Abstract Grain yield per ear of maize (Zea mays L.) is composed of both kernel number and grain weight. The number of kernels on an ear is determined by not only the number of kernel rows which is closely related to the inflorescence development, but also the number of fertile florets generated by the flower meristem. Therefore, those genes for inflorescence architecture and flower development are potentially involved in the genetic control of kernel number. Maize kernel is a single-seeded fruit comprised of the maternally derived pericarp, and embryo and endosperm derived from double fertilization. Both embryo and endosperm account for the vast majority of the mature kernel mass, and directly determine the kernel size and weight. In this paper, we outlined the genetic controls of kernel number with the emphasis on the inflorescence and floret related genes that are involved in the CLAVATA- WUSCHEL (CLV-WUS) feedback loop, hormone biosynthesis and signaling, floral organ development and sex determination. In particular, we described the regulatory network models for interplays among phytohormones including auxin, gibberellin, cytokinin and strigolactone in the inflorescence architecture and floral organ development. We also summarized those embryo and endosperm developmental genes involving in processing and editing of mitochondrial transcripts, transcription and translation of some chloroplast DNAs as well as nuclear RNAs. Most of these genes encode PPR proteins targeted to mitochondria or plastids. Recently, several studies have identified a new pathway to control kernel development by regulating the transcription and processing of pre-mRNA within the nucleus. Here, we also discussed the association between these genes and kernel number or kernel weight, and the potential areas of research for deciphering molecular mechanisms of grain yield in maize. Keywords:Zea mays L.;kernel number per ear;kernel weight;inflorescence;floret;embryo;endosperm
PDF (984KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 赵然, 蔡曼君, 杜艳芳, 张祖新. 玉米籽粒形成的分子生物学基础[J]. 中国农业科学, 2019, 52(20): 3495-3506 doi:10.3864/j.issn.0578-1752.2019.20.001 ZHAO Ran, CAI ManJun, DU YanFang, ZHANG ZuXin. Molecular Basis of Kernel Development and Kernel Number in Maize (Zea mays L.)[J]. Scientia Acricultura Sinica, 2019, 52(20): 3495-3506 doi:10.3864/j.issn.0578-1752.2019.20.001
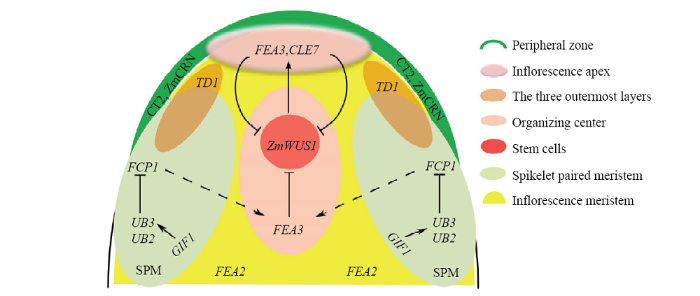
图片显示的是玉米雌花序分生组织和成对小穗分生组织的原基。不同颜色块表示基因表达的组织部位或组织。 表示正调控。 表示负调控 Fig. 1Expression domain and regulatory pathway of genes involved in the regulation of activity of meristems on the ear inflorescence
The figure shows the ear inflorescence meristem and primordia of spikelet-paired meristems. Different color blocks show the domains where genes are expressed.: Positive regulation. : Negative regulation
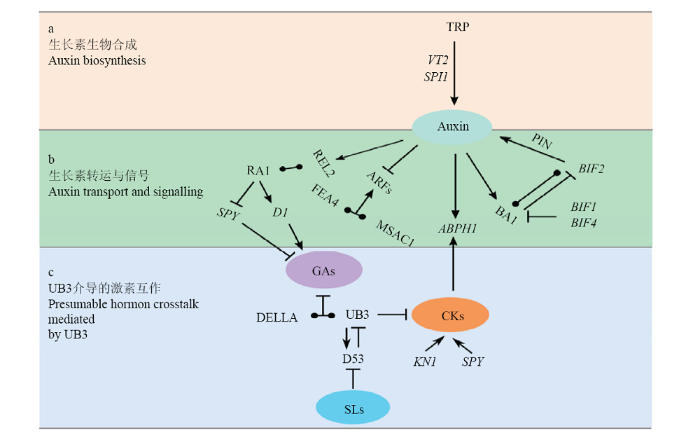
a:SPI1和VT2调控色氨酸依赖的生长素合成。b:生长素转运与信号相关基因及其调控途径。c:UB3介导的激素交互作用。 表示正调控。 表示负调控 Fig. 2Regulation pathways for activity of meristems on the ear inflorescence by genes in hormone biosynthesis and signaling
