 ,�й�ˮ���о���/ˮ������ѧ�����ص�ʵ����,���� 311400
,�й�ˮ���о���/ˮ������ѧ�����ص�ʵ����,���� 311400Effects of Various Paddy-Upland Crop Rotations and Nitrogen Fertilizer Levels on CH4 Emission in the Middle and Lower Reaches of the Yangtze River
LIU ShaoWen, YIN Min, CHU Guang, XU ChunMei, WANG DanYing, ZHANG XiuFu, CHEN Song,China National Rice Research Institute/State Key Laboratory of Rice Biology, Hangzhou 311400ͨѶ����:
���α༭: ����ϼ
�ո�����:2019-01-11��������:2019-02-25�����������:2019-07-16
| ��������: |
Received:2019-01-11Accepted:2019-02-25Online:2019-07-16
����� About authors
������,E-mail: 990155952@qq.com��

ժҪ
�ؼ��ʣ�
Abstract
Keywords��
PDF (554KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر���
�������ø�ʽ
������, ����, �ҹ�, �촺÷, ����Ӣ, ���㸣, ����. ���������ε�����ͬˮ������ģʽ�͵���ˮƽ �Ե���CH4�ŷŵ�Ӱ��[J]. �й�ũҵ��ѧ, 2019, 52(14): 2484-2499 doi:10.3864/j.issn.0578-1752.2019.14.008
LIU ShaoWen, YIN Min, CHU Guang, XU ChunMei, WANG DanYing, ZHANG XiuFu, CHEN Song.
0 ����
���о����塿ȫ�������ů������Ӱ������������滷��������[1],���䷢������������Ũ�ȵ������й�[2]��CH4��Ϊ��Ҫ����������֮һ,������ЧӦԼռȫ������������ЧӦ��15%��20%,��ÿ����1.1%���ٶ�����[3]���о�����ũ���Ǵ���CH4����Ҫ�ŷ�Դ,Լ17.9%��CH4�ŷ���Դ�ڵ���[4]��ˮ�����й���Ҫ����ʳ����,����ֲ���Լռ����ˮ����ֲ�������23%[5]�������о�����CH4�ŷ��ص㼰��Ӱ������,���ڴ˻������ƶ���Ӧ�ļ��Ŵ�ʩ,�Ի���ȫ������ЧӦ�����ش�ǰ���о���չ������CH4�IJ������ɲ���������������л�����ͨ����������ת������[6],CH4���������������ˮ���������������Ҫ���ɷ�����ɢ�����ݱŷ���ͨ��ֲ���ŷ�[7],���г���90%�ĵ���CH4ͨ��ˮ��ֲ���ŷŵ�����[8]���Ե���CH4�ŷż���Ӧ�ļ��Ŵ�ʩ���д����о��������������������ԡ�������ʽ�Լ���������Ӱ�쵾��CH4�ŷŵ���Ҫ����[9,10,11]��һ����Ϊˮ�������ܹ�������������������,����������ԭ���ʺ���,����CH4�IJ���[12];�����������ˮ,���谭��������ɢ,��������������ԭ��λ,���ڲ�������Ŀ��ٷ�ֳ,����CH4�ŷŵ�����[13];���ڿ������ܼ���36%��65%���ϵ�CH4�ŷ�[14,15]������,�о�����,�ո��������������ӵ���CH4�ŷ���,���ŷ������ͽոѻ������������[16]����������,ˮ���������������������Ŷ�������ո�ȫ������,���������ε�����Ҫ��ˮ���������������͡����̡����е�ģʽ[17,18],��������,�ոѸ�����ֲ������Ҳ���㷺�ƹ�[19]�������Ľոѻ��������������л��ʺ���,��ʵ��������������ṹ[20],����ͬ���ԵĽոѻ�������������ŷŵ�Ӱ��,Ŀǰ��δ��ϵͳ����������о�����,��䵪��ʩ�öԵ���CH4�ŷ���������Ӱ��,��Ŀǰ��δ��һ�µĽ���,���о���Ϊ���ʻ����ӵ���CH4�ŷ�[21,22],Ҳ���о���������ʩ����������,����CH4�ŷż���[23]�����ڵ��ʻ�Ӱ�컹��ոѵķֽ⼰��CH4ת������[24],��˵��ʶ�CH4�ŷŵ�Ӱ����ܻ��뵾��Ľո������йء������о�����㡿Ŀǰ����ˮ������ģʽ����ͬ���Խոѻ��ﴦ�����뵪��ˮƽ�Ե���CH4���ŷ����ԵĻ���ЧӦ�о���������,����������ŷŻ������о�����δ���������������Ĺؼ����⡿����,���о��Զ��곤�ڶ�λ���飨2003����Ϊ����,ѡ��4�ֲ�ͬˮ������ģʽ����ͬ���Խոѻ��ﴦ����,���3�ֲ�ͬ����ˮƽ,ͨ���������ɼ���������Ʒ����,��ҪĿ������:��1����������ո������뵪��ˮƽ�����Ե���CH4���ŷŹ��ɼ��ۻ��ŷ�����Ӱ��;��2��������������л�����������������ո������ۺϷ���,��ȷ��ͬ�ոѻ����뵪�ʴ����µ���CH4�ŷŲ������������Ϊ�ҹ��������������ŷ��Լ��������Ŵ�ʩ���ƶ��ṩ��ѧ���ݡ�1 �����뷽��
1.1 �������ſ�
������2016��2017��,���㽭ʡ�������й�ˮ���о���ũ�����У�120.2��E,30.3��N,����11 m������ũ��λ�ڳ���������ƽԭ,�����ȴ���������,��ƽ������Ϊ13��20��,����7�·�Ϊ35��;��ƽ����ˮ��Ϊ1200��1600 mm,����4�·ݺ�9�·ݽ�����Լռ80%�����о�����ĵ�����ֲģʽ��λ������2003�꿪ʼʵʩ,��2016��2017��ⶨ����CH4�ŷ�,������������������ȼ���첻����,����ָ�����1��Table 1
��1
��1��ͬ����������ˮƽ�������������ʣ�����ƽ����
Table 1
| ���� Rotation | ���� Fertilizer | TN (g��kg-1) | AN (g��kg-1) | SOM (g��kg-1) | pH |
|---|---|---|---|---|---|
| RF | N0 | 2.52g | 0.14fc | 31.2e | 6.18ab |
| N1 | 3.27cd | 0.17e | 39. 7bcd | 5.87bc | |
| N2 | 2.91ef | 0.18cd | 39.8cd | 5.76c | |
| RC-G | N0 | 2.77f | 0.19cd | 37.8d | 6.27a |
| N1 | 2.87ef | 0.19cd | 38.1cd | 6.12abc | |
| N2 | 3.48abc | 0.19bc | 41.9b | 5.84bc | |
| RW | N0 | 3.10de | 0.17de | 39.7bcd | 5.89bc |
| N1 | 3.29bcd | 0.20bc | 40.8bc | 5.80c | |
| N2 | 3.24cd | 0.19cd | 38.3bcd | 5.78c | |
| RP | N0 | 3.40abc | 0.21b | 45.0a | 5.83bc |
| N1 | 3.52ab | 0.25a | 46.1a | 5.86bc | |
| N2 | 3.58a | 0.19bc | 46.4a | 5.84bc |
�´��ڴ�|����CSV
1.2 �������
���о��Բ�ͬˮ������ģʽ���ڶ�λ����Ϊ������2003������2016��2017��ⶨ����CH4�ŷš���λ���������������������,4�ֲ�ͬ����ģʽΪ����,�ֱ�Ϊˮ��-���У�RF��,ˮ��-����Ӣ��RC-G��,ˮ��-С��RW����ˮ��-���ݸ�����ֲ��������RP��;ˮ��������ˮƽΪ����,�ֱ�ΪN0��0����N1��142.5 kg N��hm-2����N2��202.5 kg N��hm-2��,С��λ�ú�������ڹ̶�,ÿ��С����8 m����8 m,3����������ظ���������������:RF,��䲻���κδ���,���з���,����ˮ������ǰ,��������;RC-G,����Ӣ��10������Ѯ����,�����ʩ������15 kg��hm-2��Ϊ����,����ʢ���ڣ�3�µ�4�³�������,��������;RW,С����10����Ѯ11�³���ֱ��,���ںͳ����ڷֱ�ʩ�븴�Ϸ�75 kg��hm-2�����ʣ�������15 kg��hm-2���ʣ�,����5����Ѯ�ջ�,��������,����ոѷ�������;RP,����ȫ�̸�����ֲ������,��1����Ѯ��������ֱ�Ӱڷ�����,������8��10 cm��,ʩ�븴�Ϸ�75 kg��hm-2��Ϊ����,5����Ѯ���������龥,���ಿ�ַ�������,������л���Ϊ���õĵ��ݺ���������Ҷ�ȡ�
��������ֲ:����Ʒ��Ϊ��58�����澬������ˮ������6����Ѯ������ˮ�����ִ�ѿ��,5������Ѯ��������,��3Ҷ1����������,�����26.7 cm��13.3 cm��ˮ����ʩ����,�����������ݵ��ʴ�������,���л���:������:��ʰ���40%:30%:30%ʩ��;�ʣ��������,60 kg��hm-2����Ϊ��һ����ʩ��;�طʣ��Ȼ���,135 kg hm-2�����������ʸ�50%�ִ�ʩ�롣
