Regeneration Capacity Evaluation of Some Largely Popularized Wheat Varieties in China
ZHANGWei**, YINMi-Qi**, ZHAOPei, WANGKe, DULi-Pu, YEXing-Guo* 1 Institute of Crop Sciences, Chinese Academy of Agricultural Sciences / National Key Facility for Gene Resources and Genetic Improvement, Beijing 1000812 Hainan Sugarcane Breeding Station, Guangzhou Sugarcane Industry Research Institute, Sanya 572025, Hainan, China 通讯作者:叶兴国, E-mail: yexingguo@caas.cn ** 同等贡献(Contributed equally to this work) 收稿日期:2017-03-26 接受日期:2017-09-10 网络出版日期:2018-02-12 版权声明:2018作物学报编辑部作物学报编辑部 基金资助:本研究由国家重点研发计划项目(2016YFD0102001)和国家转基因生物新品种培育重大专项(2016ZX08010004)资助
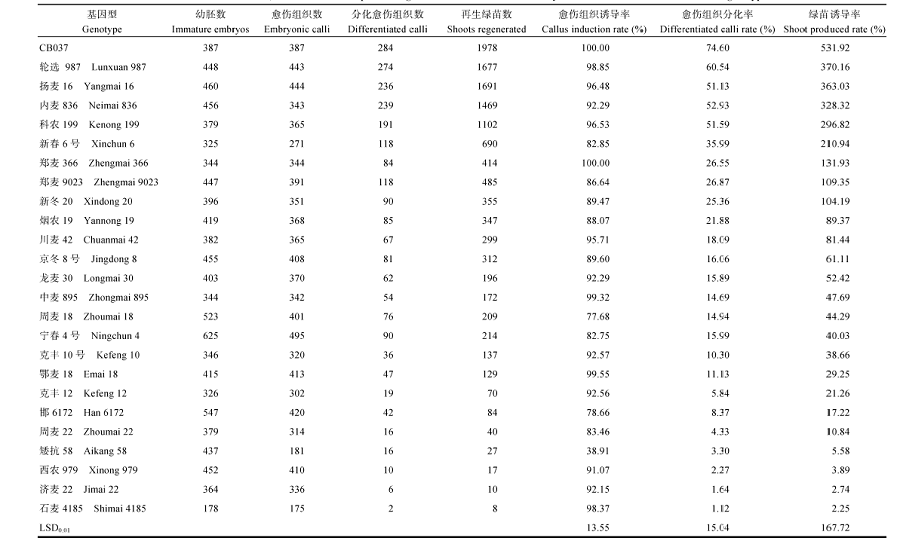
关键词:小麦;花药;幼胚;成熟胚;组织培养 Abstract Strong genotype dependence exists in wheat doubling haploid and genetic engineering breeding, in which high regeneration ability is a main restrain. In this study, we evaluated the regeneration abilities of various explants of 24 commercial popularized wheat varieties and a new breeding line (CB037) with high powdery mildew resistance. The explants used in the two-year experiment were anther, immature embryo, and mature embryo. The regeneration potential was assessed based on callus induction rate, callus differentiation rate, and shoot induction rate. The plantlet regeneration rates of the all genotypes tested were 0-41.75% for anther culture, 2.25%-531.92% for immature embryo culture, and 3.24%-84.34% for mature embryo culture, showing significant differences among genotypes. Generally, immature embryos had stronger regeneration ability (119.79%) than mature embryos (36.23%) and anthers (4.91%). Among all the used genotypes, CB037 showed the highest regeneration rates for the three explant types. Lunxuan 987, Yangmai 16, Neimai 836, Kenong 199, Xinchun 6, Zhengmai 366, Zhengmai 9023, Xindong 20, Yannong 19, and Chuanmai 42 exhibited ideal regeneration abilities in immature embryo culture; Xinchun 6, Jingdong 8, Shimai 4185, Kenong 199, and Luanxun 987 performed good in mature embryo culture; and Shimai 4185 and Han 6172 were characterized with high regeneration rate of green plantlet in anther culture. Plant regeneration efficiency of wheat was closely associated with genotype and explant type. The same explant type from different genotypes showed significantly different regeneration abilities, and different explants of the same genotype showed different regeneration abilities. However, there was no correlation among regeneration abilities of the three types of explants. The selected genotypes with high regeneration potential are recommended for application in genetic and cell engineering breeding of wheat.
