 ,11
,11 2
Effect of Midgut Specific Binding Protein ABCC1 on Cry1Ac Toxicity Against Helicoverpa armigera
CHEN Lin1, WEI JiZhen2, LIU Chen1, NIU LinLin1, ZHANG CaiHong1, LIANG GeMei,11 2
ͨѶ����:
���α༭: ��÷
�ո�����:2019-04-23��������:2019-05-27�����������:2019-10-01
| ��������: |
Received:2019-04-23Accepted:2019-05-27Online:2019-10-01
����� About authors
����,E-mail��chenlincaas@126.com��

ժҪ
�ؼ��ʣ�
Abstract
Keywords��
PDF (857KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر���
�������ø�ʽ
����, κ����, ����, ţ����, �Ųʺ�, ����÷. ������г������Խ�ϵ���ABCC1��Cry1Ac������Ӱ��[J]. �й�ũҵ��ѧ, 2019, 52(19): 3337-3345 doi:10.3864/j.issn.0578-1752.2019.19.005
CHEN Lin, WEI JiZhen, LIU Chen, NIU LinLin, ZHANG CaiHong, LIANG GeMei.
0 ����
���о����塿���ƽ�ѿ�߸˾���Bacillus thuringiensis,Bt�������ľ��嵰���۳�Ŀ��˫��Ŀ���ʳ�Ŀ�ȶ���������߳���������Զ���,����������[1,2,3,4]��Bt�ܹ���������ɱ����Եİ��߾���,Cry�����ǰ��߾����е���Ҫһ��,Ŀǰ�ѷ���Լ��300���ֿ��Է�Ϊ40����ͬ��С��Cry����,Cry1Ac�������������类Ӧ���ں�����ε�[1,2,3,4]������Bt���Ŀ���ת����ũ��������������������ӡ�ȡ��й��ȶ�����ҹ㷺��ֲ,�ⲻ��������Ч�������ũҵ�����Σ��,�����Լ������ڻ�ѧũҩ����ʹ�ô����Ļ�����Ⱦ[4,5]����������תBt����ũ������ֲ���������,�����Bt���Ŀ���[3,6-8]����Ҳ�������ء����,�о������Bt���ײ������ԵĻ��������ӳٿ��Բ���������Ҫ����ǰ���о���չ��������ȡʳBt����,Bt�����������г����Ի����б��ܽ⡢�,�����г���Ƥϸ���ϵ�������������,�������г�Ĥ����,ϸ���ܽ�,���յ�����������[3]��Bt�����������г����������䷢��ɱ�����õĹؼ�,��Ҫ���嵰�ױ������ı仯�����ͻ�������������Bt���ײ������Ե���Ҫԭ��Ŀǰ��������Ҫ���嵰���и�𤵰�ף�cadherin,CAD��������øN��aminopeptidase N,APN��������������ø��alkaline phosphatase,ALP����������������ת�˵��ף�ATP-binding cassette transporter,ABC����[9,10,11,12,13,14,15]��ABCת�˵����嵰�װ���4�����Ľṹ��������Ĥ�ṹ��TMD��,ÿ���ṹ����6����Ĥ���������;����λ�ڰ��ʲ�ĺ�����������NBD��,��������Ĥ�ṹ���档ABCת�˵��Ĺ�����ϸ������ת�˺�������ʵĴ�л�й�[16,17,18],���б�������,ABCת�˵��ײ�����Bt������,����ABC�����ͻ���������ĸı��������Bt�����������[14,15,16]�������о�����㡿ǰ���о�����,��Bt������������棨Helicoverpa armigera����HaABCC2����ͻ��,���HaABCC2����ĵ���������ǰ��ֹ,HaABCC2������ɥʧ,�ᵼ��������Cry1Ac�����ԵIJ���[14]��ABCת�˵�����һ���Ӵ�ļ���,����ϸ��Ĥ���Զ���ABC����ʽ��ϴ���,����о�����ABC��Cry1Acɱ����Ƽ�������Cry1Ac���Ի����е����ú��б�Ҫ���������Ĺؼ����⡿��ȷ�����HaABCC1�Ƿ�ΪCry1Ac�Ĺ��������嵰��,HaABCC1�������ı仯�Ƿ���������Cry1Ac�Ŀ����й�,��һ����ʾCry1Ac��ɱ����Ƽ������������Եķ��ӻ���,Ϊ����Ӧ��Bt��ת��������ƶ������Ŀ������������ṩ�������ݡ�1 �����뷽��
������2017��2018�����й�ũҵ��ѧԺֲ�ﱣ���о���ֲ�ﲡ�溦����ѧ�����ص�ʵ������ɡ�1.1 ��������
�������������Ʒϵ��96S����1996��6�²��Ժ���ʡ����������,��ʵ���������˹�����[19]��������,δ�Ӵ����κ�ɱ��������������Ʒϵ���������������¶ȣ�27��2����,������14L�s10D,���ʪ�ȣ�75��10��%���������������ɳ��ڲ���������������10%����ˮ����Ӫ���������Cry1Ac����Ʒϵ��BtR����������3.6 mg��mL-1 Cry1Acԭ����96S����Ʒϵɸѡ��210��,���Ա���Ϊ3 600��[20]��
1.2 Cry1Ac����
���Cry1Ac�����ڱ�����ʿ���Ƽ�����˾��Envirologix���������ر���Լ��й����人������������Ƽ��ɷ�����˾��Elabscience�����������ر�ǻ��Cry1Ac����,��ǹ��̲ο��Լ���˵���顣1.3 ȡ������
�ռ�����Ʒϵ��96S���뿹��Ʒϵ��BtR�������5���׳��10ͷ,�ڱ��Ͻ���,ȥ��ʳ���ȡ�г�,��4��Ԥ���0.7% NaCl��Һ���ں����ϴ�ɾ�,������ֽ����ˮ��,-80�汣�汸�á�1.4 ��RNA����ȡ��cDNA�ĺϳ�
����Invitrogen����˵���ֱ���ȡ����Ʒ��RNA,RT-PCR��ӫ�ⶨ��RT-PCR���õ�cDNAģ��ĺϳɲ���Tiangen��˾Fast QuantRT Kit˵�����SuperRealPreMix��Probe��˵���顣������Ʒ��3������ѧ�ظ���1.5 ԭ�˱��PLigand blot����
����Primer 5.0�����ֱ����ABCC1������Ĥ����TMD��Я��ø��λ��EcoRV��Hind��������1��,���Ʒֱ�ΪTM1-EcoRV-F��TM1-HindIII-R;TM2-EcoRV-F��TM2-HindIII-R����PCR�ϳɵ�Ŀ�Ļ���ת���¡����,ȡ������ȷ�������ͱ�������pET32a����˫ø��,��T4-DNA����ø��Ŀ��Ƭ�κͱ������幹��Ϊ������������������ȷ����������ת��BL21��DE3������̬ϸ���в�ת��500 mL LB������������,����0.8 mmol��L-1 IPTG����25���ҹ�յ�������ٵ��������ռ����岢��������,��SDS-PAGE��Ӿ����ȡĿ��Ƭ�����������������з����Ľ�������⡣���ù����������﹤������˾��PAGE�������������Լ��л���Ŀ�ĵ��ס�Table 1
��1
��1��RT-PCR��ӫ�ⶨ��RT-PCR��RNAi��Ӧ��������
Table 1
| �������� Primer name | �������� Primer sequence (5��- 3��) |
|---|---|
| ABCC1-F | TTAATTAACCAGTCCGGCGTCC |
| ABCC1-R | ATGTCTTACAATTCTACGCTTGA |
| GAPDH-F | CATTGAAGGTCTGATGACCACTGT |
| GAPDH-R | CAGAGGGTCCATCCACTGTCTT |
| GAPDH-Probe | CACGCCACCATTGCCACCCA |
| ��-actin-F | GGCCCCGTCCACAATGA |
| ��-actin-R | CCGATCCATACGGAGTACTTCCT |
| ��-actin-Probe | ATCAAGATCATCGCGCCCCCAGA |
| ABCC1-F | GGCAGCGTGAAAAGAAAGAC |
| ABCC1-R | GTGATCACGGACGAGAGGAT |
| ABCC1-Probe | CGAAGCTGAGAAAACCGAGACTGGAAG |
| TMD1-EcoRV-F | GATATCGCGTTCGGCGGTCAGTTC |
| TMD1-HindIII-R | AAGCTTACGTGACATCGAGTTCATCACAATT |
| TMD2-EcoRV-F | GATATCTTGATGAGCGTCGGAATCCT |
| TMD2-HindIII-R | AAGCTTATGTTTCTACTTCTGATGTCATGCGTAC |
| siRNA-ABCC1 | GGAUGUACCUGGUGGGCAUTT |
| siRNA-EGFP | GCGUUGGGAAGUCAAGUUUTT |
| ABCC1-PmeI-F | GTTTAAACATGTCTTACAATTCTACGCTTG |
| ABCC1-StuI-R | AGGCCTTTAATTAACCAGTCCGGCGTCC |
| ABCC1-F�� | AGAAGCCTGCCTCCATACTACC |
| ABCC1-R�� | AAGAAGCGCAACTGCATAAAC |
�´��ڴ�|����CSV
��Ligand blot���ԭ�˱���õ���HaABCC1������Ĥ��Ƭ�ε����ܷ�����Cry1Ac�������ϡ���������������Ӿת��PVDFĤ��;����30 mL���Һ���Է���в�����PVDFĤ,����ҡ����80 r/min���·���2 h;���¼����µ�30 mL��֬ţ���ټ���5 ��L�����ر�ǵĻCry1Ac,����ҡ����80 r/min����1 h;���¼���20 mL��֬ţ���ټ���2 ��L������HRP��ǵ�ɽ��С���壩���Է����,���·���1 h,PBSTϴĤ3��,ÿ��10 min;��ECL���������ع�,���õ��Լ���ΪEASYsee�����Һ��
1.6 RNAi������ⶨ
����RNAi��������Ʒϵ��96S��������HaABCC1��Ϊ�����RNAi���ŵ�������,�Ӹ������Ե�HaABCC1��Ĥ��Ƭ�����siRNA���С����ü�ǿӫ�⵰����EGFP����Ϊ���ա�siRNA���������﹤������˾�ϳɣ���1��,���Ʒֱ�ΪsiRNA-ABCC1��siRNA-EGFP����5 ��L��ע������3���������׳�ĸ���ע��2 ��g����L-1��siABCC1 1 ��L��ע����÷�ʿ�ַ��,��ֹ��Һ������ʧ���ò���ע���������Լ�ע��siEGFP��DEPCˮ���������Ϊ���ա�ÿ������24ͷ�����,����3���ظ������ź�48 h����ӫ�ⶨ��RT-PCR��ⲻͬ���������HaABCC1�ı�������
��������ⶨ�ķ����ⶨCry1Ac�Ը��������HaABCC1�������ʵı仯����24�װ��ÿ�����м���1 mL��������ɵ�������˹����ϣ�Һ̬�����������̺���ÿ�����м���75 ��L���Cry1Ac����ʹÿ�����е���Ũ��Ϊ120 ��g��cm-2 ��������Ŵ���48 h��������,ÿ������24ͷ,3���ظ����Լ��������PBS��ҺΪ���մ�����5 d��,ͳ�Ƹ���������������,������Ӧ��������[21]��
1.7 ϸ��תȾ��ϸ������ⶨ
����Primer 5.0������ƴ�ø��λ��PmeI��StuI��HaABCC1ȫ������ABCC1-PmeI-F��ABCC1-StuI- R����1������PCR���������ȷ������ͨ��˫ø�к�T4-DNA����ø���ӵ�pAc5.1b������,������������pAc-ABCC1���������ڱ�����Խ����������˾����Sf9ϸ���ֱ�ƽ�̵�12�װ���,ÿ��Լ9��105��ϸ��,�ŵ�28����������ϸ��������ڣ�2 h������Cellfectin��Invitrogen;8 ��L/�ף�������,ÿ������תȾ2 ��g pAc5.1b������,����������Ŀ�Ļ�������,����5 h����1.5 mL Sf-900 II SFM��������Sf9ϸ�����û�תȾҺ,Ȼ�����28������������64 h����ÿ�ζ�����תȾ������,ϸ��������,����ƽ�̵�96�װ��3������,ÿ��100 ��L��ϸ��10 000�����������2 h��,�û��Cry1Ac����תȾϸ��,���Cry1AcŨ��Ϊ18.5 ��g��mL-1��ÿ���������ظ�3��תȾ��ϸ����,ÿ�ζ�����תȾ�û��Cry1Ac����3���ظ�������ⶨ����ϸ������ⶨ��,Sf9ϸ����ȡRNA����ת¼ΪcDNA,��HaABCC1����������ABCC1-F����ABCC1-R������1������PCR��֤,����pAc-ABCC1���������Ƿ�ɹ�ת��Sf9ϸ���С�
1.8 ���������жԱȷ���
����GenBank��HaABCC1��cDNA���У�KY796050��,��Primer 5.0�����ֱ����ABCC1-F��ABCC1-R�����1��,�Կ����������5���г���cDNAΪģ��,����PCR����,�ֱ�������ABCC1���С������г�������PCR��94��Ԥ����5 min;94�����30 s,55���˻�30 s,72������5 min,��35��ѭ��;72�汣��10 min��PCR��Ӧ���õĸ߱���ۺ�ø������Ĭ�������Ƽ����й�������˾��Thermo Fisher����������ɺ���1%��֬��������Ӿ���,����AxygenDNA�����Լ��л���Ŀ������,Ȼ��ѻ��ղ����¡��Peasy-T3������,ת����Trans1-T1����̬ϸ����,��ȡ���Ե���¡�ͱ��������£�Biomed���������вⶨ��PCR��û���ȫ������Blast���бȶ�ȷ��ΪHaABCC1��,��DNAMAN�ֱ�Ա����Ӧ�İ��������С�1.9 ӫ�ⶨ��RT-PCR