�ոѻ��ﴦ��:ˮ���˹��ջ�,��ո�ȫ���Ƴ����ڶ����������������ࡣ��������ո�ȫ������,��ͬ����ģʽ�½ոѻ�����������ո�C/N����ͼ1��ʾ����ͬ����ģʽ�½ոѻ���������������ͼ1-A,��2017��RW��RC-G���첻������,��RP���,RW��RC-G�Ľոѻ������ֱ����45.7%��78.3%��2016��,51.6%��62.5%��2017�������ڻ���ոѵ�C/N�ȣ�ͼ1-B������ΪRW��������RP��RC-G��RP��RC-G���첻������;�����RW,RP��RC-G������C/N�ֱ�58.0%��71.3%��2016��,58.9%��74.1%��2017�������,���ݽո�����,���Խ�4������ģʽ����ΪRP���߽ո�������C/N�ȣ���RW���߽ո�������C/N�ȣ���RC-G���ͽո�������C/N�ȣ���RF������-�ոѣ���
ͼ1
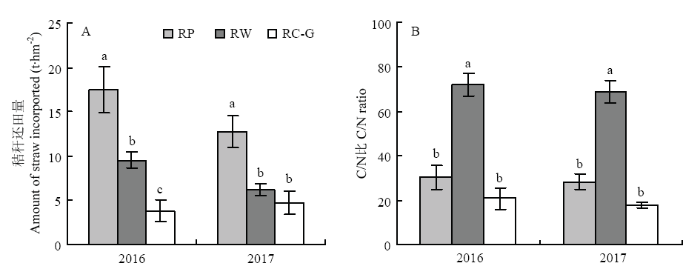 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ1��ͬ����ģʽ�½ոѻ�������A���ͻ���ո�C/N�ȣ�B��
���ϲ�ͬСд��ĸ��ʾ��ͬ����ģʽ�����������P<0.05��
Fig. 1Amount (A) and C/N ratio (B) of straw incorporated under different rotations
Different small letters in same year indicated significant difference among different rotation systems at P<0.05
1.3 ��Ʒ�ɼ������
CH4����ɼ���2016��2017��6��10�¼俪չ,�����ܱ�ʽ��̬��������-����ɫ���ⶨ���CH4�ŷ�ͨ��������������ɵ����Ͳ��������,��̬�������Ϊ0.5 m��0.6 m,��������PVC�����Ƴ�,����߶���ˮ���߶ȶ����ӣ��ν�ǰ 0.5 m���νں� 1.2 m��,�в��̶�����,����Ƕ���¶ȼ�,�����¼�¶ȱ仯,����Ϊ������U��,Ƕ��ر�,����������ˮʹ֮�ܷ⡣�ɼ�Ƶ��:ˮ�������Ժ��3����,����ʵ�����ÿ5��10�����1�Ρ�����ʱ��Ϊ8:00��11:00������ɼ�ǰ�����ڶ���12 VС�����Գ�ֻ�����������,����ʱ,�������䴹ֱ�����ڵ���5 cm��İ����ڲ���ˮ�ܷ�,��Ʒ�ɼ��ֱ��ڹ�����0��6��12��24 min���С���������CH4Ũ��������ɫ�ף�Agilent 7890A,���������Ƽ�����˾�������ⶨ��CH4�������FID����������ӻ��������,����¶�300��,���� 60��,����Ϊ 99.999%�ߴ� N2,���� 30 mL��min-1��
�������ո����Բⶨ:�л��ʺ�������ˮ�����ظ��������-��ɫ���ⶨ[67];ȫ����ȫ�Զ����϶����ǣ�����K9840,���Ϻ��������ɷ�����˾���ⶨ;�����ü����ɢ�����вⶨ[67];����pH��pH�ƣ�F2,÷����-�������������Ϻ�������˾�����вⶨ��
����������鴦��,��ȼ����һ��,�Ҿ������궨λ����,�������ʻ����ȶ�,�����2018������ڲⶨ����������ԭ��λ������mcrA��pmoA��������̽��CH4�ŷŻ������졣
����������ԭ��λ��Eh���ⶨ��2018�꣩:��ˮ�����Ժ�ÿ5��7��ⶨһ�Ρ���������������ԭ��λ�ǣ�FJA-6��,�Ͼ����������豸����˾���ⶨ���������е�Ehֵ��
����mcrA��pmoA�����Ȳⶨ:�ֱ���ˮ�����Ժ�3 d��2018/6/28����14 d��2018/7/9����28 d��2018/7/23������ȡ�����ɼ�ÿ��С���ൾ20 cm,���㣨0��20 cm��������Ʒ��5��ȡ����ȡ����,ȥ���ʺ��ϴ��á�����mcrA��pmoA�����Ȳ���ʵʱӫ�ⶨ��PCR��������������DNA������ȡ�Լ�����ȡ����DNA,��0.8%����֬��������Ӿ�������DNAƬ�δ�С,���ó�������ֹ��ȼƲⶨDNAŨ�ȼ���������Ʋ��������mcrA��������������pmoA��������,����ʵʱӫ�ⶨ��PCR����,��Ӧ��ϵΪ20 ��L[25]����������PCR�������õ�����ͷ�Ӧ��������2,���вɼ��ź��¶�Ϊ72�档
Table 2
��2
��2��ʵʱ����PCR�������P��Ӧ����
Table 2
| Ŀ�Ļ��� Target gene | ���� Primer | ��������(5���3��) Sequence (5���3��) | ����PCR��Ӧ���� Thermal profile |
|---|---|---|---|
| mcrA[26] | MLf | GGTGGTGTMGGATTCACACARTAYGCWACAGC | 94��Ԥ����3 min,94�����25 s,50���˻�45 s,72������60 s,35��ѭ�� Pre-denaturation at 94�� for 3 min, denaturation at 94�� for 25 s, annealing at 50�� for 45 s, extension at 72��C for 60 s, 35 cycles |
| MLr | TTCATTGCRTAGTTWGGRTAGTT | ||
| pmoA[27] | PmoA A189f | GGNGACTGGGACTTCTGG | 95��Ԥ����5 min,92�����1 min,55���˻�1.5 min,72������60 s,35��ѭ�� Pre-denaturation at 95�� for 5 min, denaturation at 92�� for 1 min, annealing at 55�� for 1.5 min, extension at 72�� for 60 s, 35 cycles |
| pmoA mb661r | CCGGMGCAACGTCYTTACC |
�´��ڴ�|����CSV
1.4 ���ݴ���
���������ŷ�ͨ���ɸ�����������Ũ����ʱ��ı仯�ʼ���ó�,���㷽��Ϊ�����ŷ�ͨ�����㹫ʽ����:$F=\frac{dc}{dt}\cdot{h}\cdot{p}\cdot\frac{273}{273+T}$
ʽ��,F Ϊ�ŷ�ͨ����mg��m-2��h-1��;dc/dtΪ������������������Ũ����ʱ��ı仯�ʣ�mL��m-3��h-1)��;hΪ��������߶ȣ�m��;�� Ϊ��״̬��������ܶ� ��0.714 kg��m-3��;TΪ����ʱ�����¶ȣ��棩,273Ϊ��̬���̳�����ÿ�ι۲�ʱ�����������ŷ�ͨ����3���ظ���ƽ��ֵ��ʾ;����������������ƽ���ŷ�ͨ���ļ��������������β���ʱ����ΪȨ��,�������ŷ�ͨ�����м�Ȩƽ��;�����ۻ��ŷ�����ƽ���ŷ�ͨ����ȫ������ʱ��ij˻���ʾ��
���IJ���Excel 2010��SAS 9.2���������ݽ�����м���ͷ���,����Duncan���������ݼ�IJ��������ԣ�P=0.05����
2 ���
2.1 ����CH4�ŷ�ͨ��
ˮ�������ڵ���CH4�ŷ�ͨ���ı仯��ˮ�ֹ���Ӱ������,�������ԵĽ��Ա仯��ͼ2��,�ɹ۲�����ŷŸ߷��ڡ������ں�������CH4�ŷŽΡ����з�����CH4�ŷŽΣ�Լ���Ժ�7��30 d������ʱ��ռȫ�����ڵ�19.0%��19.7%�������к��ڣ����Ժ�37 d���ң�,�������������,������CH4�ŷ�ͨ�������ֲ�ͬ�̶��½������︴ˮ��,CH4�ŷ�С������,����������CH4�ŷŽΣ�Լ���Ժ�50��70 d��,�ýγ���ʱ��Լռ�����ڵ�16.7%�� 17.2%��֮�������ŷ�ͨ�����ڡ�ͼ2
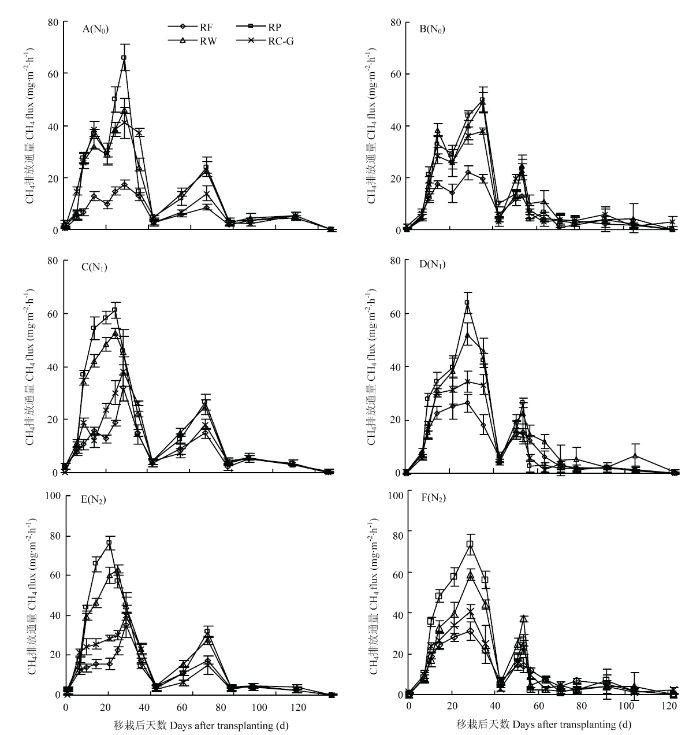 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ22016��A��C��E����2017��B��D��F����Ȳ�ͬ����ģʽ�������ݶ��µ���CH4�ŷ�ͨ���ļ��ڱ仯
Fig. 2Seasonal variations of CH4 fluxes in rice season under different rotation systems and N fertilizer rates in 2016 (A, C, E), 2017 (B, D, F)
����ģʽ�Ե���CH4�ŷŵ�Ӱ����ˮƽ�����������ͬ��N0ˮƽ��,������CH4ƽ���ŷ�ͨ��,RP��RW��RC-G����������,������������RF,�ֱ����72.1%��168.7%��70.8%��145.2%��56.5%�� 145.2%��ͼ2- A ��2-B���������CH4�ŷ�ͨ��,��ģʽ������������,��2016��RP������������������,N0ˮƽ�·�����CH4�ŷű���Ϊ˫���ص㣨ԼΪ���Ժ��12��15 d��25��30 d���������������Ժ�25��30 d��������ŷŷ�,RP��RW��RC-G������ŷ�ͨ��������ƽ�����ֱ�Ϊ55.0��42.9��38.6 mg��m-2��h-1�����RF,RP��RW��RC-G������ŷ�ͨ���ֱ����182.1%��120.0%��97.9%��
����ʩ����������,CH4�ŷ�ͨ������������Ӧ���ַֻ���RF��RP��RW�ķ����ں�������CH4ƽ���ŷ�ͨ������ʩ���������Ӷ�������ߡ�N2ˮƽ��,RP������CH4ƽ���ŷ�ͨ��Ϊ36.2 mg��m-2��h-1,Ҫ��N1��N0�ֱ��14.8%��40.4%;��RP����,RW��Ҫ��13.9%��43.9%��RF�����Ϊ������CH4�ŷ�ͨ��,N1��N2��������N0,���������Ժ�25��30 d,��ƽ���ŷ�ͨ���ﵽ23.8��27.5 mg��m-2��h-1������ƽ����,���N0,�ֱ����29.9%��50.3%����RC-G������CH4�ŷ�ͨ����������ȶ���2017���������½���2016������N0���,��ʩ������RC-G������CH4ƽ���ŷ�ͨ������20.0%��29.8%��N1����11.9%�� 20.1%��N2����
2.2 ����CH4�ۻ��ŷ���
��ͼ3�п��Կ���,��ͬ����ģʽ������ˮƽ�Ե���CH4�ŷ�����Ӱ���������������N0������,���У�RF��ģʽ�ĵ���CH4�ۻ��ŷ���Ҫ�������ڶ���������ֲ��RP��RW��RC-G����RF�������CH4�ۻ��ŷŽ�18.4��21.2 g��m-2,�ֱ��RP��RW��RC-G����42.6%��59.4%��40.7%��52.1%��36.0%�� 52.1%��ͼ3
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ32016��A����2017�꣨B����ͬ����ģʽ�������ݶ��µ���CH4�ۻ��ŷ���
ͬһ��ݲ�ͬСд��ĸ��ʾ��ͬ����ģʽ�����ϼ����������P<0.05��
Fig. 3CH4 accumulation emissions in rice season under different rotations and N fertilizer rates in 2016 (A) and 2017 (B)
Different small letters in the same year indicated significant difference among different rotation systems and fertilizers at P<0.05
���ŵ��ʵ�����,RP��RW��RF�����µ���CH4�ۻ��ŷ������в�ͬ�̶ȵ����ӡ�N2ˮƽ��,RP��RW��RF��CH4�ۻ��ŷ����ֱ�Ϊ51.2��55.8��45.3��51.5��25.0��30.5 g��m-2,��N0ˮƽ���23.0%��38.4%��26.7%��33.7%��35.3%��43.5%;����N1���,�����9.9%��19.7%��20.8%��23.1%��17.4%��18.8%��2016,RFģʽ��,N1��N2���������죩����RC-G����CH4�ۻ��ŷ���������ʩ���������Ӷ��������͡���N0���,N1��N2�µ���CH4�ۻ��ŷ����ֱ�20.7%��42.4%��10.6%��16.6%����ͬ������Ƚ�,N1��N2ˮƽ��,RF��RC-G�ĵ���CH4�ۻ��ŷ�������������,����������RW��RP��
����CH4�ŷ�ͨ������ˮ����������ء�Ϊ�˽�һ����ȷ��ͬ����ģʽ�뵪��ˮƽ�µ���CH4���ŷŹ���,��ˮ��ȫ�����ڷ�Ϊ�����ڡ������ںཬ��3����Ҫ�����Ρ��������,����CH4���ۻ��ŷ���������>������>�ཬ��,���з����ڡ������ںཬ��ռ�ȷֱ�Ϊ51.85%��72.31%��22.9%��31.3%��6.0%��20.7%,�����CH4�ŷ���Ҫ����������ǰ����,������ռ�Ƚ�С��ͼ4��������ģʽ�͵���ˮƽ��ˮ����ͬ������CH4�ۻ��ŷ���Ӱ������,������Ҫ���������ڷ����ں�������,���ཬ�������������졣N0ˮƽ��,��RP��RW��RC-G���,RF�ķ�����CH4�ۻ��ŷ�������95.9%��171.1%��70.7%��144.1%��74.2%��165.8%;���ŵ��ʵ�ʩ��,������������CH4�ۻ��ŷ���ռ�Ⱦ�������ߣ�2016��RC-G���⣩,����Ϊ7.0%��48.3%��������CH4�ۻ��ŷ���,RP��RWģʽ��������RC-G��RF,��RC-G��RF���������졣�ཬ��,CH4�ŷ�����С,������֮�����������졣
ͼ4
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ42016��A��C��E����2017��B��D��F����Ȳ�ͬ����ģʽ�������ݶ���ˮ����ͬ������CH4�ۻ��ŷ���
��͢�ֱ���������ڡ������ںཬ�ڡ�ͬһ�����β�ͬСд��ĸ��ʾ��ͬ����ģʽ�����������P<0.05��
Fig. 4CH4 accumulation emissions at different rice growth periods under different rotation systems and N fertilizer rates in 2016 (A, C and E) and 2017 (B, D and F)
��, �� and �� represent tillering stage, booting stage and filling stage; respectively. Different small letters in the same growth period indicated significant difference among different rotation systems at P<0.05
2.3 ���������mcrA���ͼ�����������pmoA��������
�����в��������mcrA���ͼ�����������pmoA����Ӱ�쵾��CH4�ŷŵ���Ҫ���Ϊ��һ����ȷ��ͬ����ģʽ�µ���CH4�ŷ��������,��2018��ˮ�����Ժ�3��14��28 d�ɼ����ⶨ������mcrA��pmoA�����ȣ�ͼ5������ͬ����ģʽ�뵪��ˮƽ��,mcrA��pmoA�����Ȳ���������N0ˮƽ��,���Ժ�3 d,mcrA������������,������14 d��28 d�����������ӡ����Ժ�28 d mcrA�����ȴ�С˳��ΪRP>RW>RC-G>RF,�����RF,RP��RW��RC-Gģʽ��mcrA����������195.7%��121.3%��45.7%,��˵���ոѻ�����������������в��������ȡ�ʩ����,������ģʽmcrA�����Ⱦ���������,�����N0,N1ˮƽ��RP��RW��RC-G��RF��mcrA�����ȷֱ�����191.4%��135.6%��43.1%��105.7%�����Ժ�28 d��,������RC-G�������١�ͼ5
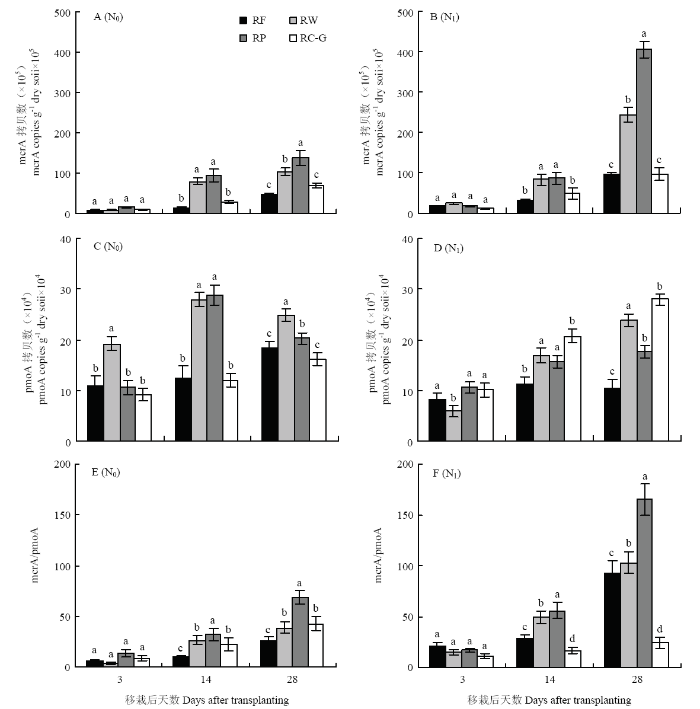 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ52018�겻ͬ����ģʽ�������ݶ��²����������mcrA;A��B���ͼ�������������pmoA;C��D����������mcrA/pmoA��ֵ��E��F��
ͬһ���Ժ�������ͬСд��ĸ��ʾ��ͬ����ģʽ�����������P<0.05��
Fig. 5Abundance of methanogenic gene mcrA (A and B) and methanotrophic gene pmoA (C and D) and mcrA/pmoA ratio (E and F) under different rotation systems and N rates in 2018
Different small letters in the same day after transplanting indicated significant difference among different rotation systems at P<0.05
��mcrA��ͬ,N0ˮƽ��,���Ժ�14��28 d,RP��RW��pmoA��������������RF��RC-G��������,RF��RW��RP��pmoA�������������ٽ����ֱ�Ϊ42.7%��14.1%��13.3%�����Ժ�28 d��,˵������һ���̶��������˼����������Ļ���,��RC-G��պ��෴,��ʩ���ʺ�pmoA����������72.8%�����Ժ�28 d����
���Ͻ������,���ŵ��ʵ���ʩ,RP��RW��RF�IJ�������������,��RC-G���ӽ���;�������������ķ�ȱ仯����CH4�ŷŸպ��෴,����ΪRP��RW��RF pmoA�����ȼ���,��RC-G���ӡ���һ������mcrA/pmoA��ֵ(ͼ6-E��6-F),����N0ˮƽ��,��ͬ����ģʽ��mcrA/pmoA��ֵ��С˳��ΪRP>RW��RC-G>RF�����Ժ�14 d��28 d��,��仯�����뵾����ͬ������ʽ��CH4�ŷű仯����һ�¡���ʩ���ʺ�RF��RP��RW mcrA/pmoA�����ȱ�ֵ�������,�����ӱ����ֱ�Ϊ71.4%��141.1%��197.1%��258.2%��84.6%��165.5%;��RC-G���½�26.8%��42.3%���������CH4���ŷŹ������ơ�
ͼ6
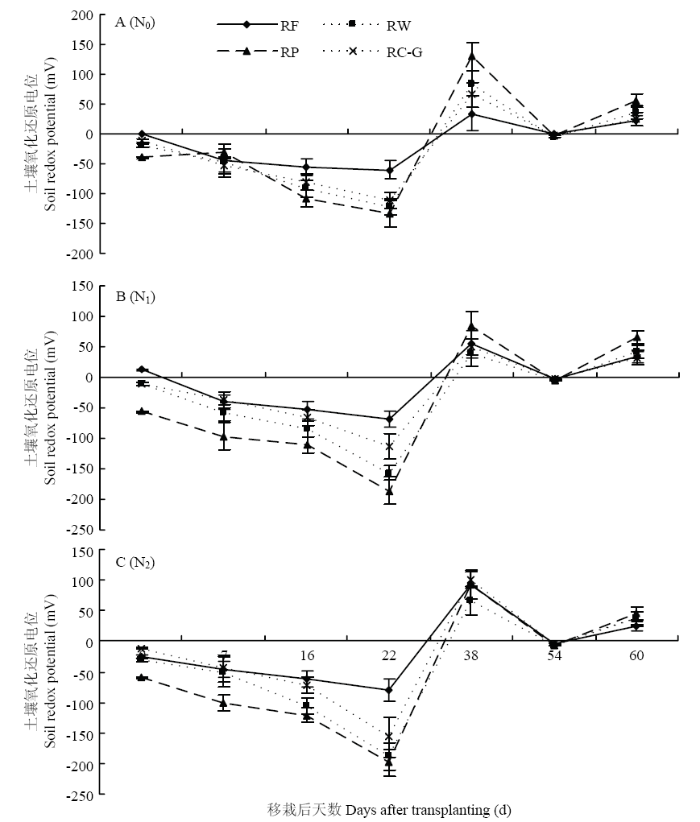 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ6��ͬ����ģʽ�������ݶ��µ�������������ԭ��λ�仯����
Fig. 6Variations of soil redox potential in rice season under different rotation systems and N fertilizer rates