在小麦孕穗期, 取处于单核靠边期的小麦幼穗, 用塑料袋包好置4℃冰箱预处理3 d。接种前用70%酒精擦拭茎秆以表面消毒, 在洁净工作台中用镊子剥去颖壳, 取出花药接种在W14愈伤组织诱导培养基上(见附表1), 先在生化培养箱中30℃、黑暗条件下预培养3 d, 然后转到28℃、黑暗下培养30~45 d。将产生的直径1 mm左右的愈伤组织转移到1/2 MSNK分化培养基(见附表1)上, 在(25±1)℃、70 µmol m-2 s-1、16 h光照条件下培养20~25 d分化植株[2, 11]。 Compositions of the media used in this study for callus induction and differentiation 附表1 附表1本研究用培养基配方 Compositions of the media used in this study for callus induction and differentiationCorrelation coefficient between tissue culture traits of different wheat explants
小麦组织培养效率与基因型和外植体类型密切相关。25个小麦品种(系)中, CB037的花药、幼胚和成熟胚3种外植体组织培养植株再生效率均最高; 轮选987、扬麦16、内麦836、科农199、新春6号、郑麦366、郑麦9023、新冬20、烟农19和川麦42幼胚培养植株再生能力较强; 新春6号、京冬8号、石麦4185、科农199和轮选987成熟胚培养植株再生率较高; 石麦4185和邯6172花药培养绿苗诱导率较高。这些不同外植体再生率较高的小麦基因型可分别用于小麦转基因育种和单倍体育种。 The authors have declared that no competing interests exist. 作者已声明无竞争性利益关系。
Ye X G. Xu H J, Du LP, He GY, WangK, Lin ZS.Establishment and application of large-scale transformation systems in wheat .Sci Agric Sin, 2014, 47: 4155-4171 (in Chinese with English abstract) [本文引用: 5]
Han XF, Tao LL, Yin GX, Liu XL, Du LP, Wei YQ, Yan YM, Ye XG.Effect of genotype and growing environment on anther culture in wheat .Acta Agron Sin, 2010, 36: 1209-1215 (in Chinese with English abstract) [本文引用: 10]
[3]
She MY, Yin GX, Li JR, LiX, Du LP, Ma WJ, Ye XG.Efficient Regeneration potential is closely related to auxin exposure time and catalase metabolism during the somatic embryogenesis of immature embryos inTriticum aestivum L . Mol Biotechnol, 2013, 54: 451-460 [本文引用: 4]
[4]
Mathias RJ, FukuiK, LawC.Cytoplasmic effects on the tissue culture response of wheat ( Triticum aestivum) callus. Theor Appl Genet, 1986, 72: 70-75 [本文引用: 1]
[5]
HeD, YangY, ScottK.A comparison of scutellum callus and epiblast callus induction in wheat: the effect of genotype, embryo age and medium .Plant Sci, 1988, 57: 225-233 [本文引用: 1]
[6]
StoberA, HessuD.Spike pretreatments, anther culture conditions, and anther culture response of 17 German varieties of spring wheat ( Triticum aestivum L.). Plant Breed, 1997, 116: 443-447 [本文引用: 1]
[7]
Mendoza MG, Kaeppler HF.Auxin and sugar effects on callus induction and plant regeneration frequencies from mature embryos of wheat (Triticum aestivum L.) . In Vitro Cell Dev-Pl, 2002, 38: 39-45 [本文引用: 1]
[8]
VarshneyA, AltpeterF.Stable transformation and tissue culture response in current European winter wheats ( Triticum aestivum L.). Mol Breed, 2002, 8: 295-309 [本文引用: 1]
[9]
TurhanH, BaserI.Callus induction from mature embryo of winter wheat ( Triticum aestivum L.). Asian J Plant Sci, 2004, 3: 17-19 [本文引用: 1]
[10]
SharmaV, HänschR, MendelR, SchulzeJ.Influence of picloram and thidiazuron on high frequency plant regeneration in elite cultivars of wheat with long‐term retention of morphogenecity using meristematic shoot segments . Plant Breed, 2005, 124: 242-246 [本文引用: 1]
Ye XG, Xu HJ, Xu QF, Du LP, Li ZW.Genetic analysis and combining ability evaluation of the anther culture response in common wheat .Sci Agric Sin, 1997, 30(6): 49-54 (in Chinese with English abstract) [本文引用: 5]
