fig. 1Effects of heat treatment on grapevines morphology
A: Cabernet Sauvignon; B: Junzi 1
‘赤霞珠’叶片为5裂,表面平滑;在45℃处理24 h后,叶片从边缘开始脱水、卷曲、变黄;在处理36 h后,这种现象逐渐向叶片中部扩展;在处理48 h后,叶片90%面积都失去绿色,而叶柄未见明显的失水症状(图2—A),可见‘赤霞珠’叶片受热最敏感的部位是叶片边缘。‘君子1号’叶片呈心形,无裂刻,边缘锯齿形,表面平滑有光泽,颜色比‘赤霞珠’深,叶柄有皮刺;在45℃处理24 h后,叶柄从靠近叶片的一端开始出现损伤;在处理36 h后,整个叶柄细胞都脱水死亡,而叶片没有出现明显的损伤迹象;在处理48 h后,与叶柄相连的叶脉干枯变褐,叶片上出现大面积的坏死斑(图2—B)。对‘君子1号’进行多次高温试验均发现此现象,可见其叶柄表皮细胞对高温比较敏感,可能是为了保存茎中的水分,具体原因需进一步研究。
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2葡萄叶片在高温处理过程中的变化
A:赤霞珠;B:君子1号
Fig. 2Changes of grape leaves during heat treatment
A: Cabernet Sauvignon; B: Junzi 1
2.2 高温对电导率、含水量和叶绿素总量的影响
高温胁迫后,植物细胞膜结构受到不同程度的损伤,导致细胞膜透性增大,细胞内电解质外渗,电导率增大,还会引起叶片含水量的降低和叶绿素的降解。在35℃和40℃处理48 h后,‘赤霞珠’和‘君子1号’叶片的相对电导率、相对含水量和叶绿素总量与对照(25℃)相比均没有显著差异;在45℃处理后,二者的相对电导率均显著升高,而相对含水量和叶绿素总量显著降低(图3)。常温下,‘赤霞珠’和‘君子1号’叶片的相对电导率约为20%,在45℃处理后,二者相对电导率分别升至22.43%和34.18%,说明‘君子1号’的细胞膜损伤程度比‘赤霞珠’更严重。‘赤霞珠’和‘君子1号’叶片的相对含水量在25℃下分别为88.34%和89.27%,在45℃处理后分别降至对照的75.94%和67.09%,‘君子1号’细胞失水更为严重。高温胁迫会抑制叶绿素合成相关酶的活性,从而降低叶绿素含量。在25℃、35℃、40℃条件下,‘赤霞珠’和‘君子1号’叶片中叶绿素总量均大于2.5 mg·g-1 FW,而45℃处理后二者叶绿素总量分别为1.61和1.48 mg·g-1 FW,与对照相比分别降低了41.67%和48.07%,‘君子1号’叶片中叶绿素总量的降低幅度更大。图3
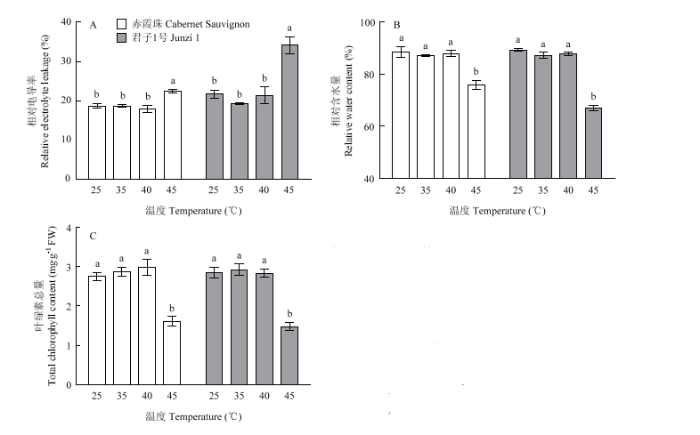 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3高温处理对葡萄叶片相对电导率、相对含水量和叶绿素总量的影响
不同小写字母表示差异显著,P<0.05。下同