2.4 ������ԭ��λ������
��ͬ����ģʽ��������ԭ��λ��Eh���仯�����ڲ�ͬ���ʴ����»���һ�£�ͼ6��,����ǰ������һֱ������ˮ״̬��,������ģʽEh�����½�,�����Ժ�18��25 d�������,Ȼ��ʼ����,����30��35 d�Ժ�Eh>0,���ʲ���ʽ�仯��N0ˮƽ��,ˮ������ǰ�ڣ�3��7 d��,��ʱ������ģʽEh��С˳��ΪRP>RW>RC-G>RF��������Eh���첻������,���������ڽոѵ��������е����������������ͨ���ԡ����Ժ�22 d,RP��RW��RC-Gģʽ֮������������,������������RFģʽ,����Ϊ118.4%��102.6%��80.5%,˵��������ˮʱ����ӳ�,�ոѵĽ��������������е�O2,������������������ԭ��λ�Ľ��͡���ʩ���ʺ�,Eh�仯����������ģʽ���������죨���Ժ�22 d��,N2ˮƽ��,RP��RW��RC-G��RF��Eh�ֱ�Ϊ-198.9��-188.5��-156.3��-80.3 mV��RP��RW��RC-Gģʽ֮������������,������������RFģʽ��,��N0���,�ֱ�33.1%��34.5%��29.7%��24.2%������N1���,��7.2%��20.2%��38.4%��15.9%�������ʵ�Ͷ��,һ���̶��ϴٽ��ոѵĸ�������,����һ������������������ԭ��λ�Ľ��͡�3 ����
3.1 ����ģʽ�Ե���CH4�ŷŵ�Ӱ��
���������뵪��ˮƽ��δ�ı䵾��CH4�ܵ��ŷŹ��ɡ������ˮ�ֹ�������������Ӱ��,����CH4�ŷ���Ҫ������ˮ��������,����ʱ�ڣ���νڳ����ڡ��ཬ��ʵ�ڣ���Խ���[28,29]����ˮ��������,CH4�ŷű���Ϊ������ǰ������,�к����ر��Ǹ��↑ʼ��,����������︴ˮ����������,�����ֵ��ŷ���ֱ��ˮ���ջ�,���ֲ���ʽ�仯[5]�����о���������ģʽ�뵪��ˮƽ�Ե���CH4��ͬ���ŷ�ͨ�������ۻ��ŷ�����������Ӱ��,��Ӱ����Ҫ�����ڷ�����,����������[30]�ͻ�̫��[31]�ȵIJ����о�����һ�¡�����,��Х�ֵ�[32]�о���Ϊ���ӽոѺ��̷���������ӵ���CH4�ŷ�,�����Ӹ��������ոѱ����ӵ��������̷������и����CH4�ŷš����뱾�о�ʩ����N1��N2�������²�ͬ����ģʽCH4�ŷŹ��ɻ���һ��;����N0��,RP��RW��RC-G���������졣��仯���ɻ���������������������ϵ,�����ӽո���������CH4�ŷ�֮��Ĺ�ϵ�ܵ���ʩ�õ���,���ܵ�����������Ե���Լ��������ֳ�뼤����Ҫ���������,��������Ч̼����Ӱ������������Ե���������[33]������CH4�ŷ���Ҫ�����ڲ�������ͼ����������ķ�ȼ�����ԡ���������ǵ��������м����������Ҫ����Դ[34]��������������,���Լ�����ΪΨһ̼Դ����Դ����ͬ�����컯��л������[35]���������[36]�о�ָ�������������뵾��CH4�ŷ����������,���뱾�о����һ�¡��ոѻ����뵪�ʴ���Ӱ�����л�̼��ת���Ͳ����������������������õ��л�̼������,����Ӱ�쵽CH4�ŷ�[37]������ֲϵͳ��,�ո�ȫ�������ܹ��������Ӳ���������,�������µ���CH4�ŷ������[38]�����о�����N0��,���������ȱ仯������ոѻ������й�,����BAO[38]��ARIF[39]�ȹ��ڴ����ո�̼��Ͷ�����ӵ��������������Ľ��һ�¡���һ����,����CH4�ŷ���Ҳ��ոѻ�����л�����������졣ǰ���о���Ϊ���ӵ��л�����C/NԽ��,��Դ����Խ�ḻ,�����й�������Ծ�Խ��,�ڻ������˵�������,CH4�ŷž�Խ��[40]�����о���RW��RP��RC-G����ոѵ�C/N�ȷֱ�Ϊ70.39��29.25��19.26��ͼ1 B��,����RP����C/N�ϵ�,����CH4�ŷ�ͨ����������RC-G,������ոѻ�������������RC-G�����ͼ1-A������Ȼ,�Ի���ո�C/N����Ϊ�жϻ���ոѶ�CH4�ŷŵ�Ӱ���ȫ�档������Ϊ��C/N���,����ո����л�̼����������CH4�ŷŹ�ϵ��������,����ЧӦ�ܵ���ʩ�õ�Ӱ�졣����,���ڶ�λ������,RP�����нϸߵ��л��ʺ�������Ҳ�ǵ���RP����ģʽCH4�ŷ���������Ҫ���أ���1����
3.2 ����ģʽ�͵���ˮƽ�Ե���CH4�ŷŵĻ���ЧӦ
�����ܹ�Ӱ�쵾��CH4�ŷ�,��ЧӦ�����ࡢʩ������ʩ�÷�ʽ��ʩ��ʱ���Ӱ��[23]�����ڻ���ոѵ�C/N���ܸ�����ʩ���ʴ�����CH4�ŷ�ЧӦ���ڱ��о���RP��RW��RF����ģʽ��,���Ƿ���,��ʩ�����ܹ��������ӵ���CH4�ŷţ�ͼ3��,��ǰ���о�ӡ֤[41]����RC-G�ı�����պ��෴������ZHU[42]�ͺ�������[5]��������Ӣ-ˮ������ģʽ��ͬʩ������CH4�ŷ��о�����һ�¡��������л����ϵĻ���ЧӦ�Ե���CH4�ŷŵ�Ӱ����Ҫ���л����Ϸֽ⼰������л��������й�[43]��һ�����,���нϸ�C/N�ȵ��л�����,��ֽ��������ŵ������������Ӷ�����[44]������ոѵķֽ����ʻ�������������������,�������ܵ���������Ч����������Լ[45],�о�����,�ڵ����Ļ�����,���ӵ���������������е�����Ч��,����Ӧ����������в�����������л��������Ч��,�ٽ��˲������������,����CH4����[46]�����о���һ����������Ե��ʵ���Ӧ����������ģʽ����������,��ʩ���ʴٽ������������ֳ,ȴ�����˼�������������ֳ��ͼ5-A,5-B��,����仯����������������������ͬ������������[25]���������п����õ��������̬������̬������mcrA����������ض���pmoA�����ȳ���������صĽ���һ�¡�������ΪCH4�ŷŶԵ��ʵ���Ӧ,ԭ�������:��1�����������Ҫ��NH4+-NΪ��Դ,��Դ��������������˲��������ֳ����������Ч��[46],��ӿ��������������[47],�Ӷ��ٽ���Ⱥ��Ŀ�������[48];��2�����鵥��ø��MMO���Ǽ����������Ĺؼ�����ø,�����������������´�����ȵ�̼������ϩ���ǻ���������[49],NH4+��MMO�ľ��������Ƽ�,ͨ����MMO�ľ���,�����˼�������,�ٽ��˼����ŷ�[50];��3���������ӻ�������ֲ���ϵ����������,����Ϊ����������������������[51];��4�����ڲ��������л�����ϸ�����,ֻ��������������ԭ��λ�ܵ͵Ļ�����[52]������������Ͷ��,�ܹ������л����ϵĸ�������,�����ͬʱ������ˮ�е�O2,�����������е�������ԭ��λ��ͼ6-B��6-C��,��һ�������˲������������,�����ٽ�CH4���ŷš�����,WATANABE��[53]�о�����,����CH4�ŷ���ոѻ��������������,���ոѻ�����Ϊ2��4��6 t��hm-2ʱ,CH4�ŷ����ֱ�����19%��97%��228%,���нո��е�̼��CH4�ŷŵĹ��ֱ�Ϊ19%��51%��60%�����о���,RW��RP�нոѻ������ﵽ15.15��7.85 t��hm-2,���ڸ��������ࡣ��������̼Դ���з�ֳ������ͬʱ,��Ҫ����ṩ�ϸߵĵ�Դ[46]����RC-G������������ո������٣�4.30 t��hm-2��,�������ϸߡ�����,������Ϊ����ϵͳ�нոѻ���C/N����Ժ��������Ǹ��ŵ���ˮƽ�Ե���CH4�ŷ����õĹؼ�������,���о�����,��������CH4�ŷ���������ȡ���ڲ������������������е�ijһ��,�������������������������ȱ�ֵ��mcrA/pmoA���ܸ��۵ķ�ӳCH4�������������ŷ�֮���������ϵ,����LIU[52]���о����һ�¡������������ɵ�CH4ͨ����ɢ������������ʱ,����CH4��������������������˵���������CH4�ŷ�������ȡ����CH4����������,����������������ԴCH4����������[54]�����о�����,���N0ˮƽ��,��ʩ���ʣ�N1����������RF��RW��RP����ģʽmcrA/pmoA��ֵ;��RC-G����ģʽȴ�����෴����仯������CH4�ŷ�������һ�¡���������Ӣ�̷ʻ�����ʩ���ʣ�N1��,�������м����������Ĵٽ�����ǿ�ڲ������,�����ڼ��ٵ���CH4�ŷš�3.3 ˮ��ֲ�꼰����������ֲ�Ե���CH4�ŷŵ�Ӱ��
��������ŷ��ǵ����������������ʹ����ۺ����õĽ��,��ˮ��������ǿ��Ӱ����3������[55]��ˮ����ϵ��Ϊ��Ҫ���м�����,���ϵ��̬��������������Ӱ��CH4�ŷ�[56],��Ͷ����л����ϼ���ֽ���л�������Ը�ϵ��Ӱ��̶Ȳ�ͬ,���ջᵼ�¼����ŷŵIJ��졣�����ݵ�[57]����ˮ����ϵΪ��ά�������ĺ�������,�������ĸ�ϵ����������,�����˸�ϵ������������ԭ��λ,�ٽ��˵������������������γ�,��Ҳ�����ǵ��²�ͬ����ģʽ�µ���Eh�����Ե�ԭ��֮һ��ͼ6��;ͬʱ,ˮ����ϵͨ��������͵��������ʽΪ��������ṩ����,�ٽ����������ֳ[58]������,ˮ��ֲ��Ҳ��Ӱ�쵾��CH4��������,AULAKH��[59]�о�ָ�������е���������ͨ��ˮ��ֲ���ͨ����֯�����ϵ,�γ�һ����������,�ٽ�����������������,�������������в����ļ���;���������в����ļ����Լ��36%[55]��80%��������[60]�ڸ���������������ϸ�����ġ����о�������Ӣ�̷ʻ�����ʩ���ʺ�CH4�ŷ���Խϵ�,�Ƿ���ˮ��ֲ���ϵ�����̶ȼ���ͨ����֯�йػ��д���������о���������Ϊ�����л����ϼ������ʵ�Ͷ���ˮ��ֲ�꼰���ϵӰ��̶ȴ��ڲ���,������ɵ����������������ʹ�����̵IJ�ͬ,���յ���CH4�ŷŴ��ڲ��졣���о�����ϵͳ��C/N��Ժ�����CH4�ŷ�������ϵ�������������������������Ӱ����������Ч������[61,62],ͬʱҲ��Ӱ�쵾���ոѻ���͵���ʩ��,��Ҳ�����ǵ���������������ԭ��֮һ����������Ч��Դ��̼Դ�������ﷱֳ����������,����ͬ����������ֲ�ܹ�Ӱ�������л��ʺ�������ʵ��������������ṹ[20],�����ı��������л����Ч�л�̼/�������������[63],�Ӷ�����Ӱ�������������������ͻ���[33]�����о���,�����RF,RP��RW��RC-G����ϵͳ������ȫ���ֱ����24.3%��14.0%��7.9%,�л�̼�ֱ����24.2%��7.2%��6.3%����1��������������[64]������[65]�ȵIJ��ֽ���һ�¡�����ȫ�����ܹ���ӳ����������������Ҫָ��,���л�̼��һ���̶���Ӱ���������ṹ�������������,ͬʱ�������������ͻ�������������Ҫ�����ʻ���[66]�����ܲ�ͬ������,����������������,�������о����������Եı仯��������������ոѻ���͵���ʩ�ô���Ѹ���Ҹ�ͨ����CH4�ŷ����,���������ı仯�Ƿ���Ӱ�쵱��CH4�ŷŵ���Ҫ����,�д���һ���о���
4 ����
N0������ͬ����ģʽ�µ���CH4�ŷŲ�������,�������ΪRP>RW>RC-G>RF;��ʩ���ʺ�,����ģʽ�뵪��ˮƽ������������,RW��RP��RF��CH4�ŷ���������,��RC-G�������½�;�����������������������ȱ�ֵ��mcrA/pmoA����CH4�ŷŹ��ɻ���һ�¡�����,������Ϊ����ϵͳ�нոѻ���C/N����Ժ��������Ǹ��ŵ���ˮƽ�Ե���CH4�ŷ����õĹؼ�,��ϵͳ��̼����ʱ,�����������ܵ���������Ч����Լ,Ͷ���������Լ��ᵪ���������ôӶ��������CH4�ŷ�;��̼����ʱ,����Ͷ������,������ﷱֳ�����ܵ�����������̼Դ�����������Ҳ���ܵ�����,CH4�ŷ���Լ��١��ο����� ԭ��˳��
������ȵ���
������������
�����ڿ�Ӱ������
URLMagsci [��������: 1]

<P><FONT face=Verdana>��Ŀ�ġ���ȫ������仯������,�й�������20����80������������ѳ�Ϊ��ʶ,��һ�仯���й�ũҵ���������Ƕ���ֲ�ƶȵ�Ӱ��Խ��Խ�ܵ��й�������ר��****�����ӡ�Ϊ�˻ش���һ����,������1981��Ϊʱ��ڵ�,�Ѵ�20����50��������Ϊ2��ʱ���,�����ͱȽϺ�һʱ�������ů��ȫ����ֲ�ƶȱ��硢��С����ֲ���硢˫������ֲ���硢������С��-�������Ȳ���ֲ����Ŀ���Ӱ��,�Լ�������ֲ����Ŀռ�λ�ƶ�����������ܵ�Ӱ�졣������������ȫ����ֲ�ƶ���������ָ�ꡢ��С���˫������ֲ����ָ���Լ�������С��-�������Ȳ���ֲ����Ľ�ˮ��ָ��,���ù��ϵ�ũҵ����ָ����㷽��,ʹ��ArcGIS�ֱ���1950s��1980�ꡢ1981��2007��2��ʱ��ȫ����ֲ�ƶȱ���ͼ,�Լ���С����ֲ����ͼ��˫������ֲ����ͼ��������С��-�������Ȳ�����ֲ����ͼ�����������1����1950s��1980�����,1981��2007��һ���������ֲ����,�ռ�λ�Ʊ仯��������������ʡ��ɽ��ʡ���ӱ�ʡ������������ʡ��һ����������ֲ����,�ռ�λ�Ʊ仯��������Ϊ����ʡ������ʡ������ʡ������ʡ���㽭ʡ���ڲ�����Ʒ�ֱ仯����ᾭ�õȷ������ص�ǰ����,��Щ������һ��һ����һ�����,������ֲģʽ����ʳ����ƽ��������54%��106%,��һ�������һ������,������ֲģʽ����ʳ����ƽ��������27%��58%����2����1950s��1980�����,1981��2007������ʡ���ӱ�ʡ��ɽ��ʡ������ʡ�����ɹš����ġ�����ʡ���ຣʡ��С�����ֲ���粻ͬ�̶ȱ����������Ժӱ�ʡΪ��,��С����ֲ����ı���,��ʹ���ޱ仯�����ɴ�С����ֶ�С��,����ƽ������Լ25%����3���㽭ʡ������ʡ������ʡ�ͺ���ʡ˫��������ֲ�������ƶ�,����������Դ�ĽǶȳ���,��ʹ����ʳ������ͬ�̶����ӡ���4��������С��-�������Ȳ�����ֲ������������Ϸ����ƶ�,�������ڽ�����������ˮ��������ɵġ������ۡ��ڹ�ȥ��50����,���������ů�����ȫ����ֲ�ƶȽ���ͬ�̶ȱ��ơ���С���˫������ֲ���籱��,���Ƶı仯����ʹ��ֲ�ƶȽ��ޱ仯�������ʳ�������ӡ�Ȼ����ˮ���ļ��������������С��-�������Ȳ��������Ϸ����ƶ���<BR></FONT></P>
URLMagsci [��������: 1]
<P><FONT face=Verdana>��Ŀ�ġ���ȫ������仯������,�й�������20����80������������ѳ�Ϊ��ʶ,��һ�仯���й�ũҵ���������Ƕ���ֲ�ƶȵ�Ӱ��Խ��Խ�ܵ��й�������ר��****�����ӡ�Ϊ�˻ش���һ����,������1981��Ϊʱ��ڵ�,�Ѵ�20����50��������Ϊ2��ʱ���,�����ͱȽϺ�һʱ�������ů��ȫ����ֲ�ƶȱ��硢��С����ֲ���硢˫������ֲ���硢������С��-�������Ȳ���ֲ����Ŀ���Ӱ��,�Լ�������ֲ����Ŀռ�λ�ƶ�����������ܵ�Ӱ�졣������������ȫ����ֲ�ƶ���������ָ�ꡢ��С���˫������ֲ����ָ���Լ�������С��-�������Ȳ���ֲ����Ľ�ˮ��ָ��,���ù��ϵ�ũҵ����ָ����㷽��,ʹ��ArcGIS�ֱ���1950s��1980�ꡢ1981��2007��2��ʱ��ȫ����ֲ�ƶȱ���ͼ,�Լ���С����ֲ����ͼ��˫������ֲ����ͼ��������С��-�������Ȳ�����ֲ����ͼ�����������1����1950s��1980�����,1981��2007��һ���������ֲ����,�ռ�λ�Ʊ仯��������������ʡ��ɽ��ʡ���ӱ�ʡ������������ʡ��һ����������ֲ����,�ռ�λ�Ʊ仯��������Ϊ����ʡ������ʡ������ʡ������ʡ���㽭ʡ���ڲ�����Ʒ�ֱ仯����ᾭ�õȷ������ص�ǰ����,��Щ������һ��һ����һ�����,������ֲģʽ����ʳ����ƽ��������54%��106%,��һ�������һ������,������ֲģʽ����ʳ����ƽ��������27%��58%����2����1950s��1980�����,1981��2007������ʡ���ӱ�ʡ��ɽ��ʡ������ʡ�����ɹš����ġ�����ʡ���ຣʡ��С�����ֲ���粻ͬ�̶ȱ����������Ժӱ�ʡΪ��,��С����ֲ����ı���,��ʹ���ޱ仯�����ɴ�С����ֶ�С��,����ƽ������Լ25%����3���㽭ʡ������ʡ������ʡ�ͺ���ʡ˫��������ֲ�������ƶ�,����������Դ�ĽǶȳ���,��ʹ����ʳ������ͬ�̶����ӡ���4��������С��-�������Ȳ�����ֲ������������Ϸ����ƶ�,�������ڽ�����������ˮ��������ɵġ������ۡ��ڹ�ȥ��50����,���������ů�����ȫ����ֲ�ƶȽ���ͬ�̶ȱ��ơ���С���˫������ֲ���籱��,���Ƶı仯����ʹ��ֲ�ƶȽ��ޱ仯�������ʳ�������ӡ�Ȼ����ˮ���ļ��������������С��-�������Ȳ��������Ϸ����ƶ���<BR></FONT></P>
URL [��������: 1]
.
