Effect of Low Temperature in Spring on Yield and Photosynthetic Characteristics of Wheat
WANGRui-Xia1, YANChang-Sheng2, ZHANGXiu-Ying2, SUNGuo-Zhong2, QIANZhao-Guo1, QIXiao-Lei1, MOUQiu-Huan1, XIAOShi-He2,*, 1 Tai’an Academy of Agricultural Sciences, Tai’an 271000, Shandong, China; 2 National Key Facility for Crop Gene Resources and Genetic Improvement / Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081, China 通讯作者:肖世和, E-mail: xiaoshihe@caas.cn 收稿日期:2017-02-4 接受日期:2017-09-10 网络出版日期:2018-02-12 版权声明:2018作物学报编辑部作物学报编辑部 基金资助:本研究由国家现代农业产业技术体系建设专项(CARS-03-21), 国家公益性行业科研专项(201203033)和山东省泰安市科研专项(20123034)资助 作者简介: -->ruixiawangli@163.com
关键词:小麦;低温;产量;光合荧光参数;抗寒性 Abstract Low temperature (LT) stress in spring occurs frequently in the Huang-Huai Rivers Valley, resulting in yield loss of wheat. The objectives of this study were to understand the physiological responses of wheat cultivars to LT and determine suitable physiological indicator(s) for cold-tolerance selection. Three cultivars, Taishan 6426, Taishan 4033, and Jimai 22, were exposed to LT stress at standing, jointing and booting stages, and the yield-related traits, photosynthetic parameters as well as leaf cell structure were compared among different cultivars. The agronomic traits of different cultivars changed in similar trends after LT treatment with the consistent result across years. Plant height received small influence of LT, especially LT at standing stage. In contrast, spike number per plant decreased significantly compared with the control (normal temperature), but the reduced percentage varied across years. Grain number per spike and yield per plant showed significantly declined trends with delaying LT stress, and the maximum decrements were observed in the treatment of LT at booting stage. The grain number per spike was 90.5%-93.3% (2012-2013) and 91.9%-93.6% (2013-2014) lower, and the yield per plant was 87.9%-97.3% (2012-2013) and 91.5%-97.8% (2013-2014) lower in LT treatment at booting than in the control. Booting stage was secondly sensitive to LT in yield, whereas jointing stage was less influenced by LT. Grain yield loss under LT stress mainly resulted from the decrease of grain number per spike. Under LT at jointing or booting stage, the chlorophyll content decreased in the three cultivars, but the decrement varied across cultivars. Moreover, the photosynthetic rate, transpiration rate and stomata conductance decreased significantly and the intercellular CO2 concentration increased greatly in the three cultivars, and those in Taishan 6426 had the minimum variations. LT also had inhibition in chlorophyll fluorescence parameters of wheat. For example, the Fv/Fm value was significantly lower in the treatment of LT at jointing or booting stage than in the control except for Taishan 6426. When exposed to LT stress, leaf epidermic cells were severely damaged, showing their loose arrangement and irregular structures of guard cell and accessory cell. Our results indicate that photosynthetic rate, transpiration rate, stomata conductance, intercellular CO2 concentration and Fv/Fm can be used as indicators to identify tolerance of wheat cultivars to spring coldness.