Fig. 3Effects of heat treatment on relative electrolyte leakage, relative water content and total chlorophyll content of grape leaves
Different small letters mean significant differences (P<0.05). The same as below
2.3 高温对抗氧化酶活和MDA含量的影响
‘赤霞珠’叶片中SOD酶活性随着处理温度的升高而增加,40℃处理后,‘君子1号’叶片中SOD酶活显著高于35℃处理,但与45℃处理无显著差异。‘赤霞珠’叶片中POD酶活性的大小顺序为40℃>35℃>25℃,45℃处理后POD酶活与40℃处理无显著差异;‘君子1号’叶片中POD酶活随温度升高显著增加,且相同温度处理后‘君子1号’POD酶活大于‘赤霞珠’。‘赤霞珠’叶片中CAT酶在40℃处理后活性最高,其次为45℃和35℃处理,高温处理后CAT酶活显著高于对照;‘君子1号’叶片中CAT酶在35℃处理后活性最高,其次是40℃处理,45℃处理后CAT酶活与对照无显著差异。随着处理温度的升高,叶片中MDA含量显著上升,且高温处理后,‘君子1号’叶片中MDA含量高于‘赤霞珠’(图4)。图4
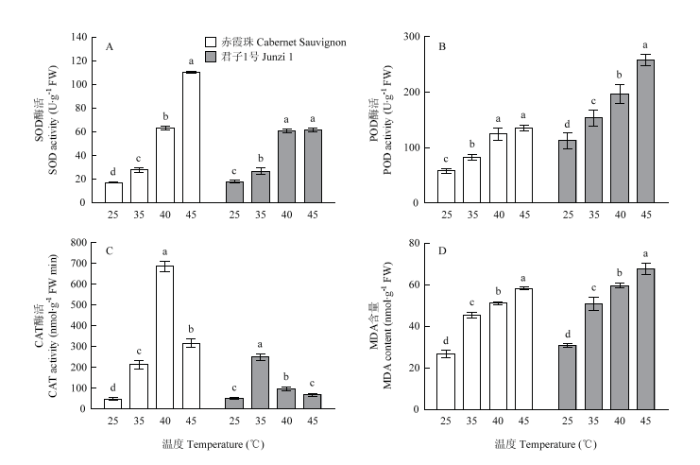 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4高温处理对葡萄叶片中抗氧化酶活性和MDA含量的影响
Fig. 4Effects of heat treatment on antioxidases activities and MDA content in grape leaves
2.4 高温处理对$O_{2^{\frac{}{.}}}$和H2O2含量的影响
随着处理温度的升高,‘赤霞珠’和‘君子1号’叶片上的蓝色斑点增多,说明$O_{2^{\frac{}{.}}}$含量有所增加(图5)。在35℃处理后叶片上几乎看不到蓝色斑点;在40℃处理后,‘赤霞珠’叶片边缘分布有蓝色斑点,而‘君子1号’叶片上的斑点零星分布;在45℃处理后,‘赤霞珠’整个叶片布满蓝色斑点,而‘君子1号’叶片上的斑点数量明显少于‘赤霞珠’。结合植株形态,‘君子1号’受胁迫的程度高于‘赤霞珠’,但在45℃处理后,‘君子1号’叶片中$O_{2^{\frac{}{.}}}$含量却低于‘赤霞珠’,有可能是在处理48 h时测定,已经错过了$O_{2^{\frac{}{.}}}$含量高峰。‘赤霞珠’和‘君子1号’叶片中H2O2含量随处理温度的升高而增加,45℃处理后,‘君子1号’叶片上布满大量棕色斑点(图6),说明在叶片中积累了大量H2O2。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5高温处理48 h后叶片$O_{2^{\frac{}{.}}}$染色
Fig. 5$O_{2^{\frac{}{.}}}$ staining of leaves after heat treatments for 48 h