URL [��������: 1]
[��������: 1]
DOI:10.1016/S0045-6535(02)00822-6URL [��������: 1]
URLMagsci [��������: 3]
<p>ͨ��������飬�о���̫��������ͬ����ģʽ�µ������������ŷŹ���.�������: ˮ��������CH<sub>4</sub>�ŷų������ߺ����ƣ�CH<sub>4</sub>�ŷ���Ҫ������ˮ������ǰ�ڣ��������ˮ���ջ��ڼ�CH<sub>4</sub>�ŷ����ϵͣ�N<sub>2</sub>O���ŷ���Ҫ������3��ʩ�ʼ�������.�����ŷŵ�CH<sub>4</sub>��ȫ������DZ��(GWP)�Ĺ���Զ����N<sub>2</sub>O����������ռ����Ϊ94.7%��99.6%��������������ŵ���Ҫ����.��ͬ����ģʽ�£�����CH<sub>4</sub>�ŷ���������GWP�����������죬����ΪС��-ˮ��>����Ӣ-ˮ��>����-ˮ������������N<sub>2</sub>O�ŷ���������GWPû�������Բ���.�벻ʩ�ʴ�����ȣ�����Ӣ-ˮ������ģʽ��ʩ�ӵ�������������CH<sub>4</sub>�ŷ�����GWP������ͬ���������µ�CH<sub>4</sub>�ŷ�����GWPû�������Բ��죬������Ӣ���ﵾ��ʩ��240 kg·hm<sup>-2</sup>���µ�ˮ������ȴ���.�ۺϾ���Ч��ͻ���Ч�棬����Ӣ���ﵾ��ʩ��240 kg·hm<sup>-2</sup>���µ����������ۺ�Ч�����ã���ֵ�õ����ƹ�ĸ����ƶ�.</p>
URLMagsci [��������: 3]
<p>ͨ��������飬�о���̫��������ͬ����ģʽ�µ������������ŷŹ���.�������: ˮ��������CH<sub>4</sub>�ŷų������ߺ����ƣ�CH<sub>4</sub>�ŷ���Ҫ������ˮ������ǰ�ڣ��������ˮ���ջ��ڼ�CH<sub>4</sub>�ŷ����ϵͣ�N<sub>2</sub>O���ŷ���Ҫ������3��ʩ�ʼ�������.�����ŷŵ�CH<sub>4</sub>��ȫ������DZ��(GWP)�Ĺ���Զ����N<sub>2</sub>O����������ռ����Ϊ94.7%��99.6%��������������ŵ���Ҫ����.��ͬ����ģʽ�£�����CH<sub>4</sub>�ŷ���������GWP�����������죬����ΪС��-ˮ��>����Ӣ-ˮ��>����-ˮ������������N<sub>2</sub>O�ŷ���������GWPû�������Բ���.�벻ʩ�ʴ�����ȣ�����Ӣ-ˮ������ģʽ��ʩ�ӵ�������������CH<sub>4</sub>�ŷ�����GWP������ͬ���������µ�CH<sub>4</sub>�ŷ�����GWPû�������Բ��죬������Ӣ���ﵾ��ʩ��240 kg·hm<sup>-2</sup>���µ�ˮ������ȴ���.�ۺϾ���Ч��ͻ���Ч�棬����Ӣ���ﵾ��ʩ��240 kg·hm<sup>-2</sup>���µ����������ۺ�Ч�����ã���ֵ�õ����ƹ�ĸ����ƶ�.</p>
[D].
[��������: 1]
[D].
[��������: 1]
DOI:10.1016/j.orggeochem.2004.10.007URL [��������: 1]
[��������: 1]
DOI:10.1098/rstb.2007.2184URL [��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
.
[��������: 1]
URLMagsci [��������: 1]
���ڵ����ˮ��ȼ������ܱվ�̬�似�����ԭλ�ɼ���������,�о��˿��ƹ��ģʽ�Ե�����鼾���ŷŹ��ɵ�Ӱ��.�������,�عൾ��ˮ��ȫ�����ڵļ����ŷ���Ҫ�����ڷ���ǰ�ں�����,�����ŷ�����Ϊ24.43 g·m-2,ƽ���ŷ���Ϊ7.95 mg·m-2·h-1,����ˮ���������39%.���ֽεľ�ֵ�����Լ�����ʾ,��"����--��������"�عൾ��ļ���ƽ���ŷ�������ˮ�����������Բ���(0.05��P=0.87),��"��������--������"�����Ե�����ˮ����(0.00=P��0.05).�عൾ������ŷ�����ˮ��ȫ�������·��ϵķ�������ˮ�������һ����,7�·��ǻ��������������ŷ������·�,���عൾ������ŷ����ڸ��µı���ϴ�.����,�عൾ����ڱ��ͺ�ˮ�ʵ���ˮ��״̬���������ٵ��������ŷ�,�Ҳ������ʩ�뵪�ʶ�������,�����ܳ�Ϊ���ռ���Ļ�.
URLMagsci [��������: 1]
���ڵ����ˮ��ȼ������ܱվ�̬�似�����ԭλ�ɼ���������,�о��˿��ƹ��ģʽ�Ե�����鼾���ŷŹ��ɵ�Ӱ��.�������,�عൾ��ˮ��ȫ�����ڵļ����ŷ���Ҫ�����ڷ���ǰ�ں�����,�����ŷ�����Ϊ24.43 g·m-2,ƽ���ŷ���Ϊ7.95 mg·m-2·h-1,����ˮ���������39%.���ֽεľ�ֵ�����Լ�����ʾ,��"����--��������"�عൾ��ļ���ƽ���ŷ�������ˮ�����������Բ���(0.05��P=0.87),��"��������--������"�����Ե�����ˮ����(0.00=P��0.05).�عൾ������ŷ�����ˮ��ȫ�������·��ϵķ�������ˮ�������һ����,7�·��ǻ��������������ŷ������·�,���عൾ������ŷ����ڸ��µı���ϴ�.����,�عൾ����ڱ��ͺ�ˮ�ʵ���ˮ��״̬���������ٵ��������ŷ�,�Ҳ������ʩ�뵪�ʶ�������,�����ܳ�Ϊ���ռ���Ļ�.
[��������: 1]
[��������: 1]
DOI:10.3969/j.issn.1001��7216.2015.02.010URLMagsci [��������: 1]
���Ϸ�������˫����������ӢΪ�о��������þ�̬�䪲����ɫ���ֱ���������̷ʺ͵��ݵȽոѻ��������²�ͬˮ�ֹ����Ե���CH4��N2O�ŷš�ˮ�������Լ��ۺ�����ЧӦ��GWP����Ӱ�졣�����������ˮ��F�������ڿ��F��D��F���ͼ�Ъ��ȣ�F��D��F��M������������������ոѻ���������˫����������CH4�ŷ�������208.3 kg/hm2��F��D��F��M��������678.2 kg/hm2��F������֮�䣬���У�����������ռ����CH4�ŷ�����60.6%~71.7%��F��������CH4�ŷ�����������F��D��F��F��D��F��M������P<0.05�����ոѻ���������˫����������N2O�ŷ���Ϊ4.75~8.19 kg/hm2����F������ȣ�F��D��F��M��������N2O�ŷ�ͨ���������ӣ�60.9%������F��F��D��F����֮��û���������졣�絾�����������������ֱ�Ϊ7.76~8.02 t/hm2��7.22~8.69 t/hm2���ոѻ��������£�˫������λ���GWP�͵�λ����GWP�ֱ�Ϊ7648.8~18471.8 kg/hm2��0��48~1.12 kg/kg������F��D��F��F��D��F��M�����ֱ���������F������P<0.05������ˣ��ڽոѻ��������²������ڿ���ͼ�Ъ������������ˮ������ͬ��ʵ��˫�����߲��ͼ���ũҵ�����������DZ��Ӱ��
DOI:10.3969/j.issn.1001��7216.2015.02.010URLMagsci [��������: 1]
���Ϸ�������˫����������ӢΪ�о��������þ�̬�䪲����ɫ���ֱ���������̷ʺ͵��ݵȽոѻ��������²�ͬˮ�ֹ����Ե���CH4��N2O�ŷš�ˮ�������Լ��ۺ�����ЧӦ��GWP����Ӱ�졣�����������ˮ��F�������ڿ��F��D��F���ͼ�Ъ��ȣ�F��D��F��M������������������ոѻ���������˫����������CH4�ŷ�������208.3 kg/hm2��F��D��F��M��������678.2 kg/hm2��F������֮�䣬���У�����������ռ����CH4�ŷ�����60.6%~71.7%��F��������CH4�ŷ�����������F��D��F��F��D��F��M������P<0.05�����ոѻ���������˫����������N2O�ŷ���Ϊ4.75~8.19 kg/hm2����F������ȣ�F��D��F��M��������N2O�ŷ�ͨ���������ӣ�60.9%������F��F��D��F����֮��û���������졣�絾�����������������ֱ�Ϊ7.76~8.02 t/hm2��7.22~8.69 t/hm2���ոѻ��������£�˫������λ���GWP�͵�λ����GWP�ֱ�Ϊ7648.8~18471.8 kg/hm2��0��48~1.12 kg/kg������F��D��F��F��D��F��M�����ֱ���������F������P<0.05������ˣ��ڽոѻ��������²������ڿ���ͼ�Ъ������������ˮ������ͬ��ʵ��˫�����߲��ͼ���ũҵ�����������DZ��Ӱ��
[��������: 1]
[D].
[��������: 1]
[D].
[��������: 1]
URLMagsci [��������: 1]
<P><��> ��ų��ڱ�ʡ��������ƶ�,�Զ��л��̷ʼ�һ���е�(����������������)Ϊ���������硰���������͡����������̷ʡ�˫������,����ˮ���������纵��ˮ���ȸ��ַ�ʽ,Ҳ��һ�����������,����С����IJ�����ġ������ޡ�����ˮ����������ʮ����ʮ���,��������ƶȸĸ���Ҫ������߸���ָ��,��չ���̷ʡ�˫��
URLMagsci [��������: 1]
<P><��> ��ų��ڱ�ʡ��������ƶ�,�Զ��л��̷ʼ�һ���е�(����������������)Ϊ���������硰���������͡����������̷ʡ�˫������,����ˮ���������纵��ˮ���ȸ��ַ�ʽ,Ҳ��һ�����������,����С����IJ�����ġ������ޡ�����ˮ����������ʮ����ʮ���,��������ƶȸĸ���Ҫ������߸���ָ��,��չ���̷ʡ�˫��
[��������: 1]
.
