 , 武晶
, 武晶Adaptability and Phenotypic Variations of Agronomic Traits in Common Bean Germplasm Resources in Different Environments
WANGLan-Fen, WUJing通讯作者:
收稿日期:2017-04-27
接受日期:2017-11-21
网络出版日期:2018-03-12
版权声明:2018作物学报编辑部作物学报编辑部
基金资助:
作者简介:
-->
展开
摘要
关键词:
Abstract
Keywords:
-->0
PDF (1031KB)元数据多维度评价相关文章收藏文章
本文引用格式导出EndNoteRisBibtex收藏本文-->
普通菜豆(Phaseolus vulgaris L.)是自花授粉一年生二倍体(2n = 2x = 22)作物, 基因组约为587 Mb [1,2]。普通菜豆属于典型的短日照作物[3,4,5], 起源于中南美洲[6,7,8,9,10], 随着它在原生境不断驯化和向外传播, 最终形成中美洲和安第斯2个独立的、包括野生种群和栽培品种, 之间存在生殖隔离的基因库[11]。普通菜豆于16世纪初传播到欧洲, 16、17世纪在欧洲迅速传播, 再由欧洲传入中东、西亚以及世界上其他国家和地区[12]。
普通菜豆是世界范围内仅次于大豆和花生的重要食用豆类, 在生长习性、种子大小、形状、籽粒颜色、生育期等性状上具有较多变异类型[13,14]。普通菜豆籽粒因含有较高的粗纤维(3%~6%)、碳水化合物(61.2%)、蛋白质(20%)、磷脂、不含胆固醇等有利于人类健康和疾病预防的功能成分[15,16], 近年来得到人们广泛的关注。研究表明, 食用普通菜豆籽粒可降低人类患肥胖、糖尿病、心血管疾病、结肠癌、前列腺癌和乳腺癌的风险[15,16,17]。
截至2010年底, 我国编目入库的普通菜豆为4661份, 包括来源于世界18个国家和地区的种质, 但国外种质所占比例仅为4.78%。加强国外引种, 并对收集引进的种质资源安全繁殖、编目入库, 丰富我国普通菜豆资源库是当前的重要任务。同时, 加强对已保存资源的鉴定评价, 才能更好地为生产和育种服务。王述民等[18] 1999年对我国收集的4029份普通菜豆(3845份地方品种、3份育成品种、179份国外品种及其他2份), 从地理分布、生长习性、籽粒性状、抗逆性等方面进行了分析评价, 分析了我国普通菜豆的种质类型。1992、1993年王述民等[19,20]在黑龙江哈尔滨、内蒙古凉城、四川昭觉和山西太原连续2年分别对10份、233份国内种质资源进行了的综合鉴定评价试验, 筛选出一批优异种质。本研究于2014—2015年在黑龙江哈尔滨、河南南阳、贵州毕节和海南乐东, 对国内外686份普通菜豆种质资源进行表型鉴定和生态适应性分析, 旨在明确不同来源的普通菜豆种质资源的生长适宜地区, 为编目入库、繁殖更新及农艺性状精准鉴定提供信息, 为种质创新提供特异资源, 进而促进我国普通菜豆产业的快速发展。
1 材料与方法
1.1 试验材料
供试材料686份普通菜豆, 其中, 国内材料507份, 国外引进材料179份, 详见表1。Table 1
表1
表1材料来源
Table 1Origin of materials
| 中国种质来源 Origin of domestic germplasm | 份数 No. | 引进种质来源 Origin of alien germplasm | 份数 No. |
|---|---|---|---|
| 中国黑龙江 Heilongjiang, China | 86 | 阿根廷 Argentina | 38 |
| 中国山西 Shanxi, China | 66 | 巴西 Brazil | 38 |
| 中国内蒙古 Inner Mongolia, China | 63 | 国际热带农业研究中心 CIAT | 37 |
| 中国贵州 Guizhou, China | 56 | 哥伦比亚 Columbia | 18 |
| 中国吉林 Jilin, China | 42 | 法国 France | 8 |
| 中国云南 Yunnan, China | 37 | 墨西哥 Mexico | 8 |
| 中国湖北 Hubei, China | 34 | 美国 America | 5 |