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6高温处理48 h后叶片H2O2染色
Fig. 6H2O2 staining of leaves after heat treatments for 48 h
2.5 高温对叶绿素荧光参数的影响
光合作用对温度比较敏感,高温会破坏叶片PS II,通过检测叶绿素荧光可以了解植物的光合状况,其参数变化可以用来评价植株的抗逆性。在35℃和40℃处理后,‘赤霞珠’和‘君子1号’叶片的Fo、Fm、Fv/Fm和ΦPS II与对照相比均无显著差异,只有在45℃处理后各参数受到不同程度的影响(图7)。Fo表示最小荧光,是PSⅡ反应中心完全开放时的荧光产量。45℃处理后,‘赤霞珠’叶片Fo值没有显著变化,而‘君子1号’叶片Fo值为465.67,显著高于对照(311.67),Fo值上升的主要原因是PS II反应中心的失活。Fm表示最大荧光,是PS II反应中心完全关闭时的荧光产量。45℃处理后,‘赤霞珠’叶片Fm值没有显著变化,而‘君子1号’叶片Fm值为1 159.33,显著低于对照(1 535.33)。Fv/Fm表示暗适应下PS II最大光化学效率,反映PS II反应中心最大光能转换效率。常温下Fv/Fm值为0.8左右,当植物受到胁迫时Fv/Fm会降低。在25℃时,‘赤霞珠’和‘君子1号’Fv/Fm分别为0.780和0.798,45℃处理后,二者Fv/Fm显著下降,分别降至0.733和0.598,‘君子1号’降低幅度更大。ΦPS II表示在光照条件下PS II反应中心部分关闭后的实际光化学效率,反映叶片在光下用于电子传递的能量占吸收光能的比例。45℃处理后,‘赤霞珠’和‘君子1号’叶片ΦPS II比对照分别降低了56.41%和74.42%,说明高温抑制了植物的实际光化学效率。图7
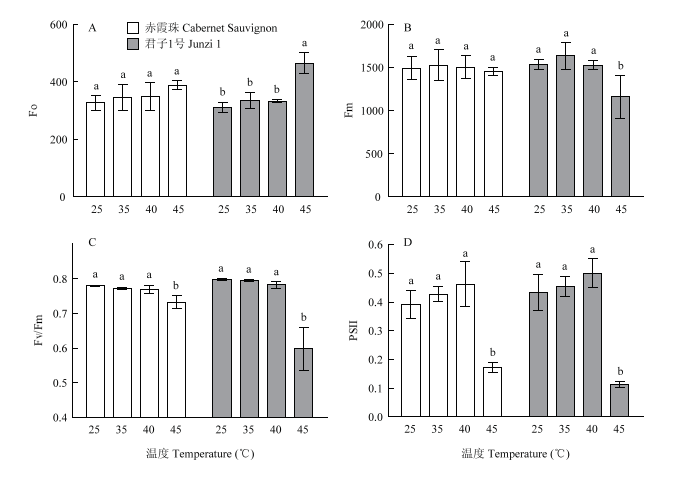 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7高温处理对葡萄叶绿素荧光参数的影响
Fig. 7Effects of heat treatment on chlorophyll fluorescence parameters of grapevines
2.6 高温对光合参数的影响
在高温处理48 h后,‘赤霞珠’和‘君子1号’叶片净光合速率(Pn)、蒸腾速率(Tr)和气孔导度(Gs)变化趋势相同,随着温度升高而降低(图8)。在35℃处理后,‘赤霞珠’叶片胞间CO2(Ci)没有显著变化,而‘君子1号’Ci显著降低;在40℃处理后,二者Ci值均显著降低,而在45℃处理后,Ci值又有所回升。说明35℃和40℃处理后,Pn降低是由气孔因素导致,而45℃处理后的Pn降低是由非气孔因素导致。图8
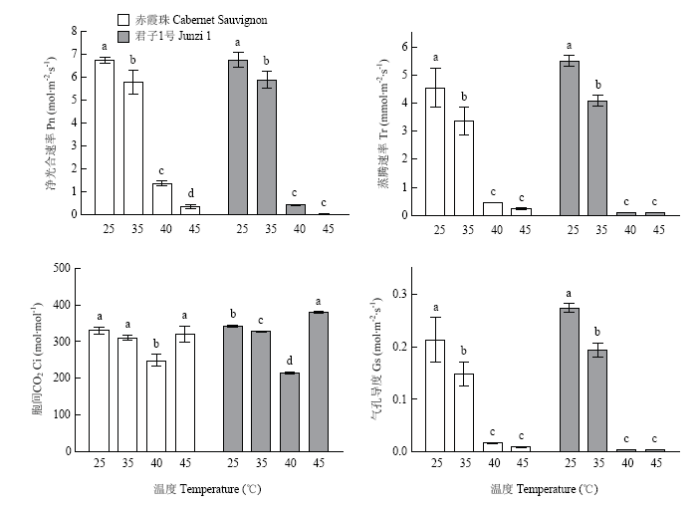 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8高温处理对葡萄光合参数的影响