����TaqMan̽�뼼������ӫ�ⶨ��RT-PCR,����HaABCC1��cDNA������ơ��ϳ�����ӫ�ⶨ��RT-PCR�������������̽��,�������GAPDH��GenBank��JF417983.1������-Actin��GenBank��EU527017.1����Ϊ˫�ڲλ���,�ϳ������������̽��,̽��5��˲���FAM���,3��˲���BHQ��ǡ������̽�����Ϻ�Invitrogen��˾�ϳɣ���1������20 ��Lӫ�ⶨ��RT-PCR��Ӧ��ϵ�к��У�2�� SuperReal Premix 10 ��L�������10 ��mol��L-1��0.6 ��L���������10 ��mol��L-1��0.6 ��L��ӫ��̽�루10 ��mol��L-1��0.4 ��L��cDNAģ��1 ��L��50��ROX ReferenceDye 0.2 ��L��RNase-Free ddH2O 7.2 ��L���������ĺ���ABI 7500ӫ�ⶨ��RT-PCR���ϰ�������������Ӧ��95��Ԥ����15 min��Ȼ��95�����3 s��60���˻�/����32 s����40��ѭ������Ӧ����ˮ��Ϊ���Զ��գ�ÿ������3�������ظ���1.10 ���ݴ���
��ӫ�ⶨ��RT-PCR�������2-����Ct�����м���[22]������SPSS�����Դ������ݽ��з���,������Ƚϲ��õ����ط���������,��Turkey�����ж��رȽϡ�2 ���
2.1 HaABCC1��Ĥ��ԭ�˱��PLigand blot����
ͨ��PCR����,����������֤�ɹ���¡�õ�HaABCC1��Ĥ��TMD1��924 bp����TMD2��882 bp��,����E. coli BL21��DE3������̬ϸ���гɹ�ԭ�˱���,����Ƭ�δ�С�ֱ�Ϊ34��33 kD,��Ԥ�ڽ�������ͼ1-A��������Ligand blot���,HaABCC1��Ĥ��ԭ�˱���Ƭ�ξ�������Cry1Ac����ϣ�ͼ1-B����ͼ1
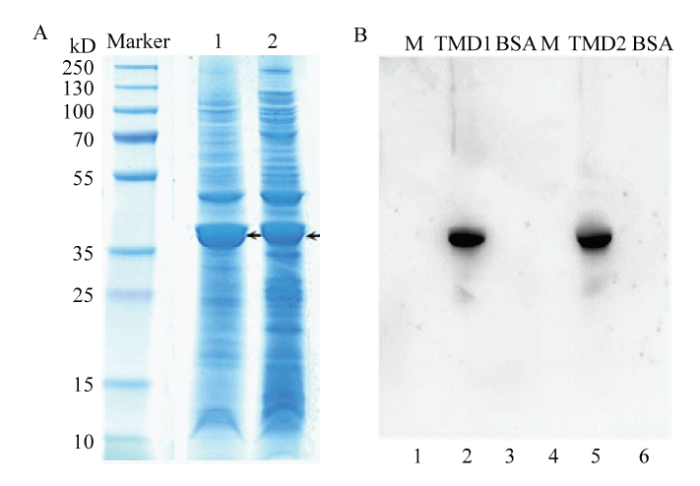 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ1ԭ�˱���HaABCC1Ƭ�ε�SDS-PAGE��Ӿͼ��Ligand blot ���Cry1Ac��HaABCC1�Ľ��
A��Marker�����ױ�ƷProtein marker;1���������鴦����TMD1Ƭ��TMD1 fragment after ultrasound treatment;2���������鴦����TMD2Ƭ��TMD2 fragment after ultrasound treatment��B��1��4�����ױ�ƷProtein marker;3��6��BSA����,��Ϊ����BSA, as a control;2��TMD1Ƭ������Cry1Ac�����TMD1 fragment bound with Cry1Ac;5��TMD2Ƭ������Cry1Ac�����TMD2 fragment bound with Cry1Ac
Fig. 1SDS-PAGE of HaABCC1 fragments by prokaryotic expression and Ligand blot detection of Cry1Ac binding with HaABCC1
2.2 HaABCC1������֤
HaABCC1�����ź�,�����δע���������Լ�ע��DEPCˮ��siEGFP�������,HaABCC1�ı������ֱ��½���54.0%��49.4%��52.2%��P��0.05����ͼ2-A����HaABCC1�����ź�������,�����������Խ�����,�׳��Cry1Ac��������Ҳ���������ı䡣�û��Cry1Ac���״���HaABCC1�����ŵ������,��δע���������Լ�ע��DEPCˮ��siEGFP����������,���׳������ʷֱ��½���65.9%��60.0%��62.2%��P��0.05����ͼ2-B����ͼ2
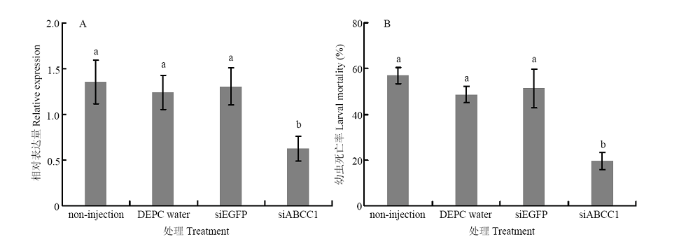 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ2�����Ĭ�������HaABCC1�ı�������A������Cry1Ac�����Ե�Ӱ�죨B��
���ϲ�ͬСд��ĸ��ʾ���������������P��0.05������ͬ
Fig. 2HaABCC1 expression after gene silencing (A) and its effect on H. armigera susceptibility to Cry1Ac (B)
Different lowercases indicate significant differences among treatments (P��0.05). The same as below
��ת��pAc��������Sf9ϸ�����,���Կ��������pAc-ABCC1���ɹ�����Sf9ϸ���У�ͼ3-A������18.5 ��g��mL-1�ĻCry1Ac���״����ɹ�ת��HaABCC1��Sf9ϸ��,�������ϸ��������������������P��0.05����ͼ3-B��,Ϊ33.8%,��������δת��HaABCC1��Sf9ϸ����ϸ�������ʣ�2.0%����
ͼ3
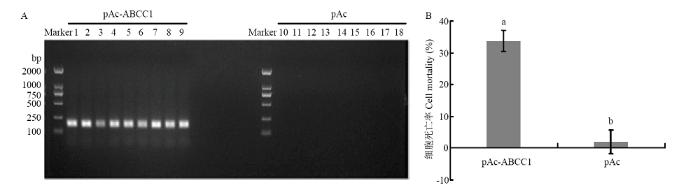 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ3�� pAc-ABCC1ת��Sf9ϸ�����û��Cry1Ac���״�������������
A��PCR���pAc-ABCC1����ת��Sf9ϸ��Confirmation of HaABCC1 expression in cells transfected with pAc-ABCC1 by PCR��1��9��ת��pAc-ABCC1������Sf9ϸ����3������ѧ�ظ��� 3�������ظ���pAc-ABCC1 plasmid transfected into the Sf9 cell line (3 biological replicates �� 3 technical replicates);10��18��ת��pAc��������Sf9ϸ����3������ѧ�ظ��� 3�������ظ���pAc empty plasmid transfected into the Sf9 cell line (3 biological replicates �� 3 technical replicates)��B���û��Cry1Ac���״���ת��pAc-ABCC1������ת��pAc��������Sf9ϸ��������The mortality of Sf9 cells exposed to Cry1Ac after transfected with the pAc-ABCC1 or pAc empty plasmid