| 中国陕西 Shaanxi, China | 33 | 秘鲁 Peru | 4 |
| 中国四川 Sichuan, China | 25 | 危地马拉 Guatemala | 3 |
| 中国甘肃 Gansu, China | 21 | 保加利亚 Bulgaria | 2 |
| 中国河北 Hebei, China | 11 | 玻利维亚 Bolivia | 2 |
| 中国北京 Beijing, China | 10 | 多米尼加共和国 Dominican Republic | 1 |
| 中国山东 Shandong, China | 9 | 厄瓜多尔 Ecuador | 1 |
| 中国新疆 Xinjiang, China | 6 | 海地 Haiti | 1 |
| 中国湖南 Hunan, China | 2 | 加拿大 Canada | 1 |
| 中国江苏 Jiangsu, China | 1 | 葡萄牙 Portugal | 1 |
| 中国辽宁 Liaoning, China | 1 | 委内瑞拉 Venezuela | 1 |
| 中国台湾 Taiwan, China | 1 | 英国 UK | 1 |
| 中国西藏 Xizang, China | 1 | 赞比亚 Zambia | 1 |
| 中国海南 Hainan, China | 1 | 智利 Chile | 1 |
| 不详 Unknown | 1 | 不详 Unknown | 7 |
| 合计 Total | 507 | 合计 Total | 179 |
新窗口打开
1.2 试验地点及设计
供试材料分别于2014年4月21日播种于贵州毕节(27°11′9.89″N, 105°18′9.26″E), 4月30日播种于河南南阳(32°55′50.98″N, 112°24′28.55″E), 5月23日播种于黑龙江哈尔滨(45°50′46.88″N, 126°51′16.74″E), 11月19日播种于海南乐东(18°26′25.26″N, 108°54′ 52.23″E)。按蔓生、矮生随机排列, 每个材料种植2行, 行长4 m, 穴距40 cm, 哈尔滨行距70 cm, 其余行距50 cm。在哈尔滨、毕节和南阳(播种前浇底水)全生育期雨养, 在乐东人工定期喷灌。1.3 调查和考种性状
参考《普通菜豆种质资源描述规范和数据标 准》[21], 调查出苗日期、开花期、成熟期、主茎节数、主茎分枝数、株高、单株荚数、荚长、荚宽、单荚粒数、单株产量和百粒重。1.4 数据处理
采用Microsoft Excel和SPSS19软件统计分析数据。遗传力公式为h2 = (V1-V2)/[V1+(r-1)V2], 其中V1为组间均方差, V2为组内均方差, r为重复数或环境数。
通径系数(P)与相关系数(r)的方程为:
rly = rllPl + rl2P2 +…+ rlnPn
r2y = r2lPl + r22P2 +…+ r2nPn
rny = rnlPl + rn2P2 +…+ rnnPn
2 结果与分析
2.1 普通菜豆在不同环境下的结实情况
普通菜豆属于光温敏型豆类, 在长日照环境下, 会出现不开花、开花不结荚或结荚不成熟等现象。从播种至成熟, 各地的日照时长及温度存在明显差异(表2), 哈尔滨的平均日照时数最长, 为14.47 h, 乐东最短, 为11.11 h, 南阳和毕节分别为13.88 h和13.33 h。南阳平均日最高气温、最低气温均最高(30.0°C、24.1°C), 其余3个试点的温度日均最高气温、最低气温相差不大。Table 2
表2
表24个试点的日照时数和温度概况及686份普通菜豆的结实情况
Table 2Sunshine duration, temperature condition and seed setting rate of 686 common bean accessions in four locations
| 地点 Location | 播种-收获日期Sowing-harvest date (month/day) | 日照时长 Sunshine hours (h) | 日最高温度 Daily max. temp. (°C) | 日最低温度 Daily min. temp. (°C) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 平均值 Mean | 最大值Max. | 最小值Min. | 平均值 Mean | 最大值Max. | 最小值Min. | 平均值 Mean | 最大值Max. | 最小值Min. | |||||||