[12]
MachiiH, MizunoH, HirabayashiT, LiH, HagioT.Screening wheat genotypes for high callus induction and regeneration capability from anther and immature embryo cultures .Plant Cell, Tiss Org, 1998, 53: 67-74 [本文引用: 1]
[13]
ShahM, KhalidQ, KhanU, ShahS, ShahS, HassanA, PervezA, OliveiraV, CaxitoF, GomesK.Variation in genotypic responses and biochemical analysis of callus induction in cultivated wheat . Genet Mol Res, 2009, 8: 783-793
[14]
Yin GX, Wang YL, She MY, Du LP, Xu HJ, Ma JX, Ye XG.Establishment of a highly efficient regeneration system for the mature embryo culture of wheat . Agric Sci China, 2011, 10: 9-17 [本文引用: 3]
[15]
ZamaniI, Gouli VavdinoudiE, KovacsG, XyniasI, RoupakiasD, BarnabasB.Effect of parental genotypes and colchicine treatment on the androgenic response of wheat F1 hybrids .Plant Breed, 2003, 122: 314-317 [本文引用: 1]
[16]
Ozias AP, Vasil IK.Plant regeneration from cultured immature embryos and inflorescences of Triticum aestivum L.(wheat): evidence for somatic embryogenesis . Protoplasma, 1982, 110: 95-105 [本文引用: 1]
[17]
Bi RM, KouM, Chen LG, Mao SR, Wang HG.Plant regeneration through callus initiation from mature embryo ofTriticum .Plant Breed, 2007, 126: 9-12 [本文引用: 1]
[18]
ArzaniA, Mirodjagh SS.Response of durum wheat cultivars to immature embryo culture, callus induction andin vitro salt stress . Plant Cell, Tiss Org, 1999, 58: 67-72 [本文引用: 1]
[19]
SzakácsE, KovácsG, PaukJ, BarnabásB.Substitution analysis of callus induction and plant regeneration from anther culture in wheat (Triticum aestivum L.) . Plant Cell Rep, 1988, 7: 127-129 [本文引用: 1]
[20]
Liu WG, Zheng MY, Enrique AP, Calvin FK.Highly efficient doubled-haploid production in wheat (Triticum aestivum L.) via induced microspore embryogenesis . Crop Sci, 2002, 42: 686-692 [本文引用: 1]
[21]
Tuvesson I KD, PedersenS, Andersen SB. Nuclear genes affecting albinism in wheat (Triticum aestivum L.) anther culture . Theor Appl Genet, 1989, 78: 879-883 [本文引用: 1]
Jia HY, YuJ, Yi DL, ChengY, Xu WQ, Zhang LX, Ma ZQ.Chromosomal intervals responsible for tissue culture response of wheat immature embryos . Plant Cell, Tiss Org, 2009, 97: 159-165 [本文引用: 1]
[24]
Jia HY, Yi DL, YuJ, Xue SL, XiangY, Zhang CQ, Zhang ZZ, ZhangL, Ma ZQ.Mapping QTLs for tissue culture response of mature wheat embryos . Mol Cells, 2007, 23: 323-330 [本文引用: 6]
[25]
PellegrineschiA, Noguera LM, SkovmandB, Brito RM, VelazquezL, Salgado MM, HernandezR, Warburton ML, Hoisington DA.Identification of highly transformable wheat genotypes for mass production of fertile transgenic plants .Genome, 2002, 45: 421-430 [本文引用: 1]
[26]
KhannaH, DaggardG.Agrobacterium tumefaciens-mediated transformation of wheat using a superbinary vector and a polyamine-supplemented regeneration medium . Plant Cell Rep, 2003, 21: 429-436
[27]
Greer MS, KovalchukI, EudesF.Ammonium nitrate improves direct somatic embryogenesis and biolistic transformation ofTriticum aestivum . New Biotechnol, 2009, 26: 44-52
[28]