[��������: 1]
[��������: 2]
[��������: 2]
URL [��������: 1]
URLMagsci [��������: 1]
ѡȡ����˫�����ﳤ�ڲ�ͬʩ���ƶ�Ϊ�о�����,���þ�̬��-����ɫ����������������ŷŽ��й۲⡣�о������������ͬʩ���ƶ��µ�������������ŷŵļ��ڱ仯����һ���Ĺ��ɣ�������������CH4���ŷ����ʳ������ߺ͵ı仯���ơ�ʩ��ոѵĴ���CH4ƽ���ŷ�ͨ�����ۻ��ŷ�ͨ�����ڵ�ʩ���ʵĴ�������ʩ���ʵĸ���������������ȱʧ����IJ�ͬ��CH4ƽ���ŷ�ͨ�����ۻ��ŷ�������һ���IJ��졣�ո���CH4ƽ���ŷ�ͨ�����ۻ��ŷ������ϴ�ȫ����������ʩ������֮��ƫʩ���ֺ�������С��ͬʱ���о��˳��ڲ�ͬʩ���ƶ������¸��������ذ��������¶ȡ����ˮ����Ⱥ�����Eh����CH4�ŷŵ�Ӱ�졣�����������ͬ��ʩ�ʴ�����������CH4�ŷŵļ��ڱ仯������Eh����������أ��������¶ȳ���������أ���ˮ�������ز����ԡ�
URLMagsci [��������: 1]
ѡȡ����˫�����ﳤ�ڲ�ͬʩ���ƶ�Ϊ�о�����,���þ�̬��-����ɫ����������������ŷŽ��й۲⡣�о������������ͬʩ���ƶ��µ�������������ŷŵļ��ڱ仯����һ���Ĺ��ɣ�������������CH4���ŷ����ʳ������ߺ͵ı仯���ơ�ʩ��ոѵĴ���CH4ƽ���ŷ�ͨ�����ۻ��ŷ�ͨ�����ڵ�ʩ���ʵĴ�������ʩ���ʵĸ���������������ȱʧ����IJ�ͬ��CH4ƽ���ŷ�ͨ�����ۻ��ŷ�������һ���IJ��졣�ո���CH4ƽ���ŷ�ͨ�����ۻ��ŷ������ϴ�ȫ����������ʩ������֮��ƫʩ���ֺ�������С��ͬʱ���о��˳��ڲ�ͬʩ���ƶ������¸��������ذ��������¶ȡ����ˮ����Ⱥ�����Eh����CH4�ŷŵ�Ӱ�졣�����������ͬ��ʩ�ʴ�����������CH4�ŷŵļ��ڱ仯������Eh����������أ��������¶ȳ���������أ���ˮ�������ز����ԡ�
[��������: 2]
[��������: 1]
[��������: 1]
[��������: 2]
[��������: 2]
DOI:10.1111/fem.2012.82.issue-2URL [��������: 1]
[��������: 1]
DOI:10.1016/j.agee.2008.11.006URL [��������: 1]
[��������: 1]
[��������: 1]
URLMagsci [��������: 1]
Ϊȷ�����ҹ��������������ŷ��嵥���ƶ��������Ŵ�ʩ�ṩ��������, ѡ��̫����������ˮ����ֲ������ʡ������, �о���������� ˮ��(����, CK)������Ӣ ˮ��(T1)������� ˮ��(T2)��С�� ˮ��(T3)���Ͳ� ˮ��(T4)5��ˮ��������ʽ, ���þ�̬�� ����ɫ��, ��չ�˲�ͬˮ��������ʽ��ˮ��������������(CH<sub>4</sub>)�������ǵ�(N<sub>2</sub>O)�ŷż�����顣����������: ��ͬˮ��������ʽ��ˮ��������CH4�ŷ�ͨ���������ߺ͵ı仯����, CH4�ŷŷ�ֵ������ˮ������ǰ��, ��������Ч�����ٽ�Ҷ����CH<sub>4</sub>�ۻ��ŷ���ռȫ�������ŷ������ı���Ϊ65%~81%, ��N2O����ˮ�������ڼ��������ŷš�ˮ��������ʽ�Ե���CH<sub>4</sub>��N<sub>2</sub>O�ŷ��м�����(<i>P</i> 0.01)Ӱ��, CH<sub>4</sub>�������ŷ�������ΪT1(283.2 kg?hm<sup>-2</sup>) CK(139.5 kg?hm<sup>-2</sup>) T3(123.4 kg?hm<sup>-2</sup>) T4(114.7 kg?hm<sup>-2</sup>) T2(100.8 kg?hm<sup>-2</sup>), N<sub>2</sub>O�������ŷ���˳��ΪT1 T4 T3 T2 CK, ����Ϊ1.06 kg?hm <sup>-2</sup>��0.87 kg?hm<sup>-2</sup>��0.81 kg?hm<sup>-2</sup>��0.72 kg?hm<sup>-2</sup>��0.53 kg?hm<sup>-2</sup>��T1���������ŷ�CH<sub>4</sub>��N<sub>2</sub>O����������DZ�����[7 396 kg(CO<sub>2</sub>)?hm<sup>-2</sup>], ����(<i>P</i> 0.05)������������, ��CK[3 646 kg(CO<sub>2</sub>)?hm <sup>-2</sup>]����103%, T2[2 735 kg(CO<sub>2</sub>)?hm <sup>-2</sup>]��CK����25%(<i>P</i> 0.05)������Ӣ ˮ��������ʽ������̫������ˮ��������������ЧӦ��
URLMagsci [��������: 1]
Ϊȷ�����ҹ��������������ŷ��嵥���ƶ��������Ŵ�ʩ�ṩ��������, ѡ��̫����������ˮ����ֲ������ʡ������, �о���������� ˮ��(����, CK)������Ӣ ˮ��(T1)������� ˮ��(T2)��С�� ˮ��(T3)���Ͳ� ˮ��(T4)5��ˮ��������ʽ, ���þ�̬�� ����ɫ��, ��չ�˲�ͬˮ��������ʽ��ˮ��������������(CH<sub>4</sub>)�������ǵ�(N<sub>2</sub>O)�ŷż�����顣����������: ��ͬˮ��������ʽ��ˮ��������CH4�ŷ�ͨ���������ߺ͵ı仯����, CH4�ŷŷ�ֵ������ˮ������ǰ��, ��������Ч�����ٽ�Ҷ����CH<sub>4</sub>�ۻ��ŷ���ռȫ�������ŷ������ı���Ϊ65%~81%, ��N2O����ˮ�������ڼ��������ŷš�ˮ��������ʽ�Ե���CH<sub>4</sub>��N<sub>2</sub>O�ŷ��м�����(<i>P</i> 0.01)Ӱ��, CH<sub>4</sub>�������ŷ�������ΪT1(283.2 kg?hm<sup>-2</sup>) CK(139.5 kg?hm<sup>-2</sup>) T3(123.4 kg?hm<sup>-2</sup>) T4(114.7 kg?hm<sup>-2</sup>) T2(100.8 kg?hm<sup>-2</sup>), N<sub>2</sub>O�������ŷ���˳��ΪT1 T4 T3 T2 CK, ����Ϊ1.06 kg?hm <sup>-2</sup>��0.87 kg?hm<sup>-2</sup>��0.81 kg?hm<sup>-2</sup>��0.72 kg?hm<sup>-2</sup>��0.53 kg?hm<sup>-2</sup>��T1���������ŷ�CH<sub>4</sub>��N<sub>2</sub>O����������DZ�����[7 396 kg(CO<sub>2</sub>)?hm<sup>-2</sup>], ����(<i>P</i> 0.05)������������, ��CK[3 646 kg(CO<sub>2</sub>)?hm <sup>-2</sup>]����103%, T2[2 735 kg(CO<sub>2</sub>)?hm <sup>-2</sup>]��CK����25%(<i>P</i> 0.05)������Ӣ ˮ��������ʽ������̫������ˮ��������������ЧӦ��
[D].
[��������: 1]
[D].
[��������: 1]
[D].
[��������: 1]
[D].
[��������: 1]
[��������: 2]
DOI:10.1016/j.soilbio.2007.05.030URL [��������: 1]
URL [��������: 1]
[��������: 1]
.
[��������: 1]
[��������: 1]
[��������: 1]
DOI:10.1111/fem.2014.88.issue-2URL [��������: 2]
DOI:10.1007/BF00335999URL [��������: 1]
[��������: 1]
[��������: 1]
[D].
[��������: 1]
[D].
[��������: 1]
[��������: 1]
[��������: 1]
.
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 3]
[��������: 1]
[��������: 1]
DOI:10.11686/cyxb2014313URLMagsci [��������: 1]
������Ϊ�����������,��Դ����ΪӰ��������������������ʹ�����̵���Ҫ���ء����������ŷ��ܵ�����Ч�Եĵ���,�������ı�������ʼ�����ͼ����ŷŹ���,����Ӱ������ŷ����������˵������������������������ʹ�����̵�Ӱ�켰���ơ��о�����,1)������Լ����ŷ�ͨ����Ӱ����ڴٽ������ƺͲ�����3�����,����Ҫ�Ǽ�������������ʹ�����̵ı仯�����;2)������Լ���������̵�Ӱ���ܲ��������Ͳ�����������ԵĿ���,������ͨ�����������л�̼�ĺ���Ϊ��������ṩ�˷ḻ�ĵ���,ͬʱ�����������ʺ�ֲ�����Ƕȵı仯ʹ������Ӱ�츴�ӻ�,������ȿɴٽ��ֿ����Ʋ�������Ļ���,�������������ܵ���̬��Ӱ��;3)������Լ����������̵�Ӱ����Ҫ��ͨ���̼������Ƽ����������Ļ���ʵ�ֵ�,����̬�IJ�ͬҲʹ�����ֱ仯��Ϊ����;4)������Լ��鴫����̵�Ӱ����Ҫ��ֲ��ͨ����֯�������Լ�����Ч�ʵĿ���,�����ڲ�ͬ��̬ϵͳ���ֿ������ò���ϴ���������,�������������������������ʹ�����̵�Ӱ�켰���ƾ������Եĸ����ԺͲ�ȷ����,����о���Ӧ�ۺϿ��ǵ�����Լ����ŷŹؼ����̵�Ӱ��,��������̽�ֵ�������������Ⱥ��ṹ����Ⱥͻ��Ե�Ӱ��,ͬʱע�ضԸ�����̬ϵͳ��Эͬ�о�,ȷ��������Ӱ���¸�����̬ϵͳ��ȫ������ŷŵĹ����ʡ�
DOI:10.11686/cyxb2014313URLMagsci [��������: 1]