Fig. 8Effects of heat treatment on photosynthetic parameters of grapevines
2.7 高温对葡萄叶片气孔特性的影响
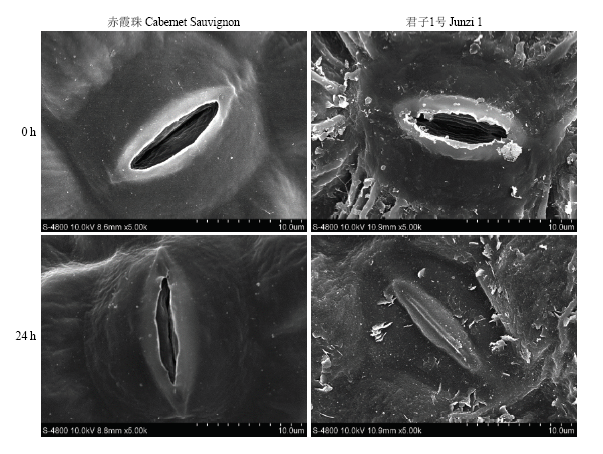
由于45℃高温48 h处理后,细胞结构破坏严重,所以取处理24 h的叶片进行扫描电镜和透射电镜的观察。扫描电镜结果显示,在45℃高温处理24 h后,‘赤霞珠’和‘君子1号’的气孔开张度明显减小(图9),这样可以缓解高温导致的水分散失。图9
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图945℃高温处理对葡萄叶片气孔形态的影响
Fig. 9Effects of heat treatment at 45℃ on the stomatal morphology of grape leaves
常温下,‘赤霞珠’和‘君子1号’气孔开张度分别为3.44 μm和1.97 μm,在45℃高温处理24 h后,二者气孔开张度显著降低,分别降低了38.66%和38.07%(表1)。高温对‘赤霞珠’气孔长度和宽度无显著影响,而‘君子1号’气孔长度和宽度在高温处理24 h后有所降低,但差异不显著。气孔密度不受高温处理的影响,‘赤霞珠’和‘君子1号’的气孔密度范围分别是177.60—187.35个/mm2和122.95—132.71个/mm2。由表1可知,常温下‘赤霞珠’的气孔大小、气孔开张度和气孔密度均大于‘君子1号’。
Table 1
表1
表145℃高温处理对葡萄叶片气孔特性的影响
Table 1
| 品种 Variety | 处理时间 Time after treatment (h) | 气孔开张度 Degree of stomatal opening (μm) | 气孔长度 Stomatal length (μm) | 气孔宽度 Stomatal width (μm) | 气孔密度 Stomatal density (No./mm2) |
|---|---|---|---|---|---|
| 赤霞珠 Cabernet Sauvignon | 0 | 3.44±0.97a | 16.59±1.32a | 7.23±1.46a | 187.35±20.65a |
| 24 | 2.11±0.78b | 16.52±1.54a | 6.33±1.03ab | 177.60±17.31a | |
| 君子1号Junzi 1 | 0 | 1.97±0.54b | 15.51±2.06ab | 5.82±0.75ab | 132.71±7.81b |
| 24 | 1.22±0.77c | 13.98±1.42b | 5.48±1.10b | 122.95±17.31b |
新窗口打开|下载CSV
2.8 高温对葡萄叶绿体结构的影响
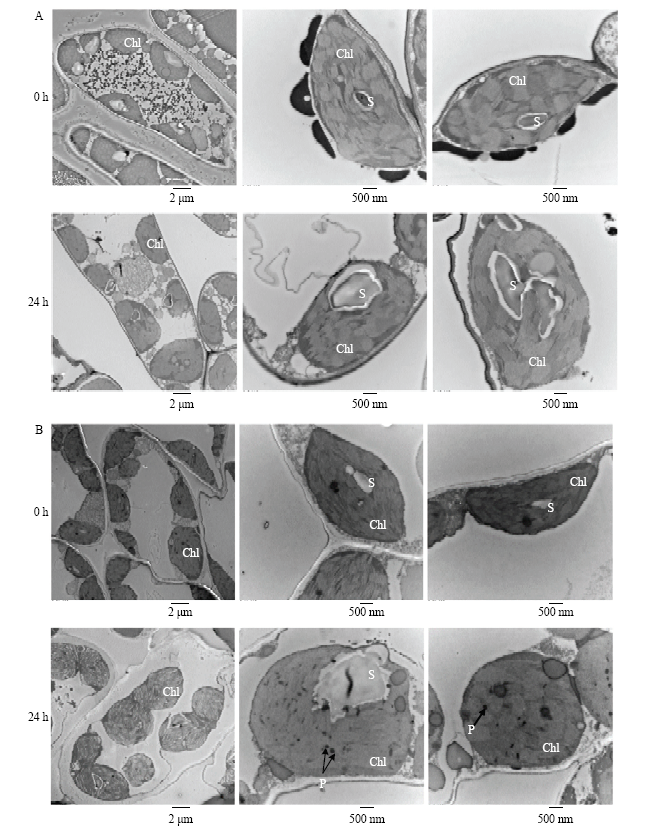