| 哈尔滨 Harbin | 5/23-9/30 | 14.47 | 15.72 | 11.77 | 26.0 | 37 | 10 | 16.0 | 24 | -2 | |||||
| 南阳 Nanyang | 4/30-8/31 | 13.88 | 14.32 | 12.88 | 30.0 | 38 | 21 | 21.2 | 28 | 11 | |||||
| 毕节 Bijie | 4/21-9/22 | 13.33 | 13.85 | 12.13 | 24.1 | 31 | 13 | 16.3 | 21 | 9 | |||||
| 乐东 Ledong | 11/19-2/23 | 11.11 | 11.63 | 10.59 | 26.3 | 32 | 19 | 16.1 | 23 | 8 | |||||
| 种质材料份数 No. of accessions | 结实率 Seed setting rate (%) | ||||||||||||||
| 未出苗Emergence-failed | 幼苗死亡 Seedling-died | 未开花 Bloom-failed | 未成熟 Immature | 正常结实 Fertile | |||||||||||
| 哈尔滨 Harbin | 6 | 10 | 45 | 16 | 609 | 88.78 | |||||||||
| 南阳 Nanyang | 0 | 6 | 69 | 46 | 565 | 82.36 | |||||||||
| 毕节 Bijie | 9 | 7 | 2 | 39 | 629 | 91.69 | |||||||||
| 乐东 Ledong | 2 | 1 | 0 | 0 | 683 | 99.56 | |||||||||
新窗口打开
4个试点除南阳外均有未出苗的材料。其中, 毕节未出苗材料较多, 为9份; 哈尔滨和乐东分别为6份和2份。后期因病害也有少量材料死亡, 哈尔滨最多, 为10份, 乐东仅有1份, 南阳和毕节分别为7份和6份。由于各地气候条件的差异, 除乐东的试点皆出现未开花或开花不成熟现象。南阳和哈尔滨的未开花材料达69份和45份, 其中44份为国外野生材料, 其余大部分来自云贵川。同时南阳未成熟材料最多, 达46份, 来自于国内外。毕节39份未成熟资源中, 38份为野生材料, 哈尔滨未成熟的16份材料中, 13份来源于云贵川。乐东参试点材料的结实率最高为99.56%, 毕节次之, 为91.69%, 南阳最低, 为82.36% (表2)。
2.2 表型性状变异及遗传力分析
对686份普通菜豆种质资源的12个表型性状统计分析表明, 不同种质在各试点皆存在很大差异, 表现出很高的变异度(表3), 平均变异系数介于8.11%~70.83%之间。其中, 株高的变异系数最大, 在毕节、乐东、哈尔滨、南阳依次为82.52%、71.13%、68.31%和61.37%; 单株荚数和主茎节数在各试点的平均变异系数相当, 分别为65.97%和65.23%; 单株荚数在南阳的变异系数最大(86.93%), 毕节次之(67.68%), 乐东最小(50.93%); 主茎节数的变异系数在各点差异较大, 哈尔滨为89.90%, 毕节66.66%, 而乐东最小, 仅为39.10%; 单株产量在毕节的变异系数最大(69.63%), 哈尔滨次之(57.22%), 乐东最小(50.35%); 百粒重的变异系数在各点变化不大, 变化范围为31.98%~45.52%; 主茎分枝数、单荚粒数、荚长、荚宽的平均变异系数介于20.0%~30.0%之间; 与物候期有关的播种至出苗时间、开花时间和生育期在4个试点的平均变异系数相对较小, 分别为8.11%、15.78%和8.94%。播种至开花时间在毕节最长, 平均56.33 d, 乐东最短, 仅36.63 d。在乐东, 播种49 d全部材料皆已开花, 而在其他三地, 80 d后仍有相当数量的材料不能开花。在乐东播种99 d全部材料均已成熟, 而在哈尔滨、南阳、毕节最晚成熟材料的生育期分别为129、122和154 d。供试材料在哈尔滨株高普遍偏高, 平均154.76 cm、南阳次之(133.20 cm), 乐东最矮(79.12 cm)。哈尔滨的单株荚数最多, 平均32.32个、南阳次之(20.23)、毕节最少(9.86)。哈尔滨的平均单荚粒数最高(5.67), 南阳的最低, 仅为3.69粒。平均百粒重、荚长在哈尔滨、毕节和乐东相差不大, 在南阳相对较低。单株产量在哈尔滨最高, 达40.55 g, 在毕节和乐东分别为9.67 g和12.39 g。平均荚宽在4个试点基本一致。Table 3