Keywords:wheat;low temperature;yield;photosynthetic and fluorescent parameters;cold resistance -->0 PDF (3096KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 王瑞霞, 闫长生, 张秀英, 孙果忠, 钱兆国, 亓晓蕾, 牟秋焕, 肖世和. 春季低温对小麦产量和光合特性的影响[J]. 作物学报, 2018, 44(02): 288-296 https://doi.org/10.3724/SP.J.1006.2018.00288 WANGRui-Xia, YANChang-Sheng, ZHANGXiu-Ying, SUNGuo-Zhong, QIANZhao-Guo, QIXiao-Lei, MOUQiu-Huan, XIAOShi-He. Effect of Low Temperature in Spring on Yield and Photosynthetic Characteristics of Wheat[J]. Acta Agronomica Sinica, 2018, 44(02): 288-296 https://doi.org/10.3724/SP.J.1006.2018.00288 低温冷(冻)害是黄淮海麦区频发的自然灾害之一, 严重影响并制约小麦的生长发育及产量的提高[1,2]。低温胁迫后, 植物体通过形态结构、内含物质和光合作用等一系列复杂的生理生化变化应对低温环境[3,4,5,6], 但小麦的耐寒机制是受多因素影响的复杂生理过程, 不同品种的抗逆机制不尽相同[7,8,9]。研究表明, 小麦多个生理生化指标与耐寒性显著相关, 可作为抗寒能力的评价指标, 如丙二醛(MDA)[7]、可溶性糖[7]、脯氨酸[7]或羟脯氨酸[10]、糖蛋白[10]、寒胁迫蛋白[11]、电导率[11]和细胞膜透性[12,13]。刘炜等[14]指出, 低温逆境下小麦Ca2+-ATPase的活性及其稳定性是决定植株抗寒能力的关键。但利用这些指标鉴定抗寒性, 均存在测定步骤繁琐的问题, 不适合进行大规模样品的快速筛选。 光合作用是植物物质代谢和能量转化的源泉[15]。在自然条件下, 叶绿素荧光和光合作用相辅相成, 植物在进行光合作用的原初反应能量转换时, 叶绿体色素在光能的吸收、传递和转换过程中起着关键作用。叶绿素荧光参数具有反映光合作用“内在性”的特点, 被称为活体测定叶片光合功能的快速、无损伤探针[16,17]。通过测定光合过程中的荧光特性参数, 可以了解植物的生长发育以及对逆境胁迫、病虫害等的生理响应[18,19]。王晓楠等[20]认为, 在低温驯化及封冻阶段, 可以利用叶绿素荧光参数来鉴定小麦品种的抗寒性。 目前, 针对小麦生理生化特性与抗寒性的关系已有很多研究报道, 但多数试验只是关注小麦某一生育期的耐寒性。例如, 姜丽娜等[7]在拔节期进行低温处理, 并利用小麦叶片的生理响应来评价抗寒性; 张志伟等[8]在孕穗期研究不同类型小麦品种低温生理反应及其抗寒性, 张军等[9]研究拔节期低温胁迫对小麦生理生化特性和产量的影响。结合起身期、拔节期和孕穗期综合评定小麦春季耐寒性的研究尚未见报道。本试验以抗寒性不同的小麦品种(系)泰山6426、泰山4033和济麦22为试验材料, 研究了不同小麦品种在起身期、拔节期和孕穗期低温处理条件下的产量性状、光合生理指标及叶片细胞结构差异, 以期为耐低温品种的选育和利用提供理论依据。
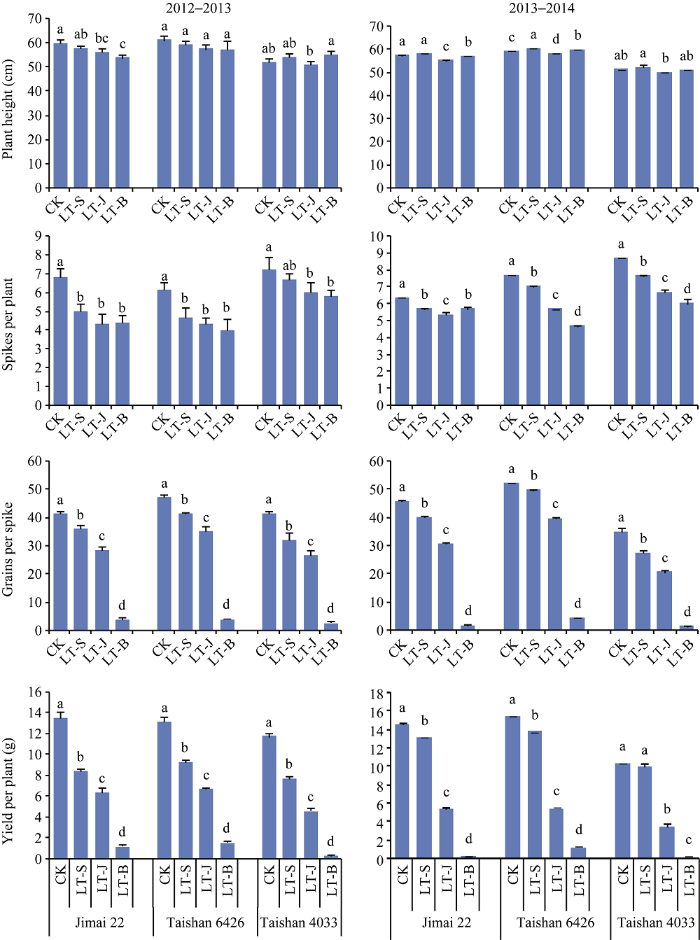