a: Both SPI1 and VT2 involve in the regulation of Trp-dependent auxin biosynthesis. b: Regulatory pathways of genes in auxin transport and signaling. c: Presumable hormone crosstalk mediated by UB3. : Positive regulation. : Negative regulation
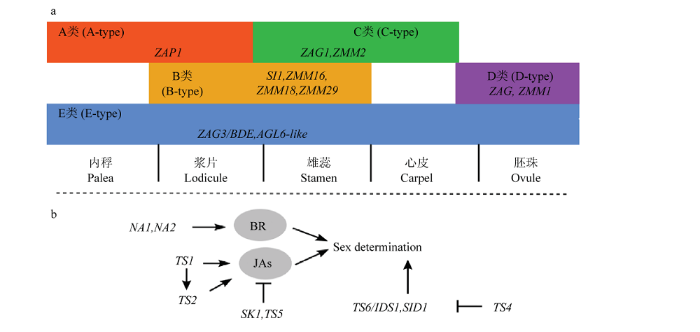
a:玉米花器官发育“ABCDE”模型调控基因。不同色块代表不同类型的花器官调控基因,下方标注代表不同花器官及其对应轮数。A类基因决定第一轮内稃和外稃的形成,第二轮和第三轮浆片和雄蕊的形成分别由A+B和B+C基因调控,第四轮心皮发育则由C类基因单独调控。D类基因主要在胚珠发育中起作用,E类基因则参与所有的花器官发育调控。b:小花性别决定基因调控网络。 表示正调控。 表示负调控 Fig. 3Genes regulating maize floral organs development and floret sex determination
a: The “ABCDE model” of floral organ identity. Different color blocks show different types of floral organ regulatory genes, and the label shows floral organ developed in the whorl. A-type genes determine the identity of palea and lemma. Lodicule and stamen are regulated by A+B and B+C, respectively. Carpel identity is regulated by the C-type genes. D-type genes act in ovule development, and the E-type genes involve in the regulation of development of all floral whirls. b: Floret sex determination genes regulatory network. : Positive regulation. : Negative regulation
除“ABCDE”花器官发育模型相关基因外,对玉米雌、雄性器官选择性发育的调控研究,丰富了对植物小花性别决定的认识。控制分生组织确定性的基因在性别决定中扮演重要角色。TASSELSEED6(TS6)/INDETERMINATE SPIKELET1(IDS1)编码APETALA2(AP2)/ERF转录因子,突变体ts6/ids1的小穗分生组织确定性丧失,产生额外小花,并且雄花序部分小花因心皮发育产生花丝[45],与其功能相似的同源基因SISTER OF INDETERMINATE SPIKELET1(SID1)则具有积加效应[46]。而TS4编码的miR172靶向TS6/IDS1和SID1,负调控它们的表达[47](图3-b)。另外,研究发现茉莉酸(JAs)和油菜素内酯(BRs)可通过调控小花的性别进而调节小花育性和授粉潜力。目前,玉米中已克隆了6个JAs合成和代谢相关基因,其中,TS1编码一个脂氧化酶[48]、TS2编码一种单子叶植物所特有的短链乙醇脱氢酶[49],OPR7和OPR8是2个同源基因,均编码12-氧-植物二烯酸还原酶(12-oxo-phytodienoic acid reductase)[50],这4个基因共同参与JA的生物合成。单突变体ts1、ts2和双突变体opr7opr8均影响JA生物合成,导致JA水平下降,雄穗上部分小花产生花丝[47,48,49]。显性Ts5突变体的雄花序上也发育出雌性小花、产生花丝。TS5编码一个ZmCYP94B1蛋白,参与JA代谢,即负调控JA水平[51](图3-b)。由此可见,高水平的JA是玉米雄花中雌蕊退化或者抑制心皮发育所必要的。那么,玉米雌花中控制雄蕊退化或者保持雌蕊发育的关键基因是什么?SK1编码一个尿苷二磷酸糖基转移酶,该酶可阻断JA生物合成;在sk1突变体中,其雄穗发育正常、但雌穗花丝发育受到抑制[52]。sk1ts2双突变体雌穗花丝恢复,而雄穗大多数小花也能发育雄蕊[53];而sk1Ts5双突变体的雌穗无花丝,雄穗产生少量花丝[51]。结果表明,SK1是雌花中雌蕊发育所必需的,并且阻断JA合成有利于雌蕊发育。除了JA外,BR也在玉米小花性别决定中起着重要作用。例如,NANA PLANT1(NA1)是一个BR生物合成途径的酶编码基因,在花药发育整个过程中均有表达、也在心皮原基表层细胞中表达直至其退化;na1突变体的雄穗部分小花花药退化而花丝发育,并有类似内外稃的变形叶,成熟植株的雄穗上着生有种子,突变体的雌穗和雌花发育正常[54]。NANA PLANT2(NA2)也参与BR生物合成途径,其突变体表现为与na1相似的表型[55]。这说明BR在雄蕊和雌蕊发育中起着不同作用,即BR促进雄蕊发育而抑制心皮分生组织的分化,而调节BR在玉米雌花和雄花发育中不同功能的机理至今未知。
近年来,沉默乙烯合成途径的1-Aminocyclopropane- 1-Carboxylase Synthase6(ACS6),能增加转基因玉米家系在缺水和低氮条件下的产量[56];过表达乙烯信号途径的组分AUXIN-REGULATED GENE INVOLVED IN ORGAN SIZE(ARGOS)可增加转基因家系和杂交种在正常和干旱条件下的穗长、穗粒数及籽粒产量[57]。结果表明,乙烯水平和信号或直接参与花序和小花发育、或通过参与玉米对胁迫的响应间接地影响花序和小花发育。这些发现也指出一条“通过调控乙烯合成和信号进而提高玉米抗性和籽粒产量”的新途径。
VOLLBRECHTE, SCHMIDT RJ . Handbook of maize // BENNETZEN J L, HAKE S, eds. Development of the Inflorescences. New York: Springer, 2009: 13-40. [本文引用: 1]
WILLIAMSL, FLETCHER JC . Stem cell regulation in the Arabidopsis shoot apical meristem Current Opinion in Plant Biology, 2005,8:582-586. [本文引用: 1]
SOMSSICHM, JE BI, SIMONR, JACKSOND . CLAVATA- WUSCHEL signaling in the shoot meristem Development, 2016,143:3238-3248. [本文引用: 1]
BOMMERTP, LUNDEC, NARDMANNJ, VOLLBRECHTE, RUNNINGM, JACKSOND, HAKES, WERRW . thick tassel dwarf1 encodes a putative maize ortholog of the Arabidopsis CLAVATA1 leucine- rich repeat receptor-like kinase Development, 2005,132(6):1235-1245. [本文引用: 1]