表3
表3686份普通菜豆材料12个农艺性状在4个试点的变异
Table 3Variation of 686 accessions of common bean in 12 agronomic traits in four locations
| 性状 Trait | 地点 Location | 最小值 Min. | 最大值 Max. | 均值 Average | 标准差 SD | 变异系数 CV (%) | 遗传力Heritability |
|---|---|---|---|---|---|---|---|
| 播种至出苗时间 Days from sowing to emergence (d) | 哈尔滨 Harbin | — | — | — | — | — | — |
| 南阳 Nanyang | 9.00 | 14.00 | 12.19 | 0.96 | 7.92 | ||
| 毕节 Bijie | 17.00 | 23.00 | 20.38 | 0.97 | 4.74 | ||
| 乐东 Ledong | 7.00 | 13.00 | 7.80 | 0.91 | 11.68 | ||
| 播种至开花时间 Days from sowing to flowering (d) | 哈尔滨 Harbin | 33.00 | 82.00 | 48.72 | 11.78 | 24.18 | 0.01 |
| 南阳 Nanyang | 33.00 | 89.00 | 49.43 | 7.86 | 15.91 | ||
| 毕节 Bijie | 45.00 | 80.00 | 56.33 | 7.49 | 13.29 | ||
| 乐东 Ledong | 30.00 | 49.00 | 36.63 | 3.57 | 9.74 | ||
| 生育期 | 哈尔滨 Harbin | 69.00 | 129.00 | 100.14 | 14.13 | 14.11 | 0.58 |
| Growth duration (d) | 南阳 Nanyang | 63.00 | 122.00 | 88.79 | 5.89 | 6.63 | |
| 毕节 Bijie | 92.00 | 154.00 | 106.64 | 7.77 | 7.29 | ||
| 乐东 Ledong | 67.00 | 99.00 | 80.41 | 6.22 | 7.74 | ||
| 株高 | 哈尔滨 Harbin | 22.00 | 385.00 | 154.76 | 105.72 | 68.31 | 0.58 |
| Plant height (cm) | 南阳 Nanyang | 14.29 | 376.14 | 133.20 | 81.75 | 61.37 | |
| 毕节 Bijie | 6.00 | 440.00 | 113.80 | 93.91 | 82.52 | ||
| 乐东 Ledong | 18.00 | 280.00 | 79.12 | 56.28 | 71.13 | ||
| 主茎节数 | 哈尔滨 Harbin | 2.00 | 27.20 | 6.52 | 5.86 | 89.90 | 0.28 |
| Node number per plant | 南阳 Nanyang | — | — | — | — | — | |
| 毕节 Bijie | 1.80 | 24.00 | 7.66 | 5.11 | 66.66 | ||
| 乐东 Ledong | 3.40 | 32.67 | 13.20 | 5.16 | 39.10 | ||
| 主茎分枝数 Branch number per plant | 哈尔滨 Harbin | 2.20 | 8.40 | 4.53 | 1.05 | 23.14 | — |
| 南阳 Nanyang | 3.00 | 18.00 | 6.49 | 2.32 | 35.78 | ||
| 毕节 Bijie | 1.00 | 5.00 | 1.86 | 0.57 | 30.85 | ||