IshidaY, TsunashimaM, HieiY, KomariT.Wheat (Triticum aestivum L.) transformation using immature embryos. In: Agrobacterium Protocols: Volume 1. Methods in Molecular Biology vol. 1223, (Wang K ed) New York: Springer Science+ Business Media, 2015. pp 189-198 [本文引用: 2]
[29]
ZhouH, Berg JD, Blank SE, Chay CA, ChenG, Eskelsen SR, Fry JE, HoiS, HuT, Isakson PJ, Lawton MB, Metz SG, Rempel CB, Ryerson DK, Sansone AP, Shook AL, Starke RJ, Tichota JM, Valenti SA.Field efficacy assessment of transgenic roundup ready wheat .Crop Sci, 2003, 43: 1072-1075 [本文引用: 1]
[30]
StokstadE.Biotechnology-Monsanto pulls the plug on genetically modified wheat .Science, 2004, 304: 1088-1089 [本文引用: 1]
Ye XG, Xu HJ, Zhao LL, Du LP.Studies on improving wheat cultivars by tissue culture .Acta Agron Sin, 1998, 24: 310-314 (in Chinese with English abstract) [本文引用: 1]
Ye XG, Xu HJ, Du LP, Xin ZY.Studies on the factors influencing the efficiency of wheat transformation .Sci Agric Sin, 2001, 34: 128-132 (in Chinese with English abstract) [本文引用: 2]
[33]
ZhangW, Wang XM, FanR, Yin GX, WangK, Du LP, Xiao LL, Ye XG.Effects of inter-culture, arabinogalactan proteins, and hydrogen peroxide on the plant regeneration of wheat immature embryos .J Integr Agric, 2015, 14: 11-19 [本文引用: 3]
Ye XG, She MY, WangK, Du LP, Xu HJ.Identification, cloning, and potential application of genes related to somatic embryogenesis in plant tissue culture . Acta Agron Sin, 2012, 38: 191-201 (in Chinese with English abstract) [本文引用: 1]
[35]
Wang XM, WangK, Li JR, Du LP, Li JR, Xu HJ, Ye XG.Effects of environmental temperature on the regeneration frequency of the immature embryos of wheat (Triticum aestivum L.) . J Integr Agric, 2014, 13: 722-732 [本文引用: 1]
[36]
ChauhanH, KhuranaP.Use of doubled haploid technology for development of stable drought tolerant bread wheat (Triticum aestivum L.) transgenics . Plant Biotechnol J, 2011, 9: 408-417 [本文引用: 1]
[37]
Wang YL, Xu MX, Yin GX, Tao LL, Wang DW, Ye XG.Transgenic wheat plants derived fromAgrobacterium-mediated transformation of mature embryo tissues . Cereal Res Commun, 2009, 37: 1-12 [本文引用: 1]
[38]
Li JR, Ye XG, An BY, Du LP, Xu HJ.Genetic transformation of wheat: current status and future prospects .Plant Biotechnol Rep, 2012, 6: 183-193 [本文引用: 2]
[39]
Tao LL, Yin GX, Du LP, Shi ZY, She MY, Xu HJ, Ye XG.Improvement of plant regeneration from immature embryos of wheat infected byAgrobacterium tumefaciens . Agric Sci China, 2011, 10: 317-326
WangK, Liu HY, Du LP, Ye XG.Generation of marker-free transgenic hexaploid wheat via anAgrobacterium-mediated co-transformation strategy in commercial Chinese wheat varieties . Plant Biotechnol J, 2017, 15: 614-623 [本文引用: 1]
Zhang YL, Wang MJ, ZhangY, Chu CP, Lin ZS, Xu QF, Ye XG, ChenX, Zhang XS.Development and application of functional markers specific to powdery mildew resistance on chromosome arm 6VS from different origins ofHaynaldia villosa . Acta Agron Sin, 2012, 38: 1827-1832 (in Chinese with English abstract)
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}