BOMMERTP, NAGASAWA NS, JACKSOND . Quantitative variation in maize kernel row number is controlled by the FASCIATED EAR2 locus Nature Genetics, 2013,45:334-337. [本文引用: 1]
JE BI, GRUELJ, LEEK, BOMMERTP, AREVALO ED, EVELAND AL, WUQ, GOLDSHMIDTA, MEELEYR, BARTLETTM, KOMATSUM, SAKAIH, J?NSSONH, JACKSOND . Signaling from maize organ primordia via FASCIATED EAR3 regulates stem cell proliferation and yield traits Nature Genetics, 2016,48:785-791. [本文引用: 3]
RODRIGUEZ-LEALD, XUC, KWON CT, SOYARSC, DEMESA-AREVALOE, MANJ, LIUL, LEMMON ZH, JONES DS, VAN ECKJ, JACKSON DP, BARTLETT ME, NIMCHUK ZL, LIPPMAN ZB . Evolution of buffering in a genetic circuit controlling plant stem cell proliferation Nature Genetics, 2019,51(5):786-792. [本文引用: 1]
BOMMERTP, JE BI, GOLDSHMIDTA, JACKSOND . The maize Gα gene COMPACT PLANT2 functions in CLAVATA signaling to control shoot meristem size Nature, 2013,502:555-558. [本文引用: 2]
JEBI, XUF, WUQ, LIUL, MEELEYR, GALLAGHER JP, CORCILIUSL, PAYNE RJ, BARTLETT ME, JACKSOND . The CLAVATA receptor FASCIATED EAR2 responds to distinct CLE peptides by signaling through two downstream effectors Elife, 2018,7:e35673. [本文引用: 1]
CHUCK GS, BROWNJ, MEELEYR, HAKES . Maize SBP-box transcription factors unbranched2 and unbranched3 affect yield traits by regulating the rate of lateral primordia initiation Proceedings of the National Academy of Sciences of the United States of America, 2014,111:18775-18780. [本文引用: 2]
ZHANGD, SUNW, SINGHR, ZHENGY, CAOZ, LIM, LUNDEC, HAKES, ZHANGZ . GRF-interacting factor1 (gif1) regulates shoot architecture and meristem determinacy in maize The Plant Cell, 2018,30:360-374. [本文引用: 1]
DUY, LIUL, LIM, FANGS, SHENX, CHUJ, ZHANGZ . UNBRANCHED3 regulates branching by modulating cytokinin biosynthesis and signaling in maize and rice New Phytologist, 2016,214(2):721-733. [本文引用: 3]
PHILLIPS KA, SKIRPAN AL, LIUX, CHRISTENSENA, SLEWINSKI TL, HUDSONC, BARAZESHS, COHEN JD, MALCOMBERS, MCSTEENP . vanishing tassel2 encodes a grass-specific tryptophan aminotransferase required for vegetative and reproductive development in maize The Plant Cell, 2011,23:550-566. [本文引用: 2]
GALLAVOTTIA, YANGY, SCHMIDT RJ, JACKSOND . The relationship between auxin transport and maize branching Plant Physiology, 2008,147:1913-1923. [本文引用: 1]
GALLAVOTTIA, ZHAOQ, KYOZUKAJ, MEELEY RB, RITTER MK, DOEBLEY JF, PEME, SCHMIDT RJ . The role of barren stalk1 in the architecture of maize Nature, 2004,432:630-635. [本文引用: 1]
SKIRPANA, CULLER AH, GALLAVOTTIA, JACKSOND, COHEN JD, MCSTEENP . BARREN INFLORESCENCE2 interaction with ZmPIN1a suggests a role in auxin transport during maize inflorescence development Plant and Cell Physiology, 2009,50:652-657. [本文引用: 1]
MCSTEENP, MALCOMBERS, SKIRPANA, LUNDEC, WU XT, KELLOGGE, HAKES . Barren inflorescence2 encodes a co-ortholog of the PINOID serine/threonine kinase and is required for organogenesis during inflorescence and vegetative development in maize Plant Physiology, 2007,144:1000-1011. [本文引用: 1]