������Ϊ�����������,��Դ����ΪӰ��������������������ʹ�����̵���Ҫ���ء����������ŷ��ܵ�����Ч�Եĵ���,�������ı�������ʼ�����ͼ����ŷŹ���,����Ӱ������ŷ����������˵������������������������ʹ�����̵�Ӱ�켰���ơ��о�����,1)������Լ����ŷ�ͨ����Ӱ����ڴٽ������ƺͲ�����3�����,����Ҫ�Ǽ�������������ʹ�����̵ı仯�����;2)������Լ���������̵�Ӱ���ܲ��������Ͳ�����������ԵĿ���,������ͨ�����������л�̼�ĺ���Ϊ��������ṩ�˷ḻ�ĵ���,ͬʱ�����������ʺ�ֲ�����Ƕȵı仯ʹ������Ӱ�츴�ӻ�,������ȿɴٽ��ֿ����Ʋ�������Ļ���,�������������ܵ���̬��Ӱ��;3)������Լ����������̵�Ӱ����Ҫ��ͨ���̼������Ƽ����������Ļ���ʵ�ֵ�,����̬�IJ�ͬҲʹ�����ֱ仯��Ϊ����;4)������Լ��鴫����̵�Ӱ����Ҫ��ֲ��ͨ����֯�������Լ�����Ч�ʵĿ���,�����ڲ�ͬ��̬ϵͳ���ֿ������ò���ϴ���������,�������������������������ʹ�����̵�Ӱ�켰���ƾ������Եĸ����ԺͲ�ȷ����,����о���Ӧ�ۺϿ��ǵ�����Լ����ŷŹؼ����̵�Ӱ��,��������̽�ֵ�������������Ⱥ��ṹ����Ⱥͻ��Ե�Ӱ��,ͬʱע�ضԸ�����̬ϵͳ��Эͬ�о�,ȷ��������Ӱ���¸�����̬ϵͳ��ȫ������ŷŵĹ����ʡ�
URLMagsci [��������: 1]
�������������Լ�����ΪΨһ̼Դ����Դ����ͬ�����컯��л������, ��ؼ�ø֮һ�Ǽ��鵥����ø(MMOs), �����������������´�����ȵ�̼������ϩ���ǻ���������, ��������������Ȼ��̼ѭ����ҵ���\���о�����Ҫ��Ӧ�ü�ֵ�����, ��20�������ڼ�����������MMOs���о�һֱ��������ѧ�ҵĹ�ע�����´��ִ����\���ĽǶ�, �Խ������������ڼ����������ķ�����ֲ�, MMOs�Ľṹ�빦�ܡ�������������MMOs�Ļ��̵ȷ���ȡ�õ��о��ɹ��������ܽ�, ȫ�������˼�����������MMOs��Ӧ�û����о���״, ����������о���Ӧ�÷��������չ����
URLMagsci [��������: 1]
�������������Լ�����ΪΨһ̼Դ����Դ����ͬ�����컯��л������, ��ؼ�ø֮һ�Ǽ��鵥����ø(MMOs), �����������������´�����ȵ�̼������ϩ���ǻ���������, ��������������Ȼ��̼ѭ����ҵ���\���о�����Ҫ��Ӧ�ü�ֵ�����, ��20�������ڼ�����������MMOs���о�һֱ��������ѧ�ҵĹ�ע�����´��ִ����\���ĽǶ�, �Խ������������ڼ����������ķ�����ֲ�, MMOs�Ľṹ�빦�ܡ�������������MMOs�Ļ��̵ȷ���ȡ�õ��о��ɹ��������ܽ�, ȫ�������˼�����������MMOs��Ӧ�û����о���״, ����������о���Ӧ�÷��������չ����
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 2]
DOI:10.1029/97JD03460URL [��������: 1]
Magsci [��������: 1]
Ϊ��̽�ֲ�ͬˮ�ֹ����ͽոѻ���Ե�������ŷŵ�Ӱ������������й���ѧԺ���ȴ�ũҵ��̬�о�����ɳũҵ��������о�վ���ڶ�λ���飬ѡ������ˮ��CF�����ոѻ���+������ˮ��HS+CF���������ȣ�IF�����ոѻ���+�����ȣ�HS+IF��4��������Ӧ��������Ƭ�γ��ȶ�̬����(T-RFLP)��ʵʱӫ�ⶨ��PCR������DNA��cDNAˮƽ�Ա��о���ͬˮ�ֹ����ͽոѻ���Ӱ�쵾�����������ŷŵ�������ӻ��������������������ˮ�����ļ����ŷ����������ڳ����ȴ������ոѻ��ﴦ���Լ����ŷŵ�Ӱ�첻��������������ͼ���������Ⱥ��ṹֻ������ˮ�ֹ���������Ӧ���Խոѻ���û����Ӧ��ˮ�ֹ����ͽոѻ�������Ӱ����������������ȴ�Լ�������������û��Ӱ�졣��������ͼ���������Ⱥ����ɺͱ���Ⱥ����ɴ������Եķ��죬cDNAˮƽ�ϲ�������ͼ�������������Ⱥ����ɶ�ˮ�ֹ�������Ӧ��DNAˮƽȺ����ɸ����С���������ŷ�ͨ����mcrA��pmoA�����Ⱥͱ����Ⱦ�û����������ԣ�ֻ��DNAˮƽ�����ȱ�ֵ��mcrA/pmoA������������ع�ϵ�����Կ�����������ˮ��������ͨ���ı������в�������ͼ�����������Ⱥ��ṹ�����������������ı��������ؼ�����ŷ�����
Magsci [��������: 1]
Ϊ��̽�ֲ�ͬˮ�ֹ����ͽոѻ���Ե�������ŷŵ�Ӱ������������й���ѧԺ���ȴ�ũҵ��̬�о�����ɳũҵ��������о�վ���ڶ�λ���飬ѡ������ˮ��CF�����ոѻ���+������ˮ��HS+CF���������ȣ�IF�����ոѻ���+�����ȣ�HS+IF��4��������Ӧ��������Ƭ�γ��ȶ�̬����(T-RFLP)��ʵʱӫ�ⶨ��PCR������DNA��cDNAˮƽ�Ա��о���ͬˮ�ֹ����ͽոѻ���Ӱ�쵾�����������ŷŵ�������ӻ��������������������ˮ�����ļ����ŷ����������ڳ����ȴ������ոѻ��ﴦ���Լ����ŷŵ�Ӱ�첻��������������ͼ���������Ⱥ��ṹֻ������ˮ�ֹ���������Ӧ���Խոѻ���û����Ӧ��ˮ�ֹ����ͽոѻ�������Ӱ����������������ȴ�Լ�������������û��Ӱ�졣��������ͼ���������Ⱥ����ɺͱ���Ⱥ����ɴ������Եķ��죬cDNAˮƽ�ϲ�������ͼ�������������Ⱥ����ɶ�ˮ�ֹ�������Ӧ��DNAˮƽȺ����ɸ����С���������ŷ�ͨ����mcrA��pmoA�����Ⱥͱ����Ⱦ�û����������ԣ�ֻ��DNAˮƽ�����ȱ�ֵ��mcrA/pmoA������������ع�ϵ�����Կ�����������ˮ��������ͨ���ı������в�������ͼ�����������Ⱥ��ṹ�����������������ı��������ؼ�����ŷ�����
[��������: 2]
[��������: 2]
[��������: 1]
[��������: 1]
URLMagsci [��������: 1]
Ϊ��ȷ�Ͳ˽ոѻ��������������ˮ����ϵ������Ӱ�죬���о������������������ģ���������ϵķ������������Ͳ˽ոѻ����������������ԭ��λ��EHֵ������ԭ������������Fe2+�����ı仯���ɼ�ˮ�����ڸ�ϵ��������������������Ⱥ�����ǿ�ȵȵı仯���ɡ����������1���Ͳ˽ոѸ��ǻ��������£���������������ԭ��λ�վ��仯����RCEHֵ������Ϊ�Ƚ��ͺ����ӵ����ƣ���������ԭ�����������վ��仯����RCRRMֵ�������������������վ��仯����RCFeֵ������Ϊ�����Ӻ͵����ƣ�����ˮ�����Ժ�14��18 d����RCEHֵ��ͣ����Ժ�10��14 d��14��22 d����RCRRMֵ(0.28��0.62 cmol/(kg?d))��RCFeֵ(5.0��31.6 ��g/(kg?d))�ﵽ���2���Ͳ˽ոѸ��ǻ��������£����Žոѻ����������ӣ�ˮ����ϵ����������������������ͳ��ȣ�����������ϵ����ǿ���������ͣ��븲����ȣ��Ͳ˽ոѷ����ˮ����ϵ�����վ��仯����RCRBI����Ӱ��ʱ�������ˮ����ϵ����վ��仯����RCRVTֵ���������վ��仯����RCRQTֵ���Ļ����������ӳ���4 d��3��ˮ�����Ժ�10��22 d���Ͳ˽ոѻ���Ե�������������ˮ����ϵ������Ӱ�������������Ժ�27��32 d���Ͳ˽ոѻ����ˮ����ϵ������Ӱ��̶��������͡��Ͳ˽ոѻ��������£����Žոѻ��������ӣ�ˮ�����ڵ�����������ԭ��λ�������ͣ�������ԭ������������Fe2+�����������ӣ�ˮ����ϵ����������������������ͳ��ȣ���ϵ��������������ϵ����ǿ�ȣ��������͡����Ĵ��͵����������Ͳ˽ոѻ�������ȫ������Ϊ�ˣ����˻�е��ҵ��������Ͳ˽ոѷ�����Ϊ�ˡ�
URLMagsci [��������: 1]
Ϊ��ȷ�Ͳ˽ոѻ��������������ˮ����ϵ������Ӱ�죬���о������������������ģ���������ϵķ������������Ͳ˽ոѻ����������������ԭ��λ��EHֵ������ԭ������������Fe2+�����ı仯���ɼ�ˮ�����ڸ�ϵ��������������������Ⱥ�����ǿ�ȵȵı仯���ɡ����������1���Ͳ˽ոѸ��ǻ��������£���������������ԭ��λ�վ��仯����RCEHֵ������Ϊ�Ƚ��ͺ����ӵ����ƣ���������ԭ�����������վ��仯����RCRRMֵ�������������������վ��仯����RCFeֵ������Ϊ�����Ӻ͵����ƣ�����ˮ�����Ժ�14��18 d����RCEHֵ��ͣ����Ժ�10��14 d��14��22 d����RCRRMֵ(0.28��0.62 cmol/(kg?d))��RCFeֵ(5.0��31.6 ��g/(kg?d))�ﵽ���2���Ͳ˽ոѸ��ǻ��������£����Žոѻ����������ӣ�ˮ����ϵ����������������������ͳ��ȣ�����������ϵ����ǿ���������ͣ��븲����ȣ��Ͳ˽ոѷ����ˮ����ϵ�����վ��仯����RCRBI����Ӱ��ʱ�������ˮ����ϵ����վ��仯����RCRVTֵ���������վ��仯����RCRQTֵ���Ļ����������ӳ���4 d��3��ˮ�����Ժ�10��22 d���Ͳ˽ոѻ���Ե�������������ˮ����ϵ������Ӱ�������������Ժ�27��32 d���Ͳ˽ոѻ����ˮ����ϵ������Ӱ��̶��������͡��Ͳ˽ոѻ��������£����Žոѻ��������ӣ�ˮ�����ڵ�����������ԭ��λ�������ͣ�������ԭ������������Fe2+�����������ӣ�ˮ����ϵ����������������������ͳ��ȣ���ϵ��������������ϵ����ǿ�ȣ��������͡����Ĵ��͵����������Ͳ˽ոѻ�������ȫ������Ϊ�ˣ����˻�е��ҵ��������Ͳ˽ոѷ�����Ϊ�ˡ�
[��������: 1]
[��������: 1]
DOI:10.1023/A:1009839929441URL [��������: 1]
DOI:10.1007/BF00334584URL [��������: 1]
[��������: 1]
.
[��������: 1]
[��������: 1]
[��������: 1]
DOI:10.1007/s13593-014-0212-yURL [��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 2]
[��������: 2]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}