| 乐东 Ledong | 1.00 | 6.71 | 2.45 | 0.69 | 28.10 | ||
| 性状 Trait | 地点 Location | 最小值 Min. | 最大值 Max. | 均值 Average | 标准差 SD | 变异系数 CV (%) | 遗传力Heritability |
| 单株荚数 | 哈尔滨 Harbin | 3.90 | 177.00 | 32.32 | 18.86 | 58.35 | 0.09 |
| Pod number per plant | 南阳 Nanyang | 0.10 | 94.00 | 20.23 | 17.58 | 86.93 | |
| 毕节 Bijie | 0.75 | 41.50 | 9.86 | 6.67 | 67.68 | ||
| 乐东 Ledong | 1.80 | 44.50 | 12.29 | 6.26 | 50.93 | ||
| 荚长 | 哈尔滨 Harbin | 4.72 | 18.58 | 10.96 | 2.28 | 20.80 | 0.53 |
| Pod length (cm) | 南阳 Nanyang | 4.39 | 22.98 | 8.19 | 2.20 | 26.80 | |
| 毕节 Bijie | 4.44 | 21.00 | 10.15 | 2.37 | 23.39 | ||
| 乐东 Ledong | 4.00 | 20.40 | 10.50 | 2.77 | 26.37 | ||
| 荚宽 | 哈尔滨 Harbin | 6.20 | 19.80 | 9.94 | 1.84 | 18.53 | 0.67 |
| Pod width (cm) | 南阳 Nanyang | 5.20 | 21.50 | 9.73 | 2.37 | 24.37 | |
| 毕节 Bijie | 5.60 | 21.00 | 10.44 | 1.91 | 18.33 | ||
| 乐东 Ledong | 4.80 | 25.00 | 9.96 | 2.04 | 20.46 | ||
| 单荚粒数 | 哈尔滨 Harbin | 2.60 | 10.00 | 5.67 | 1.26 | 22.22 | 0.30 |
| Seed number per pod | 南阳 Nanyang | 0.70 | 7.50 | 3.69 | 1.17 | 31.63 | |
| 毕节 Bijie | 1.25 | 8.20 | 4.72 | 1.17 | 24.71 | ||
| 乐东 Ledong | 1.40 | 9.60 | 5.12 | 1.06 | 20.66 | ||
| 单株产量 | 哈尔滨 Harbin | 2.88 | 175.46 | 40.55 | 23.20 | 57.22 | |
| Yield per plant (g) | 南阳 Nanyang | — | — | — | — | — | — |
| 毕节 Bijie | 0.30 | 48.86 | 9.67 | 6.74 | 69.63 | ||
| 乐东 Ledong | 1.00 | 40.07 | 12.39 | 6.24 | 50.35 | ||
| 百粒重 Hundred-seed weight (g) | 哈尔滨 Harbin | 10.80 | 90.00 | 35.08 | 13.14 | 37.46 | 0.73 |
| 南阳 Nanyang | 4.00 | 56.90 | 28.34 | 9.06 | 31.98 | ||
| 毕节 Bijie | 9.60 | 85.30 | 33.41 | 13.45 | 40.26 | ||
| 乐东 Ledong | 4.10 | 87.50 | 32.48 | 14.78 | 45.52 |
新窗口打开
采用Microsoft Excel数据分析功能的单因素方差分析, 得到各性状组间和组内的均方差, 根据遗传力公式, 计算各性状的遗传力, 结果见表3。百粒重的遗传力最高0.73, 荚宽次之, 为0.67, 其余各性状的遗传力依次为0.58 (株高)、0.53 (荚长)、0.30 (单荚粒数)、0.30 (主茎节数)、0.09 (单株荚数)和0.01 (播种至开花时间)。
2.3 农艺性状间的相关性分析