Fig. 3Effects of pAc-ABCC1 plasmid transfection into Sf9 cells on the mortality after treated with activated Cry1Ac
2.3 ������Ʒϵ�����HaABCC1��cDNA�����������жԱȷ���
�Ƚ�Cry1Ac���������������������HaABCC1ȫ��,����HaABCC1��Cry1Ac����Ʒϵ��BtR��������Ʒϵ��96S�������ֻ����������IJ���,�ڰ�����ˮƽ��û���κβ��졣2.4 ������Ʒϵ�����HaABCC1��mRNA����������
��һ���Ƚ���HaABCC1�ڿ���Ʒϵ��BtR��������Ʒϵ��96S��������б������IJ���,����HaABCC1�ڿ���Ʒϵ������еı������������ͣ�P��0.05��,����Ʒϵ�ı������ǿ���Ʒϵ��4.19����ͼ4����ͼ4
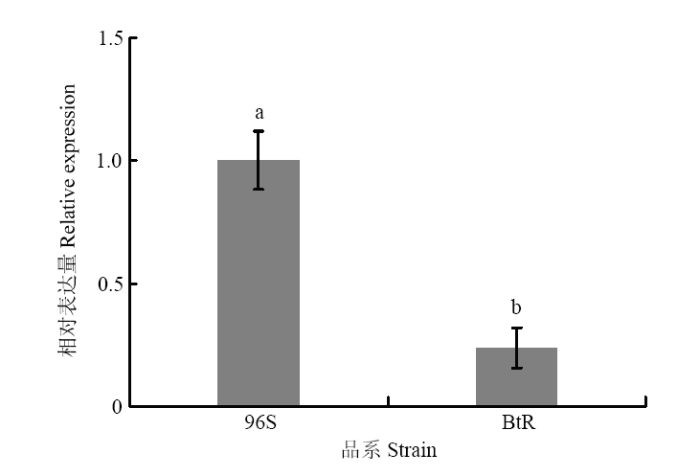 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ4HaABCC1�ڲ�ͬƷϵ�еı�����
Fig. 4The expression level of HaABCC1 in different strains of H. armigera
3 ����
�����г���Ƥϸ��ˢ״ԵĤ����BBMV�ϵ���������Bt���������Խ����Btɱ����������Źؼ�����[23,24]���Ѿ�����ر���֤ʵABCת�˵�����Bt�������塣����,ZHOU��[15]ͨ������������������,����Cry1Ac����ABCC2���;TANAKA��[25]������ϸ���ڷֱ����ͻ���ͣ��Ұ�����룩��Ұ���ͼҲϵ�ABCC2,ͨ������ϸ����Bt�������Ա仯����ABCC2������Cry1Ab��Cry1Ac��Cry1Fa��Cry8Ca��ɱ�����;CHEN��[26]ͨ��������ϸ��ϵ����б��ҹ�꣨Spodoptera litura Sl-HP��ABCC3,֤ʵ��ΪCry1Ac�Ĺ������塣���о�ѡ����HaABCC1ת�˵����������Խϸߵ�������Ĥ�ṹ��,ͨ���ڴ˾���ԭ�˱���õ�������HaABCC1��Ĥ�ṹ��,ͨ��Ligand blot����ȷ����������Ĥ��Ƭ�ε�������Cry1Ac�������ϡ�HaABCC1�����ź�������Cry1Ac����������������,������Sf9ϸ���б���HaABCC1�ܹ����Cry1Ac�������ϸ�������ʡ���Щ�������HaABCC1��Cry1Ac�������Խ�ϵ���,������Cry1Ac�Ĺ��������嵰�ס�ǰ���о�����,HaABCC1������治ͬ�����ξ��б���,��4�䡢5���׳��б��������,�ڳɳ������;�ڲ�ͬ��֯�о��б���,�����Ϲ��б��������,��ǰ������͡��������ABCת�˵�����Ĺ������,���ڲ����ද����ABCת�˵��ײ�����������Ͷ��ط��ڵĹ���,�ܹ�����ͬ����Դ���ʡ��ؽ��������䵽����,��˾��е������塢��ѧ����ҩ���ũҩ�Ĺ���[27]��������,����ABCת�˵��ı仯�������Bt�Ŀ��Թ�ϵҲ�ܵ��ձ��ע�����۳�Ŀ�����ABCB��ABCD��ABCE��ABCF֮�����һ����ͬԴ�����������Bt�Ŀ����й�[28];����������HaABCC2����ͻ��,����䷭��ĵ���������ǰ��ֹ,HaABCC2������ɥʧ,��ᵼ��������Cry1Ac�����ԵIJ���[14];С�˶꣨Plutella xylostella��ABCC2ͻ����������Cry1Ac�����ԵIJ���[29];С�˶�ABCH1�����ź�,������������Cry1Ac���Ŀ���[30];�����ABCA2��ͻ�������������Cry2Ab���Ŀ���[31]�����о�����,��ȻCry1Ac���������HaABCC1����İ���������û�з����仯,��������Ʒϵ���,����Ʒϵ������HaABCC1��������������,�Ʋ�HaABCC1�������Ľ��Ϳ�����������Cry1Ac���ԵIJ�����ء�
���������HaABCC1������������ABC���������ƵĽṹ,����2����Ĥ�ṹ���2��Ĥ������,����14��N-�ǻ���λ���16��O-�ǻ���λ�㡣������2�������ǻ���λ��Ŀ�Ĥ�ṹ�������Btɱ�����ü������Bt�Ŀ�����ء��й�ABCC2��Btɱ������е�����,GAHAN��[32]�Ʋ�Cry������ABCC2��Ͽ������γɹѾ���Ļ���,���Ѵ���ǰ�Ľṹ��������Ĥ���Ա��ڴ���;TANAKA��[25]�о�����,��ϸ���ڹ������𤵰�ף�CaLP����ABCC2,��������ϸ���Ƚ�����ABCC2ʱ����;���Ƶ�,��Sf9ϸ���й�����HevABCC2��Heliothis virescens����HevCaLP,��ϸ���и��ߵĶ���;��TAY���Ʋ�ABCA2���Զ�������Cry2Ab������ģ��,����ɽ�Ϻʹ���[31];��˱�����мҲ�ABCC2,��������û���Э������[33];ABCC2�����κ��ó������谭��Bt�������û��Ƶ����һ��[24]�����ڲ�ͬABCת�˵���֮�����ƵĽṹ,�����ABCC1Ҳ����������ABCת�˵���һ��,������Ĥ�ṹ��ı仯�����������Bt�Ŀ�����ء������о���û�з���HaABCC1������Ĥ�ṹ��İ��������з����ı�,HaABCC1ȫ������������Ҳû�б仯,��HaABCC1�����������˸ı䡣����ABC������һ���Ӵ�ļ���,����ϸ��Ĥ��Ҳ��ϴ���,�п����Ǻܶ��ֵ���ͬ�����á����,�����Ʋ�ABCC1���ܺ�����ABC���嵰�����������嵰��ͬ��Btɱ����Ƽ����Ի����������á�
TAY��[31]����EPIC��exon-primedinton-crossing��������Ŀ�Ļ���IJ�����,ͨ��˫���Ŵ����������˿�Cry2Ab�����,�����俹�ԵIJ���������ATP��Ϻ�ת�������ABCA�ϵ�3���뿹����ص�INDEL��ͻ�������,��HaABCA1��HaABCA2��ͻ����������Cry2Ab�Ŀ������;XIAO��[14]Ҳ������HaABCC2��ͻ������������Cry2Ab�Ŀ��ԡ�ǰ���о�����HaABCC1�����ź�,������Cry2Ab����������������,�Ʋ�ABCC1Ҳ��Cry2Ab�Ĺ��������嵰��[34]�������������,ABCת�˵����ܲ�����Cry1A�����嵰��,Ҳ��Cry2A������,��Щ��ͬ�������Ƿ���Cry1A��Cry2A�൰��Ľ����������,�д���һ���о���
4 ����
ԭ�˱�������������HaABCC1��Ĥ��Ƭ�ε�������Cry1Ac��������;HaABCC1�����ź�,����������Cry1Ac�������Ķ���;��HaABCC1����Sf9ϸ�������������Cry1Ac�������������;������Ʒϵ���,Cry1Ac����ƷϵHaABCC1�������������͡����,HaABCC1������Cry1Ac�Ĺ������嵰��,�����ܲ����Cry1Ac�Ŀ��Ի��ơ��ο����� ԭ��˳��
������ȵ���
������������
�����ڿ�Ӱ������
[��������: 2]
[��������: 2]
[��������: 4]
[��������: 3]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 5]
[��������: 3]
[��������: 2]
[��������: 1]
Magsci [��������: 1]

�����������Ϻ�ת�˵��ף�ATP-binding cassette transporter)�����ABCת�˵��ף�ABC transporter�����Ǽ�ϸ��ɫ��P450������ø��������ø��������S��ת��ø֮����һ����Ҫ�Ľⶾøϵ��������ɱ����ⶾ�ȷ������ŷdz���Ҫ�����ã����������ܵ��㷺��ע��ABCת�˵�����һ�����Ĥ���ף�����Ľṹͨ����4���ṹ����ɣ�����2���߶���ˮ�Ŀ�Ĥ�ṹ��transmembrane domains , TMD����2������������nucleotide binding domains, NBD�����������������Ժͱ��ؽṹ����ABCת�˵������Ϊ8���Ǽ��壬ÿ���Ǽ���ij�Ա�������ܲ�ͬ�����൰���ڸ����������ھ��зֲ�������Ҫ���ܰ���ת�����ʡ��źŴ�����ϸ���������弰����ϸ����DNA����ת¼�����ڻ���ı�����̵ȡ����⣬���������о�������ABCת�˵���ͻ�������ﲻ�����֫����Ի�ѧũҩ�Ŀ�ҩ��������أ������ڿ�Bt���ط���Ҳ���ŷdz���Ҫ�����ã���תBt�������������в�����������˽�֫����ABCת�˵��Ľṹ��ATPˮ��鵼�����û��ƣ��Ǽ���ķ��ࡢ�ṹ���������ܣ��Լ���ABCת�˵��鵼�Ŀ�ҩ���о���չ��ּ�������˽�ABCת�˵����о���״�����ڽ�֫���ҩ�Է�������ã�Ϊ������֫���ҩ�Ի����ṩ�µ��������ݣ��ԸĽ�ũҵ����Ŀ��Լ�����������Ҳ����һ����ָ�����塣��
Magsci [��������: 1]
�����������Ϻ�ת�˵��ף�ATP-binding cassette transporter)�����ABCת�˵��ף�ABC transporter�����Ǽ�ϸ��ɫ��P450������ø��������ø��������S��ת��ø֮����һ����Ҫ�Ľⶾøϵ��������ɱ����ⶾ�ȷ������ŷdz���Ҫ�����ã����������ܵ��㷺��ע��ABCת�˵�����һ�����Ĥ���ף�����Ľṹͨ����4���ṹ����ɣ�����2���߶���ˮ�Ŀ�Ĥ�ṹ��transmembrane domains , TMD����2������������nucleotide binding domains, NBD�����������������Ժͱ��ؽṹ����ABCת�˵������Ϊ8���Ǽ��壬ÿ���Ǽ���ij�Ա�������ܲ�ͬ�����൰���ڸ����������ھ��зֲ�������Ҫ���ܰ���ת�����ʡ��źŴ�����ϸ���������弰����ϸ����DNA����ת¼�����ڻ���ı�����̵ȡ����⣬���������о�������ABCת�˵���ͻ�������ﲻ�����֫����Ի�ѧũҩ�Ŀ�ҩ��������أ������ڿ�Bt���ط���Ҳ���ŷdz���Ҫ�����ã���תBt�������������в�����������˽�֫����ABCת�˵��Ľṹ��ATPˮ��鵼�����û��ƣ��Ǽ���ķ��ࡢ�ṹ���������ܣ��Լ���ABCת�˵��鵼�Ŀ�ҩ���о���չ��ּ�������˽�ABCת�˵����о���״�����ڽ�֫���ҩ�Է�������ã�Ϊ������֫���ҩ�Ի����ṩ�µ��������ݣ��ԸĽ�ũҵ����Ŀ��Լ�����������Ҳ����һ����ָ�����塣��
[��������: 1]
[��������: 1]
[��������: 1]
Magsci [��������: 1]
<P class=MsoNormal ><FONT size=3><SPAN >����ø</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">N</FONT></SPAN><SPAN >��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">aminopeptidase N, APN</FONT></SPAN><SPAN >����ճ���ף�</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">cadherin</FONT></SPAN><SPAN >���Ǵ������۳�Ŀ�����г�ˢ״ԵĤ�ң�</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">brush border membrane vesicles, BBMV</FONT></SPAN><SPAN >����</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Bt</FONT></SPAN><SPAN >����</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Cry<?xml:namespace prefix = st1 /><st1:chmetcnv UnitName="a" SourceValue="1" HasSpace="False" Negative="False" NumberType="1" TCSC="0" w:st="on">1A</st1:chmetcnv></FONT></SPAN><SPAN >�����塣��ʵ�齫�����</SPAN><I ><SPAN lang=EN-US><FONT face="Times New Roman">Helicoverpa armigera</FONT></SPAN></I><SPAN >����ø</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">N1</FONT></SPAN><SPAN >����</SPAN><I ><SPAN lang=EN-US><FONT face="Times New Roman">Haapn1</FONT></SPAN></I><SPAN >��ճ������</SPAN><I ><SPAN lang=EN-US><FONT face="Times