BARAZESHS, MCSTEENP . Barren inflorescence1 functions in organogenesis during vegetative and inflorescence development in maize Genetics, 2008,179:389-401. [本文引用: 1]
GALLIM, LIUQ, MOSS BL, MALCOMBERS, LIW, GAINESC, FEDERICIS, ROSHKOVANJ, MEELEYR, NEMHAUSER JL, GALLAVOTTIA . Auxin signaling modules regulate maize inflorescence architecture Proceedings of the National Academy of Sciences of the United States of America, 2015,43:13372-13377. [本文引用: 2]
SKIRPANA, WUX, MCSTEENP . Genetic and physical interaction suggest that BARREN STALK1 is a target of BARREN INFLORESCENCE2 in maize inflorescence development The Plant Journal, 2008,55:787-797. [本文引用: 1]
PAUTLERM, EVELAND AL, LARUET, YANGF, WEEKSR, LUNDEC, JEB, MEELEYR, KOMATSUM, VOLLBRECHTE, SAKAIH, JACKSOND . FASCIATED EAR4 encodes a bZIP transcription factor that regulates shoot meristem size in maize The Plant Cell, 2015,1:104-120. [本文引用: 1]
YANGF, BUI HT, PAUTLERM, LLACAV, JOHNSTONR, LEE BH, KOLBEA, SAKAIH, JACKSOND . A maize glutaredoxin gene, abphyl2, regulates shoot meristem size and phyllotaxy The Plant Cell, 2015,27(1):121-131. [本文引用: 1]
JACKSOND, HAKES . Control of phyllotaxy in maize by the abphyl1 gene Development, 1999,126:315-323. [本文引用: 1]
GIULINIA, WANGJ, JACKSOND . Control of phyllotaxy by the cytokinin-inducible response regulator homologue ABPHYL1 Nature, 2004,430:1031-1034. [本文引用: 1]
JIAOY, WANGY, XUED, WANGJ, YANM, LIUG, DONGG, ZENGD, LUZ, ZHUX, QIANQ, LIJ . Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice Nature Genetics, 2010,6:541-544. [本文引用: 1]
MIURAK, IKEDAM, MATSUBARAA, SONG XJ, ITOM, ASANOK, MATSUOKAM, KITANOH, ASHIKARIM . OsSPL14 promotes panicle branching and higher grain productivity in rice Nature Genetics, 2010,6:545-549. [本文引用: 1]
LIUM, SHIZ, ZHANGX, WANGM, ZHANGL, ZHENGK, LIUJ, HUX, DIC, QIANQ, HEZ, YANG DL . Inducible overexpression of Ideal Plant Architecture1 improves both yield and disease resistance in rice Nature Plants, 2019,5:389-400. [本文引用: 1]
SONGX, LUZ, YUH, SHAOG, XIONGJ, MENGX, JINGY, LIUG, XIONGG, DUANJ, YAOX, LIUC, LIH, WANGY, LIJ . IPA1 functions as a downstream transcription factor repressed by D53 in strigolactone signaling in rice Cell Research, 2017,27:1128-1141. [本文引用: 1]
AMBROSE BA, LERNER DR, CICERIP, PADILLA CM, YANOFSKY MF, SCHMIDT RJ . Molecular and genetic analyses of the silky1 gene reveal conservation in floral organ specification between eudicots and monocots Molecular Cell, 2000,5(3):569-579. [本文引用: 3]
CHENG PC, GREYSON RI, WALDEN DB . Organ initiation and the development of unisexual flowers in the tassel and ear of Zea mays American Journal of Botany, 1983,70:450-462. [本文引用: 1]
MENAM, MANDEL MA, LERNER DR, YANOFSKY MF, SCHMIDT RJ . A characterization of the MADS-box gene family in maize The Plant Journal, 1995,8(6):845-854. [本文引用: 1]
BARTLETT ME, WILLIAMS SK, TAYLORZ, DEBLASIOS, GOLDSHMIDTA, HALL DH, SCHMIDT RJ, JACKSON DP, WHIPPLE CJ . The maize PI/GLO ortholog Zmm16/sterile tassel silky ear1 interacts with the zygomorphy and sex determination pathways in flower development The Plant Cell, 2015,11:3081-3098. [本文引用: 2]