在66个性状组合中, 31个同时在测试点呈显著或极显著的相关性(表4), 占46.97%。其中, 21个性状组合呈显著正相关, 10个呈显著负相关。在正相关性状中, 播种至开花与生育期,主茎节数、单荚粒数与株高, 单株产量与单株荚数, 百粒重与荚宽的相关系数较高。在负相关性状中, 百粒重与播种至开花时间、单株荚数、单荚粒数, 主茎分枝数与株高的相关系数较高。在单株产量及其三要素之间, 单株产量与单株荚数、单荚粒数呈极显著正相关, 平均相关系数分别为0.668和0.338; 与百粒重的相关性在各点不一致, 在乐东呈显著正相关, 在毕节呈极显著负相关, 在哈尔滨无相关性; 单株荚数与单荚粒数呈极显著正相关; 百粒重与单株荚数、单荚粒数呈极显著负相关。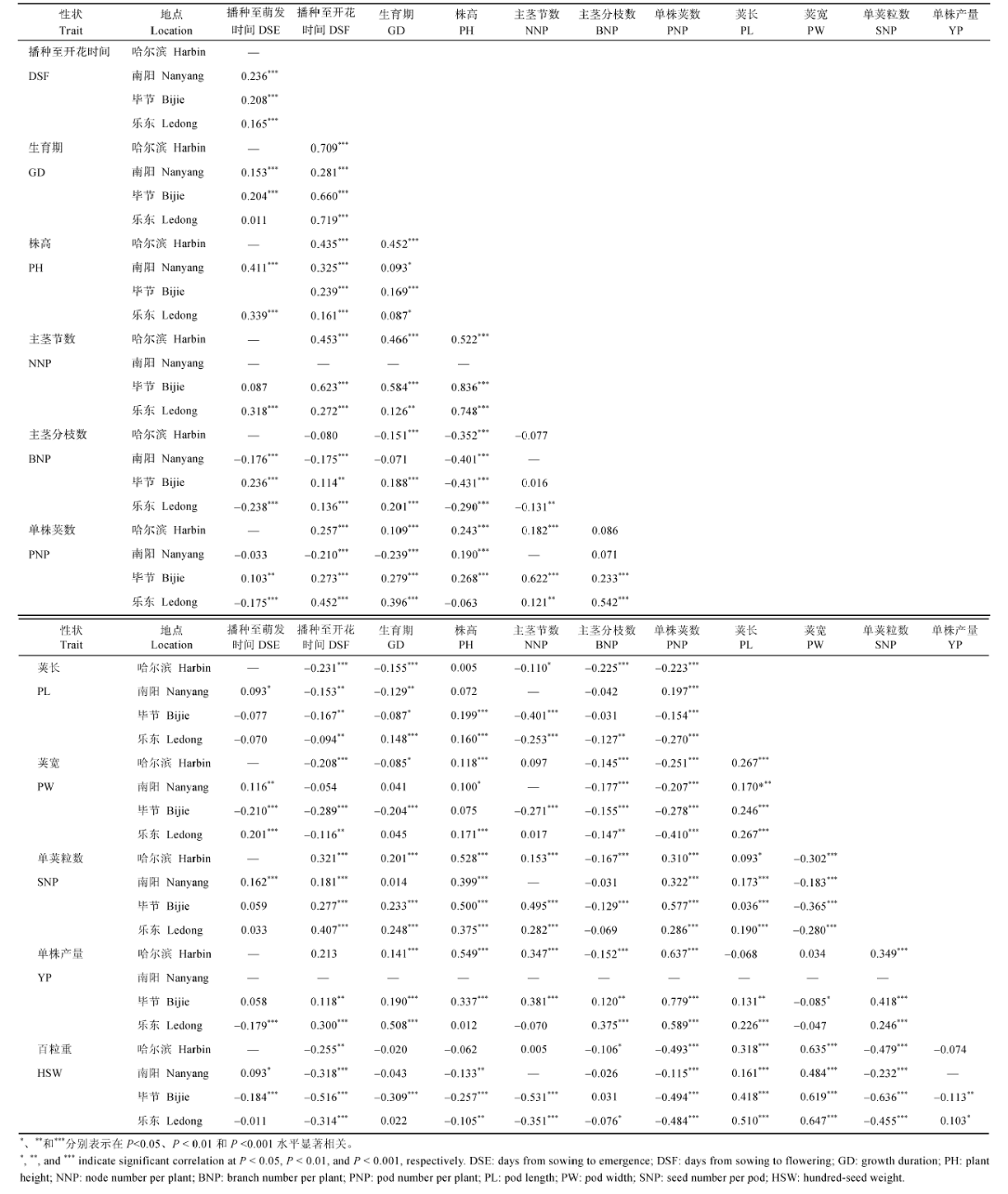 |
2.4 单株产量与农艺性状的通径分析
采用SPSS软件的逐步回归方法, 对哈尔滨、毕节和乐东3个试点的农艺性状进行通径分析, 结果见表5。在哈尔滨, 自变量为单株荚数、株高、百粒重、单荚粒数和播种至开花时间, 与单株产量的通径系数分别为0.691、0.373、0.327、0.121和0.081。在毕节, 自变量为单株荚数、百粒重、荚长和单荚粒数, 与单株产量的通径系数分别为0.906、0.365、0.113和0.124。在乐东, 自变量为单株荚数、百粒重、单荚粒数、生育期、播种至开花时间和荚长, 通径系数分别为0.750、0.468、0.226、0.223、-0.142和0.099。在3个试点, 单株荚数、单荚粒数和百粒重为影响单株产量的共同的自变量。其中, 单株荚数的直接作用最大, 在3个试点的通径系数分别为0.691、0.906和0.750, 单株荚数通过其他性状的间接作用尽管为负值, 但相对较小(-0.054、-0.109和-0.161), 因而使单株荚数对单株产量的贡献较大, 二者的相关系数达到了0.637、0.779和0.589; 其次是百粒重, 在3个试点的通径系数分别为0.327、0.365和0.468, 百粒重通过单株荚数产生较大的负向作用(-0.341、-0.447和-0.363), 使百粒重对单株产量的贡献较小, 二者的相关系数较小, 甚至为负值(-0.074、-0.096和0.103); 单荚粒数的直接作用最小, 在3个试点的通径系数分别为0.121、0.124和0.226, 但单荚粒数通过单株荚数对单株产量产生较大的正向作用(0.214、0.523和0.214), 因而对单株产量的贡献较大, 相关系数分别为0.349、0.388和0.246。Table 5
表5
表5单株产量与其他性状相关系数的分解
Table 5Simple correlation coefficient decomposition between yield per plant and other traits
| 地点 Location | 性状 Trait | 相关系数 Correlation coefficient | 通径系数 Path coefficient | 间接通径系数Indirect path coefficient | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| PPL | HGT | HSW | SPP | MAT | DFF | PL | 合计Total | ||||
| 哈尔滨 | 单株荚数PNP | 0.637 | 0.691 | — | 0.091 | -0.161 | 0.037 | — | -0.021 | — | -0.054 |
| Harbin | 株高PH | 0.549 | 0.373 | 0.168 | — | -0.020 | 0.064 | — | -0.035 | — | 0.177 |
| 百粒重HSW | -0.074 | 0.327 | -0.341 | -0.023 | — | -0.058 | — | 0.021 | — | -0.401 | |
| 单荚粒数SNP | 0.349 | 0.121 | 0.214 | 0.197 | -0.157 | — | — | -0.026 | — | 0.228 | |
| 播种至开花时间DSF | 0.213 | -0.081 | 0.177 | 0.162 | -0.083 | 0.039 | — | — | — | 0.295 | |