New Roman">Ha_BtR</FONT></SPAN></I><SPAN >˫��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">RNA</FONT></SPAN><SPAN >��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">dsRNA</FONT></SPAN><SPAN >��ע�������</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">4</FONT></SPAN><SPAN >���׳�����</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">, </FONT></SPAN><SPAN >���о���������������Ĭ���</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Cry<st1:chmetcnv UnitName="ac" SourceValue="1" HasSpace="False" Negative="False" NumberType="1" TCSC="0" w:st="on">1Ac</st1:chmetcnv></FONT></SPAN><SPAN >������Ӱ�졣�������</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">: </FONT></SPAN><SPAN >ע��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">dsRNA</FONT></SPAN><SPAN >��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">1 </FONT></SPAN><SPAN >��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">g/</FONT></SPAN><SPAN >ͷ�����л����Ĭ��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">, <I >Haapn1</I> mRNA</FONT></SPAN><SPAN >��������ע�仺��Һ��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">elution solution, ES</FONT></SPAN><SPAN >���Ķ����½���</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">30%~49%, <I >Ha_BtR</I> mRNA</FONT></SPAN><SPAN >�������½���</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">30%~37%</FONT></SPAN><SPAN >��ע��</SPAN><FONT face="Times New Roman"><I ><SPAN lang=EN-US>Haapn1</SPAN></I><SPAN lang=EN-US> dsRNA</SPAN></FONT><SPAN >���׳���</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">40</FONT></SPAN><SPAN >��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">70 ��g/cm<SUP>2</SUP> Cry<st1:chmetcnv UnitName="ac" SourceValue="1" HasSpace="False" Negative="False" NumberType="1" TCSC="0" w:st="on">1Ac</st1:chmetcnv></FONT></SPAN><SPAN >������µ���������������ע��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">ES</FONT></SPAN><SPAN >���׳�</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">, </FONT></SPAN><SPAN >����</SPAN><SPAN lang=EN-US><FONT face="Times New Roman"> 100 </FONT></SPAN><SPAN >��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman"> 170 ��g/cm<SUP>2</SUP> Cry<st1:chmetcnv UnitName="ac" SourceValue="1" HasSpace="False" Negative="False" NumberType="1" TCSC="0" w:st="on">1Ac</st1:chmetcnv></FONT></SPAN><SPAN >ԭ���ش�������������������������</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">; Cry<st1:chmetcnv UnitName="ac" SourceValue="1" HasSpace="False" Negative="False" NumberType="1" TCSC="0" w:st="on">1Ac</st1:chmetcnv></FONT></SPAN><SPAN >������Լ�ԭ���ض�ע��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Ha_BtR dsRNA</FONT></SPAN><SPAN >�׳���ע��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">ES</FONT></SPAN><SPAN >�׳�Ķ��������������졣��ͬʱע��</SPAN><I ><SPAN lang=EN-US><FONT face="Times New Roman">Haapn1</FONT></SPAN></I><SPAN >��</SPAN><FONT face="Times New Roman"><I ><SPAN lang=EN-US>Ha_BtR</SPAN></I><SPAN lang=EN-US> dsRNA</SPAN></FONT><SPAN >��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">, </FONT></SPAN><SPAN >���ź���׳��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Cry<st1:chmetcnv UnitName="ac" SourceValue="1" HasSpace="False" Negative="False" NumberType="1" TCSC="0" w:st="on">1Ac</st1:chmetcnv></FONT></SPAN><SPAN >����غ�ԭ���ص������Ծ������½������о���һ��֤���������</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Haapn1</FONT></SPAN><SPAN >��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Ha_BtR</FONT></SPAN><SPAN >����</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Bt</FONT></SPAN><SPAN >����</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Cry<st1:chmetcnv UnitName="ac" SourceValue="1" HasSpace="False" Negative="False" NumberType="1" TCSC="0" w:st="on">1Ac</st1:chmetcnv></FONT></SPAN><SPAN >�Ĺ�������</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">, </FONT></SPAN><SPAN >���������嵰��ͬ����</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Cry<st1:chmetcnv UnitName="ac" SourceValue="1" HasSpace="False" Negative="False" NumberType="1" TCSC="0" w:st="on">1Ac</st1:chmetcnv></FONT></SPAN><SPAN >�Ķ�ɱ���ù��̡��ý��Ҳ��ʾ</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">, <I >Haapn1</I></FONT></SPAN><SPAN >��</SPAN><I ><SPAN lang=EN-US><FONT face="Times New Roman">Ha_BtR</FONT></SPAN></I><SPAN >�������ͻ�䶼���ܵ���������</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Cry<st1:chmetcnv UnitName="ac" SourceValue="1" HasSpace="False" Negative="False" NumberType="1" TCSC="0" w:st="on">1Ac</st1:chmetcnv></FONT></SPAN><SPAN >�������ԡ�</SPAN></FONT></P>
Magsci [��������: 1]
<P class=MsoNormal ><FONT size=3><SPAN >����ø</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">N</FONT></SPAN><SPAN >��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">aminopeptidase N, APN</FONT></SPAN><SPAN >����ճ���ף�</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">cadherin</FONT></SPAN><SPAN >���Ǵ������۳�Ŀ�����г�ˢ״ԵĤ�ң�</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">brush border membrane vesicles, BBMV</FONT></SPAN><SPAN >����</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Bt</FONT></SPAN><SPAN >����</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Cry<?xml:namespace prefix = st1 /><st1:chmetcnv UnitName="a" SourceValue="1" HasSpace="False" Negative="False" NumberType="1" TCSC="0" w:st="on">1A</st1:chmetcnv></FONT></SPAN><SPAN >�����塣��ʵ�齫�����</SPAN><I ><SPAN lang=EN-US><FONT face="Times New Roman">Helicoverpa armigera</FONT></SPAN></I><SPAN >����ø</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">N1</FONT></SPAN><SPAN >����</SPAN><I ><SPAN lang=EN-US><FONT face="Times New Roman">Haapn1</FONT></SPAN></I><SPAN >��ճ������</SPAN><I ><SPAN lang=EN-US><FONT face="Times New Roman">Ha_BtR</FONT></SPAN></I><SPAN >˫��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">RNA</FONT></SPAN><SPAN >��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">dsRNA</FONT></SPAN><SPAN >��ע�������</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">4</FONT></SPAN><SPAN >���׳�����</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">, </FONT></SPAN><SPAN >���о���������������Ĭ���</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Cry<st1:chmetcnv UnitName="ac" SourceValue="1" HasSpace="False" Negative="False" NumberType="1" TCSC="0" w:st="on">1Ac</st1:chmetcnv></FONT></SPAN><SPAN >������Ӱ�졣�������</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">: </FONT></SPAN><SPAN >ע��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">dsRNA</FONT></SPAN><SPAN >��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">1 </FONT></SPAN><SPAN >��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">g/</FONT></SPAN><SPAN >ͷ�����л����Ĭ��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">, <I >Haapn1</I> mRNA</FONT></SPAN><SPAN >��������ע�仺��Һ��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">elution solution, ES</FONT></SPAN><SPAN >���Ķ����½���</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">30%~49%, <I >Ha_BtR</I> mRNA</FONT></SPAN><SPAN >�������½���</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">30%~37%</FONT></SPAN><SPAN >��ע��</SPAN><FONT face="Times New Roman"><I ><SPAN lang=EN-US>Haapn1</SPAN></I><SPAN lang=EN-US> dsRNA</SPAN></FONT><SPAN >���׳���</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">40</FONT></SPAN><SPAN >��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">70 ��g/cm<SUP>2</SUP> Cry<st1:chmetcnv UnitName="ac" SourceValue="1" HasSpace="False" Negative="False" NumberType="1" TCSC="0" w:st="on">1Ac</st1:chmetcnv></FONT></SPAN><SPAN >������µ���������������ע��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">ES</FONT></SPAN><SPAN >���׳�</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">, </FONT></SPAN><SPAN >����</SPAN><SPAN lang=EN-US><FONT face="Times New Roman"> 100 </FONT></SPAN><SPAN >��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman"> 170 ��g/cm<SUP>2</SUP> Cry<st1:chmetcnv UnitName="ac" SourceValue="1" HasSpace="False" Negative="False" NumberType="1" TCSC="0" w:st="on">1Ac</st1:chmetcnv></FONT></SPAN><SPAN >ԭ���ش�������������������������</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">; Cry<st1:chmetcnv UnitName="ac" SourceValue="1" HasSpace="False" Negative="False" NumberType="1" TCSC="0" w:st="on">1Ac</st1:chmetcnv></FONT></SPAN><SPAN >������Լ�ԭ���ض�ע��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Ha_BtR dsRNA</FONT></SPAN><SPAN >�׳���ע��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">ES</FONT></SPAN><SPAN >�׳�Ķ��������������졣��ͬʱע��</SPAN><I ><SPAN lang=EN-US><FONT face="Times New Roman">Haapn1</FONT></SPAN></I><SPAN >��</SPAN><FONT face="Times New Roman"><I ><SPAN lang=EN-US>Ha_BtR</SPAN></I><SPAN lang=EN-US> dsRNA</SPAN></FONT><SPAN >��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">, </FONT></SPAN><SPAN >���ź���׳��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Cry<st1:chmetcnv UnitName="ac" SourceValue="1" HasSpace="False" Negative="False" NumberType="1" TCSC="0" w:st="on">1Ac</st1:chmetcnv></FONT></SPAN><SPAN >����غ�ԭ���ص������Ծ������½������о���һ��֤���������</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Haapn1</FONT></SPAN><SPAN >��</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Ha_BtR</FONT></SPAN><SPAN >����</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Bt</FONT></SPAN><SPAN >����</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Cry<st1:chmetcnv UnitName="ac" SourceValue="1" HasSpace="False" Negative="False" NumberType="1" TCSC="0" w:st="on">1Ac</st1:chmetcnv></FONT></SPAN><SPAN >�Ĺ�������</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">, </FONT></SPAN><SPAN >���������嵰��ͬ����</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Cry<st1:chmetcnv UnitName="ac" SourceValue="1" HasSpace="False" Negative="False" NumberType="1" TCSC="0" w:st="on">1Ac</st1:chmetcnv></FONT></SPAN><SPAN >�Ķ�ɱ���ù��̡��ý��Ҳ��ʾ</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">, <I >Haapn1</I></FONT></SPAN><SPAN >��</SPAN><I ><SPAN lang=EN-US><FONT face="Times New Roman">Ha_BtR</FONT></SPAN></I><SPAN >�������ͻ�䶼���ܵ���������</SPAN><SPAN lang=EN-US><FONT face="Times New Roman">Cry<st1:chmetcnv UnitName="ac" SourceValue="1" HasSpace="False" Negative="False" NumberType="1" TCSC="0" w:st="on">1Ac</st1:chmetcnv></FONT></SPAN><SPAN >�������ԡ�</SPAN></FONT></P>
[��������: 1]
[��������: 1]
[��������: 2]
[��������: 2]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 3]
[��������: 1]
[��������: 1]
[��������: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}