MüNSTERT, WINGEN LU, FAIGLW, WERTHS, SAEDLERH, THEISSENG . Characterization of three GLOBOSA-like MADS-box genes from maize: Evidence for ancient paralogy in one class of floral homeotic B-function genes of grasses Gene, 2001,262(1/2):1-13. [本文引用: 1]
SCHMIDT RJ, VEITB, MANDEL MA, MENAM, HAKES, YANOFSKY MF . Identification and molecular characterization of ZAG1, the maize homolog of the Arabidopsis floral homeotic gene AGAMOUS The Plant Cell, 1993,5(7):729-737. [本文引用: 2]
SCHREIBER DN, BANTINJ, DRESSELHAUST . The MADS box transcription factor ZmMADS2 is required for anther and pollen maturation in maize and accumulates in apoptotic bodies during anther dehiscence Plant Physiology, 2004,134:1069-1079. [本文引用: 1]
THEISSENG, STRATERT, FISHERA, SAEDLERH . Structural characterization, chromosomal localization and phylogenetic evaluation of two pairs of AGAMOUS-like MADS-box genes from maize Gene, 1995,156(2):155-166. [本文引用: 3]
THOMPSON BE, BARTLINGL, WHIPPLEC, HALL DH, SAKAIH, SCHMIDTR, HAKES . Bearded-ear encodes a MADS box transcription factor critical for maize floral development The Plant Cell, 2009,21(9):2578-2590. [本文引用: 2]
MENAM, AMBROSE BA, MEELEY RB, BRIGGS SP, YANOFSKY MF, SCHMIDT RJ . Diversification of C-function activity in maize flower development Science, 1996,274(5292):1537-1540. [本文引用: 1]
CACHARRóNJ, SAEDLERH, THEISSENG . Expression of MADS-box genes ZMM8 and ZMM14 during inflorescence development of Zea mays discriminates between the upper and the lower floret of each spikelet Development Genes and Evolution, 1999,209:411-420. [本文引用: 1]
CIAFFIM, RITAA, ANTONIOO, ENRICOT . Molecular aspects of flower development in grasses Sexual Plant Reproduction, 2011,24:247-282. [本文引用: 1]
CHUCKG, MEELEY RB, HAKES . The control of maize spikelet meristem fate by the APETALA2-like gene indeterminate spikelet1 Genes Development, 1998,12(8):1145-1154. [本文引用: 1]
CHUCKG, MEELEYR, HAKES . Floral meristem initiation and meristem cell fate are regulated by the maize AP2 genes ids1 and sid1 Development, 2008,135(18):3013-3019. [本文引用: 1]
CHUCKG, MEELEYR, IRISHE, SAKAIH, HAKES . The maize tasselseed4 microRNA controls sex determination and meristem cell fate by targeting Tasselseed6/indeterminate spikelet1 Nature Genetics, 2007,39:1517-1521. [本文引用: 2]
ACOSTA IF, LAPARRAH, ROMERO SP, SCHMELZE, HAMBERGM, MOTTINGER JP, MORENO MA, DELLAPORTA SL . tasselseed1 is a lipoxygenase affecting jasmonic acid signaling in sex determination of maize Science, 2009,323:262-265. [本文引用: 2]
DELONGA, CALDERON-URREAA, DELLAPORTA SL . Sex determination gene TASSELSEED2 of maize encodes a short-chain alcohol dehydrogenase required for stage-specific floral organ abortion Cell, 1993,74:757-768. [本文引用: 2]
YANY, CHRISTENSENS, ISAKEITT, ENGELBERTHJ, MEELEYR, HAYWARDA, EMERY R JN, KOLOMIETS MV . Disruption of OPR7 and OPR8 reveals the versatile functions of jasmonic acid in maize development and defense The Plant Cell, 2012,24(4):1420-1436. [本文引用: 1]
LUNDEC, KIMBERLINA, LEIBOFFS, KOO AJ, HAKES . Tasselseed5 overexpresses a wound-inducible enzyme, ZmCYP94B1, that affects jasmonate catabolism, sex determination, and plant architecture in maize Communications Biology, 2019,2:114. [本文引用: 2]