常温下,葡萄叶绿体呈梭形,紧贴细胞膜生长,叶绿体膜完整,基粒片层清晰可见,表面附着少量淀粉粒和嗜锇颗粒(图10)。高温处理后,葡萄叶绿体体积变大,形状变圆,叶绿体膜出现解体,基粒片层断裂甚至解体,出现大量巨型淀粉粒,而且由于液泡膜的破坏,叶绿体向细胞内部移动;同时在‘君子1号’叶绿体上观察到大量嗜锇颗粒,可加速叶绿体老化。由此可见,高温下‘君子1号’叶绿体结构变化较明显。图10
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1045℃高温处理对葡萄叶片中叶绿体超微结构的影响
A:赤霞珠;B:君子1号。Chl:叶绿体;S:淀粉粒;P:嗜锇颗粒
Fig. 10Effects of heat treatment at 45℃ on the chloroplast ultrastructure of grape leaves
A: Cabernet Sauvignon; B: Junzi 1. Chl: Chloroplast; S: Starch grain; P: Plastoglobule
在45℃高温处理后,‘赤霞珠’和‘君子1号’的叶绿体数目和长度与对照相比无显著差异,但叶绿体的宽度显著增加,分别增加了29.04%和75.61%(表2),说明高温使叶绿体体积变大,形状变圆(图10)。高温处理后,叶绿体表面的淀粉粒数目显著增加,而且淀粉粒的体积也随之增大。‘赤霞珠’的淀粉粒长度平均增加均0.62 μm,宽度平均增加约0.38 μm,而‘君子1号’淀粉粒体积变化更大,其长度和宽度分别平均增加约1.26 μm和0.94 μm(表2)。由此可知,高温下‘赤霞珠’叶绿体结构的稳定性高于‘君子1号’。
Table 2
表2
表245℃高温处理对葡萄叶片中叶绿体和淀粉粒的影响
Table 2
| 品种 Variety | 时间 Time (h) | 叶绿体Chloroplast | 淀粉粒Starch grain | ||||
|---|---|---|---|---|---|---|---|
| 数目/细胞 (No./cell) | 长度 Length (μm) | 宽度 Width (μm) | 数目/叶绿体 (No./chloroplast) | 长度 Length (μm) | 宽度 Width (μm) | ||
| 赤霞珠 Cabernet Sauvignon | 0 h | 6.4±1.9a | 5.83±0.63a | 3.03±0.24b | 0.6±0.5b | 1.21±0.25b | 0.68±0.14c |
| 24 h | 5.0±2.2a | 5.42±0.98a | 3.91±0.76a | 1.1±0.8a | 1.83±0.54a | 1.06±0.21b | |
| 君子1号 Junzi 1 | 0 h | 6.3±1.4a | 5.41±0.96a | 2.46±0.37c | 0.7±0.6b | 0.87±0.09b | 0.42±0.05d |
| 24 h | 5.7±0.8a | 5.39±0.75a | 4.32±0.48a | 1.2±0.9a | 2.13±0.62a | 1.36±0.40a | |
新窗口打开|下载CSV
3 讨论
3.1 高温影响葡萄抗氧化酶活性