| 毕节 | 单株荚数PNP | 0.779 | 0.906 | — | — | -0.180 | 0.071 | — | — | -0.018 | -0.109 |
| Bijie | 百粒重HSW | -0.096 | 0.365 | -0.447 | — | — | -0.079 | — | — | 0.047 | -0.526 |
| 荚长PL | 0.132 | 0.113 | -0.140 | — | 0.153 | 0.004 | — | — | — | 0.017 | |
| 单荚粒数SNP | 0.388 | 0.124 | 0.523 | — | -0.232 | — | — | — | 0.004 | 0.290 | |
| 乐东 | 单株荚数PNP | 0.589 | 0.750 | — | — | -0.227 | 0.065 | 0.092 | -0.064 | -0.027 | -0.161 |
| Ledong | 百粒重HSW | 0.103 | 0.468 | -0.363 | — | — | -0.103 | 0.005 | 0.045 | 0.051 | -0.365 |
| 单荚粒数SNP | 0.246 | 0.226 | 0.214 | — | -0.213 | — | 0.058 | -0.058 | 0.019 | 0.020 | |
| 生育期 GD | 0.508 | 0.233 | 0.297 | — | 0.010 | 0.056 | — | -0.102 | 0.015 | 0.275 | |
| 播种至开花时间DSF | 0.300 | -0.142 | 0.339 | — | -0.147 | 0.092 | 0.168 | — | -0.009 | 0.443 | |
| 荚长PL | 0.226 | 0.099 | -0.203 | — | 0.239 | 0.043 | 0.035 | 0.013 | — | 0.127 | |
新窗口打开
2.5 普通菜豆特异资源筛选
参考《中国食用豆类优异资源》[22]和《普通菜豆生产技术》[23], 筛选到特异资源218份, 其中, 同时具有3个特异性状资源9份, 具有2个特异性状资源53份(表6)。矮秆资源59份, 在3个或4个地点植株较矮。划分标准是哈尔滨株高低于40 cm, 南阳和毕节低于30 cm, 乐东低于25 cm。大粒资源30份, 划分标准是在哈尔滨、毕节和乐东百粒重均高于50 g。早熟资源16份, 在3个或4个地点成熟较早。划分标准是哈尔滨80 d、南阳85 d、毕节100 d、乐东75 d内成熟。多荚资源18份, 在3个或4个地点单株荚数较高。划分标准是哈尔滨大于50个, 南阳大于40个, 毕节和乐东大于20个。多粒资源39个, 在3个或4个地点单荚粒数较多。划分标准是哈尔滨大于7粒, 南阳大于5粒, 毕节、乐东大于6粒。高产资源41份, 在2个或3个地点单株产量较高。划分标准是哈尔滨大于70 g, 毕节和乐东大于20 g。长荚资源38份, 在3个或4个地点荚长较长。划分标准是哈尔滨、乐东大于14 cm, 南阳大于12 cm, 毕节大于13 cm。宽荚资源45份, 在3个或4个地点荚宽较宽。划分标准是毕节大于13 mm, 其他试点大于12 mm。 |
3 讨论
3.1 普通菜豆种质资源的生态适应性
普通菜豆起源于中、南美洲, 对光照、温度相对敏感。本研究686份种质资源, 在4个试点开花结实情况存在较大差异。在哈尔滨, 来自南美的44份野生资源及1份中国云南资源不能正常开花, 16份未成熟的资源13份来自中国云贵高原和四川盆地, 其余3份分别来自中国陕西、哥伦比亚和墨西哥, 说明来自南美及部分中国云贵川的资源不适宜种植在日照时间较长、开花后期气温较低的哈尔滨。在河南南阳, 69份资源不能正常开花, 46份不能正常成熟, 不能结实资源占播种资源总数的16.76%, 这些资源既有中国品种, 也有国外引进品咱。尽管南阳的日照时间短于哈尔滨, 但由于南阳5月之后的气温较高, 在整个生育期, 日最高气温达38°C, 平均在30°C; 日最低气温高达28.0°C, 平均21.2°C, 加之2014年天气干旱, 致使不开花和不能成熟结实的资源数超过哈尔滨, 成熟资源也由于灌浆时间较短, 籽粒普遍不饱满, 平均百粒重仅为28.34 g, 低于哈尔滨(35.08 g)、毕节(33.41 g)和乐东(32.48 g), 说明南阳的气候条件不适合普通菜豆的生长。贵州毕节位于中国的西南部, 日照时间较短, 气候冷凉, 较适合普通菜豆的生长。686份材料中, 只有2份未开花, 39份未正常成熟, 41份未开花或成熟资源中40份为来自国外的野生资源, 只有1份来自中国云南, 说明除了南美野生资源, 国内资源较适宜种植在毕节。乐东县冬天日照时间较短, 在菜豆生育期平均日照长度为11.11 h。供试材料在乐东全部开花结实, 说明乐东的气候条件能满足来自国内和国外普通菜豆资源基本的生长发育需求, 尤其是国外引进种质, 是南美种质良好繁育基地, 也是菜豆种质创新和育种的良好加代场所。