HAYWARD AP, MORENO MA, HOWARD TP, HAGUEJ, NELSONK, HEFFELFINGERC, ROMEROS, KAUSCH AP, GLAUSERG, ACOSTA IF, ET AL . Control of sexuality by the sk1-encoded UDP-glycosyltransferase of maize Science Advances, 2016,2:e1600991. [本文引用: 1]
IRISH EE, LANGDALE JA, NELSON TM . Interactions between tassel seed genes and other sex determining genes in maize Developmental Genetics, 1994,15:155-171. [本文引用: 1]
HARTWIGT, CHUCK GS, FUJIOKAS, KLEMPIENA, WEIZBAUERR, POTLURI DP V, CHOES, JOHAL GS, SCHULZB . Brassinosteroid control of sex determination in maize Proceedings of the National Academy of Sciences of the United States of America, 2011,108(49):19814-19819. [本文引用: 1]
BEST NB, HARTWIGT, BUDKAJ, FUJIOKAS, JOHALG, SCHULZB, DILKES BP . nana plant2 encodes a maize ortholog of the Arabidopsis brassinosteroid biosynthesis protein Dwarf1, identifying developmental interactions between brassinosteroids and gibberellins Plant Physiology, 2016,171:2633-2647. [本文引用: 1]
HABBEN JE, BAOX, BATE NJ, DEBRUIN JL, DOLAND, HASEGAWAD, HELENTJARIS TG, LAFITTE RH, LOVANN, MOH, REIMANNK, SCHUSSLER JR . Transgenic alteration of ethylene biosynthesis increases grain yield in maize under field drought-stress conditions Plant Biotechnology Journal, 2014,12:685-693. [本文引用: 1]
SHIJ, HABBEN JE, ARCHIBALD RL, DRUMMOND BJ, CHAMBERLIN MA, WILLIAMS RW, LAFITTE HR, WEERS BP . Overexpression of ARGOS genes modifies plant sensitivity to ethylene, leading to improved drought tolerance in both Arabidopsis and maize Plant Physiology, 2015,169(1):266-282. [本文引用: 1]
SCANLON MJ, TAKACS EM. Kernel biology //BENNETZEN J, HAKE S, eds.,Handbook of Maize: Its Biology. New York: Springer Press, 2009: 121-143. [本文引用: 1]
LI XJ, ZHANG YF, HOUM, SUNF, SHENY, XIU ZH, WANGX, CHEN ZL, SUN SS, SMALLI, TAN BC . Small kernel 1 encodes a pentatricopeptide repeat protein required for mitochondrial nad7 transcript editing and seed development in maize (Zea mays) and rice (Oryza sativa) The Plant Journal, 2014,79(5):797-809. [本文引用: 2]
YANG YZ, DINGS, WANGY, LI CL, SHENY, MEELEYR . Small kernel2 encodes a glutaminase in vitamin B6 biosynthesis essential for maize seed development Plant Physiology, 2017,174(2):1127-1138. [本文引用: 1]
NEUFFER MG, SHERIDAN WF . Defective kernel mutants of maize: I. Genetic and lethality studies Genetics, 1980,95(4):929-944. [本文引用: 1]
SCANLON MJ, STINARD PS, JAMES MG, MYERS AM, ROBERTSON DS . Genetic analysis of 63 mutations affecting maize kernel development isolated from Mutator stocks Genetics, 1994,136(1):281-294. [本文引用: 1]
FUS, MEELEYR, SCANLON MJ . Empty pericarp2 encodes a negative regulator of the heat shock response and is required for maize embryogenesis The Plant Cell, 2002,14(12):3119-3132. [本文引用: 2]
HECKELT, WERNERK, SHERIDAN WF, DUMASC, ROGOWSKY PM . Novel phenotypes and developmental arrest in early embryo specific mutants of maize Planta, 1999,210(1):1-8. [本文引用: 1]
SHENY, LIC, MCCARTY DR, MEELEYR, TAN BC . Embryo defective12 encodes the plastid initiation factor 3 and is essential for embryogenesis in maize The Plant Journal, 2013,74(5):792-804. [本文引用: 2]