高温胁迫会诱导植物体产生ROS,为避免ROS对生物膜及其他大分子物质的伤害,植物体内抗氧化酶系统发挥积极的作用。SOD、POD和CAT是植物体内重要的抗氧化酶,SOD可以将$O_{2^{\frac{}{.}}}$转化为H2O2和O2,而POD和CAT可以进一步将H2O2分解为H2O和O2[3]。这3种酶可在一定程度上清除体内过剩的ROS,维持ROS代谢平衡,保护膜结构,使植物具有抵抗逆境胁迫的能力。但是当胁迫超过植物的承受极限时,SOD、POD和CAT活性会下降或结构受到破坏,膜脂过氧化程度加剧,细胞的正常代谢和植物生长受到抑制。在本研究中,高温处理后,葡萄叶片中SOD、POD和CAT酶活均高于对照,SOD和POD在45℃处理后活性最高,而CAT在35℃或40℃处理后活性最高。ZOU等[17]对耐热性不同的两种油菜进行高温处理,结果显示中度高温胁迫(30℃和35℃)可以激活SOD和POD,但重度高温胁迫(40℃)会抑制这两种酶的活性,而35℃处理后CAT酶活显著低于对照,说明与SOD和POD相比,CAT酶对高温更敏感,与本研究结果一致。油菜耐热品种WS-1的3种酶活性在不同温度下均高于不耐热品种WS-6,而在本研究中,‘赤霞珠’(耐热)SOD和CAT酶活高于‘君子1号’(不耐热),而且与POD酶活性相比,‘赤霞珠’SOD和CAT酶活的增加幅度较大,说明SOD和CAT在葡萄高温响应过程中发挥重要作用。田治国等[18]研究发现高温胁迫提高了孔雀草和万寿菊SOD、POD和CAT的活性,而且耐热品种3种酶活性的增幅高于不耐热品种,SOD和POD变化幅度大于CAT,可见孔雀草和万寿菊在高温胁迫下SOD和POD在清除ROS方面起主要作用。ZHOU等[19]对两个番茄品种进行高温处理,结果显示在处理第3天,两个品种的SOD和APX活性显著升高,而CAT活性显著降低。由此可见,高温胁迫下植物抗氧化酶的活性受到物种、品种、处理时间、处理温度等因素的影响。在实际生产中,人们可以对植物进行预处理以提高抗氧化酶活性,从而提高植株对高温的抗性。孙军利等[20]以二年生‘克瑞森’无核葡萄为材料,研究外源水杨酸对高温胁迫下抗氧化酶和抗氧化剂含量的影响,结果表明外源水杨酸促进了高温胁迫下葡萄叶片中抗坏血酸和谷胱甘肽的积累,并维持较高的抗氧化酶活性,包括谷胱甘肽还原酶(GR)、APX、SOD、POD和CAT,从而降低高温胁迫下ROS对植株的伤害。H2O2预处理也可以显著提高高温下黄瓜叶片的抗氧化酶活性,包括脱氢抗坏血酸还原酶(DHAR)、单脱氢抗坏血还原酶(MDHAR)、谷胱甘肽过氧化物酶(GSH)、GR、SOD和APX,从而降低膜脂过氧化程度,保护叶绿体结构免受高温破坏[21]。HUANG等[22]研究发现在营养液中添加30 mmol·L-1葡萄糖,高温下黄瓜叶片抗氧化酶的活性显著提高,同时Cu/Zn-SOD、Mn-SOD、CAT、GR等基因表达量显著增加,从而抑制了ROS的积累,缓解了高温对植株生长的抑制作用。
3.2 高温影响葡萄光合作用