3.2 协调产量性状三要素是提高产量的关键
单株荚数、单荚粒数和百粒重是产量构成三大要素, 本研究结果显示, 单株产量与单株荚数、单荚粒数呈极显著正相关, 与百粒重的相关性在各点不一致, 在乐东二者呈显著正相关, 在毕节呈显著负相关, 在哈尔滨相关不显著。单株荚数、单荚粒数与百粒重呈极显著负相关。前人报道大豆和普通菜豆产量与单株荚数、单株粒数呈显著正相关[24,25,26], 本研究也得到同样的结果。李莹[24]对大豆的研究结果显示, 产量与百粒重呈显著正相关,本研究在乐东的结果与此相吻合。杨光宇[26]对大豆、Hamblin和Evans[26]对普通菜豆的研究结果表明, 单株产量与百粒重呈显著负相关,毕节的结果与此一致。由于各要素之间相互制约, 使得高产菜豆资源大多是多荚、多粒的小粒资源, 大粒资源单株产量相对较低。本研究筛选的41份高产资源中, 2014年在哈尔滨只有F0004395、F0004228和F0002503同时具有高产和大粒的特性, F0000705尽管大粒但产量不高, 其余39份资源的百粒重均小于45 g, 且其中的65.7%百粒重小于30 g。因此, 在普通菜豆育种中, 如何协调三者之间的关系, 提高大粒资源的产量是亟待解决的问题。理论上看, 可以直接或间接通过增加单株荚数、单荚粒数等来提高产量, 但在育种实践中难度很大[27,28,29,30]。Nienhuis和Singh [31] 1988年通过对80个亲本及其200个F2群体后代的产量及其组成成分的遗传分析, 认为以产量本身作为选择来改进产量是可行的。直接以产量选择使产量增加4.3%, 百粒重增加0.21%; 直接以百粒重作为选择目标使百粒重增加11.76%, 产量增加0.28%; 直接以荚选择会导致产量、单荚粒数和百粒重降低; 而直接以单荚粒数选择会导致荚数减少、百粒重降低, 但种子产量增加0.37%。在今后的育种中, 应多配置杂交组合, 将大量的杂交组合后代播种在在不同的试验点, 从中选择单株荚数、单荚粒数、百粒重协调的高产资源, 尤其是高产大粒型资源, 培育大粒高产品种。4 结论
乐东、毕节分别是南美引进菜豆资源、国内菜豆资源的最佳繁殖地, 哈尔滨是普通菜豆平均单株单产最高的试验地; 数量性状在试点间的平均变异系数介于8.11%~70.83%之间。株高的变异系数最大、单株荚数次之。百粒重、荚宽、株高和荚长遗传力较高, 分别为0.73、0.67、0.58和0.53。51.65%的性状组合同时在测试点呈显著或极显著的相关性, 单株产量与单株荚数、单荚粒数极显著正相关, 平均相关系数分别为0.668 和 0.338。单株荚数对单株产量直接作用最大、百粒重次之、单荚粒数最小。筛选出特异资源218份, 其中, 同时具有3种特异性状资源9份, 2种特异性状资源53份。The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | . |
| [2] | . |
| [3] | . |
| [4] | . |
| [5] | . |
| [6] | . |
| [7] | . |
| [8] | . |
| [9] | . |
| [10] | . |
| [11] | |
| [12] | |
| [13] | . |
| [14] | . |
| [15] | . |
| [16] | . |
| [17] | . |
| [18] | . . |
| [19] | . . |
| [20] | . . |
| [21] | |
| [22] | |
| [23] | |
| [24] | . . |
| [25] | . . |
| [26] | . |
| [27] | . |
| [28] | . |
| [29] | . |
| [30] | . |
| [31] | . |