QIW, YANGY, FENGX, ZHANGM, SONGR . Mitochondrial function and maize kernel development requires dek2, a pentatricopeptide repeat protein involved in nad1 mRNA splicing Genetics, 2017,205(1):239-249. [本文引用: 1]
CHENX, FENGF, QIW, XUL, YAOD, WANGQ , SONG R. dek35 encodes a PPR protein that affects cis-splicing of mitochondrial nad4 intron 1 and seed development in maize Molecular Plant, 2017,10(3):427-441. [本文引用: 1]
DAID, LUANS, CHENX, WANGQ, FENGY, ZHUC, QIW, SONGR . Maize dek37 encodes a p-type PPR protein that affects cis-splicing of mitochondrial nad2 intron 1 and seed development Genetics, 2018,208(3):1069-1082. [本文引用: 1]
QIW, TIANZ, LUL, CHENX, CHENX, ZHANGW, SONGR . Editing of mitochondrial transcripts nad3 and cox2 by dek10 is essential for mitochondrial function and maize plant development Genetics, 2017,205(4):1489-1501. [本文引用: 1]
WANGG, ZHONGM, SHUAIB, SONGJ, ZHANGJ, HANL, LINGH, TANGY, WANGG, SONGR . E+ subgroup PPR protein Defective Kernel 36 is required for multiple mitochondrial transcripts editing and seed development in maize and Arabidopsis New Phytologist, 2017,214(4):1563-1578. [本文引用: 1]
LIX, GUW, SUNS, CHENZ, CHENJ, SONGW, ZHAOH, LAIJ . Defective Kernel 39 encodes a PPR protein required for seed development in maize Journal of Integrative Plant Biology, 2018,60(1):45-64. [本文引用: 1]
SUNF, ZHANGX, SHENY, WANGH, LIUR, WANGX, GAOD, YANG YZ, LIUY, TAN BC . The pentatricopeptide repeat protein EMPTY PERICARP8 is required for the splicing of three mitochondrial introns and seed development in maize The Plant Journal, 2018,95:919-932. [本文引用: 1]
CAIM, LIS, SUNF, SUNQ, ZHAOH, RENX, ZHAOY, TAN BC, ZHANGZ, QIUF . Emp10 encodes a mitochondrial PPR protein that affects the cis-splicing of nad2 intron 1 and seed development in maize The Plant Journal, 2017,91(1):132-144. [本文引用: 1]
RENX, PANZ, ZHAOH, ZHAOJ, CAIM, LIJ, ZHANGZ, QIUF . EMPTY PERICARP11 serves as a factor for splicing of mitochondrial nad1 intron and is required to ensure proper seed development in maize Journal of Experimental Botany, 2017,68(16):4571-4581. [本文引用: 1]
SUNF, XIUZ, JIANGR, LIUY, ZHANGX, YANG YZ, LIX, ZHANGX, WANGY, TAN BC . The mitochondrial pentatricopeptide repeat protein EMP12 is involved in the splicing of three nad2 introns and seed development in maize Journal of Experimental Botany, 2019,70(3):963-972. [本文引用: 1]
XIUZ, SUNF, SHENY, ZHANGX, JIANGR, BONNARDG, ZHANGJ, TAN BC . EMPTY PERICARP16 is required for mitochondrial nad2 intron 4 cis-splicing, complex I assembly and seed development in maize The Plant Journal, 2016,85(4):507-519. [本文引用: 1]
LIU YJ, XIU ZH, MEELEYR, TAN BC . Empty pericarp5 encodes a pentatricopeptide repeat protein that is required for mitochondrial RNA editing and seed development in maize The Plant Cell, 2013,25(3):868-883. [本文引用: 1]
SUNF, WANGX, BONNARDG, SHENY, XIUZ, LIX, GAOD, ZHANGZ, TAN BC . Empty pericarp7 encodes a mitochondrial E-subgroup pentatricopeptide repeat protein that is required for ccmFN editing, mitochondrial function and seed development in maize The Plant Journal, 2015,84(2):283-295. [本文引用: 1]