本研究发现,35℃和40℃对‘赤霞珠’和‘君子1号’的植株形态、叶片相对电导率、相对含水量、叶绿素总量和叶绿素荧光参数均没有显著影响,但是光合参数却随着处理温度的升高而降低。Fo值可以反映PS II反应中心的损伤程度,Fo值越高,说明损伤程度越大。在35℃和40℃处理48 h后,Fo值与对照无显著差异,说明PS II反应中心并没有受到高温损伤,但是Pn值却显著低于对照,推测由气孔因素导致,而Ci值显著降低也证明了这一点。在45℃处理后,‘赤霞珠’和‘君子1号’的植株形态、叶片相对电导率、相对含水量、叶绿素总量、叶绿素荧光参数和光合参数均发生了显著变化,Fo值显著上升说明了PS II反应中心失活,直接导致Pn值的下降,同时Ci值上升,说明45℃下光合速率的降低由非气孔因素导致。罗海波等[14]对‘赤霞珠’在每日9:30—15:30进行40℃处理,其余时间为30℃,研究发现葡萄叶片光合速率显著下降,在常温下1 d可以恢复,作者认为40℃下Pn下降不是气孔因素导致,主要与PS II供体侧和反应中心活性以及激发能的分配有关。这个结果与本试验结果不一致,可能与处理方式、光照强度等因素有关。查倩等[5]对‘夏黑’进行35℃和45℃高温处理,处理时间为10:00—16:00,结果显示在处理6 h后,Fv/Fm值在35℃下无显著变化,而在45℃下显著降低,与本试验结果一致。值得注意的是,在45℃处理150 h后,Fv/Fm又恢复至原初水平,推测‘夏黑’具有较强的高温适应能力。韩晓等[23]评价8种砧木对‘87-1’葡萄光合特性和叶绿素荧光特性的影响,结果表明‘87-1’/‘SO4’、‘87-1’/‘华葡1’号在不同温度下净光合速率变化较小,在高温下净光合速率较大,说明SO4和华葡1号两种砧木可以提高‘87-1’的耐热性。本研究使用的两个葡萄品种为欧亚种,‘赤霞珠’是世界上广泛种植的酿酒葡萄品种,原产自法国波尔多,能适应各种不同的气候条件,在我国新疆等炎热产区具有良好的表现;刺葡萄是我国的野生葡萄品种,广泛分布在我国南方地区,生于山坡、沟谷疏林或灌丛中,具有较强抗病能力。研究结果表明,‘赤霞珠’的耐热性显著高于‘君子1号’,可能由于刺葡萄比较适合高温高湿的环境,能耐受的绝对温度没有‘赤霞珠’高。徐洪国[16]将葡萄离体叶片在47℃下处理40 min后测定Fv/Fm值,用以区分不同品种的耐热性,结果显示刺葡萄和‘赤霞珠’的Fv/Fm值分别为0.68和0.63,刺葡萄的耐热性应该高于‘赤霞珠’,与本研究结果不一致。这可能由于试验方法不同,或者是由于刺葡萄分为很多品系,不同品系的耐热性有所不同。在今后的试验中可以设计不同湿度的高温处理,对欧亚种和刺葡萄的高温响应进行比较和研究。
另外,本试验用调制式叶绿素荧光仪只能了解PS II的总体特征,而不能了解PS II各个部分的具体变化情况,包括供体侧、反应中心和受体侧。在下一步试验中,可以用植物效率分析仪测定快速叶绿素荧光诱导动力学曲线(OJIP),以获得更多的PS II光化学信息,进一步探讨葡萄在高温胁迫下光合作用的变化机理。
3.3 高温影响葡萄气孔特性
气孔是植物叶片上的孔状结构,是植物进行气体交换的重要器官。植物叶片通过改变气孔开张度、大小、密度和空间分布格局等特性来优化气体交换效率。本研究中,45℃高温处理后‘赤霞珠’和‘君子1号’叶片的气孔密度和气孔大小无显著变化,而气孔开张度显著降低。在高温条件下,植物通过减小气孔开张度或者关闭部分气孔来减少叶片的水分散失。高温处理后,北高丛越橘‘布里吉塔’和‘蓝丰’的气孔长度、宽度和密度均显著降低[24],与本研究结果不一致,可能与品种、温度、处理方式等因素有关。有研究者认为耐热品种比不耐热品种气孔密度高,使蒸腾作用加强,有效降低叶片表面的温度,使叶片组织免受高温伤害[25]。本研究发现‘赤霞珠’叶片的气孔密度显著大于‘君子1号’,这可能是‘赤霞珠’耐热性高的原因之一。3.4 高温影响葡萄叶绿体结构
叶绿体对高温胁迫比较敏感,在高温条件下叶绿体会产生ROS,如果不能及时清除,会导致叶绿体膜及类囊体膜的解体[26]。本研究中2个葡萄品种的叶绿体在高温胁迫后受到不同程度的损伤,表现为叶绿体膨胀、基粒片层断裂、叶绿体膜解体、淀粉粒和嗜锇颗粒增多、变大。黄瓜在受到高温胁迫后叶绿体会发生肿胀,异常叶绿体占总数的71.4%,而H2O2预处理可以显著降低高温导致的异常叶绿体的比例(33.3%)[21]。北高丛越橘受到高温胁迫后,叶绿体肿胀为近圆形,叶绿体内出现较多的淀粉粒和嗜锇颗粒[27],与本研究结果一致。‘赤霞珠’叶片受到盐胁迫后,叶绿体也会发生类似变化[28],但产生这一现象的原因尚不明确。光合产物主要以蔗糖的形式通过韧皮部向外运输,也有一部分同化产物以淀粉的形式在体内积累。在高温处理后植株韧皮部胼胝体增多,堵塞维管束,影响同化产物在韧皮部的运输,从而导致淀粉粒的积累[29]。在常温下,叶绿体内嗜锇颗粒处于动态平衡中;在高温条件下,叶绿体或类囊体的膜脂被ROS氧化从而形成嗜锇颗粒,所以嗜锇颗粒的大小和数量可以反映叶绿体膜脂过氧化的程度[30]。在本研究中,在高温处理后‘君子1号’叶绿体表面出现大量嗜锇颗粒,说明其叶绿体膜脂过氧化程度大于‘赤霞珠’。
叶绿体是光合作用的场所,高温会导致叶绿体超微结构的变化和光合机构的破坏,从而影响植株的光合作用。在本研究中,‘君子1号’叶绿体结构在高温下的稳定性较差,这也许是其光合速率降低的主要原因之一。植物的耐热性是多基因控制的数量性状,具体反映在植物的组织解剖结构、气孔特性、细胞器变化、膜脂过氧化程度、抗氧化酶等方面。要准确评价植物的耐热性,需要综合考虑形态结构、生理生化、遗传等因素,并参考植株田间受害及受害后恢复状况,最终得出结论。
4 结论
通过抗氧化酶、光合参数等指标的测定和植株形态、气孔形态、叶绿体结构的观察,确定‘赤霞珠’的耐热性显著大于‘君子1号’。在中度高温胁迫下(35℃和40℃),‘赤霞珠’和‘君子1号’的PSII活性中心未发生不可逆损伤,净光合速率(Pn)降低由气孔因素导致;而在重度高温胁迫下(45℃),‘君子1号’PSII活性中心的损伤程度大于‘赤霞珠’,Pn降低由非气孔因素导致。‘赤霞珠’的气孔密度大,叶绿体结构稳定,高温处理后超氧化物歧化酶和过氧化氢酶活性大幅增加是其耐热性高的重要原因。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 2]
[D].
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}