YANG YZ, DINGS, WANG HC, SUNF, HUANG WL, SONGS, XUC, TAN BC . The pentatricopeptide repeat protein EMP9 is required for mitochondrial ccmB and rps4 transcript editing, mitochondrial complex biogenesis and seed development in maize New Phytologist, 2017,214(2):782-795. [本文引用: 1]
LI XL, HUANG WL, YANG HH, JIANG RC, SUNF, WANG HC, ZHAOJ, XU CH, TAN BC . EMP18 functions in mitochondrial atp6 and cox2 transcript editing and is essential to seed development in maize New Phytologist, 2019,221(2):896-907. [本文引用: 1]
GUTIERREZ-MARCOS JF, DAL PRAM, GIULINIA, COSTA LM, GAVAZZIG, CORDELIERS, SELLAMO, TATOUTC, PAULW, PEREZP, DICKINSON HG, CONSONNIG . Empty pericarp4 encodes a mitochondrion-targeted pentatricopeptide repeat protein necessary for seed development and plant growth in maize The Plant Cell, 2007,19(1):196-210. [本文引用: 1]
OFFLER CE, MCCURDY DW, PATRICK JW, TALBOT MJ . Transfer cells: Cells specialized for a special purpose Annual Review of Plant Biology, 2003,54:431-454. [本文引用: 1]
BRYANTN, LLOYDJ, SWEENEYC, MYOUGAF, MEINKED . Identification of nuclear genes encoding chloroplast-localized proteins required for embryo development in Arabidopsis Plant Physiology, 2011,155(4):1678-1689. [本文引用: 1]
SOSSOD, CANUTM, GENDROTG, DEDIEUA, CHAMBRIERP, BARKANA, CONSONNIG, ROGOWSKY PM . PPR8522 encodes a chloroplast-targeted pentatricopeptide repeat protein necessary for maize embryogenesis and vegetative development Journal of Experimental Botany, 2012,63(16):5843-6857. [本文引用: 1]
LIC, SHENY, MEELEYR, MCCARTY DR, TAN BC . Embryo defective 14 encodes a plastid-targeted cGTPase essential for embryogenesis in maize The Plant Journal, 2015,84(4):785-799. [本文引用: 1]
ZHANG YF, HOU MM, TAN BC . The requirement of WHIRLY1 for embryogenesis is dependent on genetic background in maize PLoS ONE, 2013,8(6):e67369. [本文引用: 1]
MAZ, DOONER HK . A mutation in the nuclear-encoded plastid ribosomal protein S9 leads to early embryo lethality in maize The Plant Journal, 2004,37(1):92-103. [本文引用: 2]
LIQ, WANGJ, YEJ, ZHENGX, XIANGX, LIC, FUM, WANGQ, ZHANGZ, WUY . The maize imprinted gene Floury3 encodes a PLATZ protein required for tRNA and 5S rRNA transcription through interaction with RNA Polymerase III The Plant Cell, 2017,29(10):2661-2675. [本文引用: 1]
LIJ, FUJ, CHENY, FANK, HEC, ZHANGZ, LIL, LIUY, ZHENGJ, REND, WANGG . The U6 Biogenesis-Like 1 plays an important role in maize kernel and seedling development by affecting the 3' end processing of U6 snRNA Molecular Plant, 2017,10(3):470-482. [本文引用: 1]
ZUOY, FENGF, QIW, SONGR . Dek42 encodes an RNA-binding protein that affects alternative pre-mRNA splicing and maize kernel development Journal of Integrative Plant Biology, 2019,61(6):728-748 [本文引用: 1]
WANGH, WANGK, DUQ, WANGY, FUZ, GUOZ, KANGD, LI WX, TANGJ . Maize Urb2 protein is required for kernel development and vegetative growth by affecting pre-ribosomal RNA processing New Phytologist, 2018,218(3):1233-1246. [本文引用: 1]
HEY, WANGJ, QIW, SONGR . Maize Dek15 encodes the Cohesin- Loading Complex Subunit SCC4 and is essential for chromosome aegregation and kernel development The Plant Cell, 2019,31(2):465-485. [本文引用: 1]
 ,华中农业大学作物遗传改良国家重点实验室,武汉 430070
,华中农业大学作物遗传改良国家重点实验室,武汉 430070
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}