不同低温处理对小麦株高有一定影响, 品种间、年度间存在差异, 总体来看, 起身期低温对株高影响较小, 拔节期和孕穗期低温可导致株高不同程度的下降(图1-A, B)。 显示原图|下载原图ZIP|生成PPT 图1不同时期低温处理对小麦品种产量构成因素的影响 CK: 对照; LT-S: 起身期低温; LT-J: 拔节期低温; LT-B: 孕穗期低温处理。误差线上不同字母表示处理间差异显著(P < 0.05)。 -->Fig. 1Variation of yield components after low temperature stress at different stages CK: control; LT-S: low temperature at standing stage; LT-J: low temperature at jointing stage; LT-B: low temperature at booting stage. Different letters above error bars indicate significant difference among treatments at P < 0.05. -->
由低温引起的叶绿素含量下降, 在品种间表现不完全一致, 孕穗期低温对泰山6426的叶绿素含量无显著影响, 而另2个品种则表现为显著或极显著低于对照; 4个光合特性参数中, 低温胁迫下光合速率、蒸腾速率和叶片气孔导度较对照下降, 胞间CO2浓度则有较大幅度的升高, 除2013—2014年度济麦22孕穗期低温处理的蒸腾速率与常温对照无显著差异外, 其余3个品种的各项参数指标均与对照有显著或极显著差异(表1)。 Table 1 表1 表1低温处理后各品种的光合特性差异 Table 1Variation of photosynthetic characteristics after low temperature stress
年份 Year
品种 Cultivar
参数 Parameter
CK1
LT-J
CK2
LT-B
2012-2013
济麦22 Jimai 22
Chl (mg mg-1)
63.40
55.10**
62.90
54.60**
Pn (μmol m-2 s-1)
17.30
8.44**
10.22
2.35**
E (mmol m-2 s-1)
1.93
0.68**
0.94
0.67*
Cleaf (mmol m-2 s-1)
187.30
73.50**
74.30
48.60**
Cint (μmol mol-1)
97.90
340.50**
260.50
337.30**
泰山6426 Taishan 6426
Chl (mg mg-1)
61.20
54.80**
57.50
54.20
Pn (μmol m-2 s-1)
15.96
5.14**
10.35
2.39**
E (mmol m-2 s-1)
0.65
0.31**
0.76
0.51*
Cleaf (mmol m-2 s-1)
131.10
69.70**
63.40
49.50**
Cint (μmol mol-1)
164.80
353.80**
269.6
356.20**
泰山4033 Taishan 4033
Chl (mg mg-1)
58.40
49.90**
56.10
45.00**
Pn (μmol m-2 s-1)
20.01
5.17**
9.07
1.89**
E (mmol m-2 s-1)
0.60
0.11**
0.60
0.32*
Cleaf (mmol m-2 s-1)
134.50
64.80**
59.00
45.90**
Cint (μmol mol-1)
138.60
375.80**
234.10
354.80**
2013-2014
济麦22 Jimai 22
Chl (mg mg-1)
65.80
57.10**
60.90
57.40*
Pn (μmol m-2 s-1)
26.50
4.77**
27.66
5.54**
E (mmol m-2 s-1)
1.62
0.80**
0.80
0.78
Cleaf (mmol m-2 s-1)
305.90
83.60**
66.20
53.80**
Cint (μmol mol-1)
280.40
341.20**
271.10
289.60**
泰山6426 Taishan 6426
Chl (mg mg-1)
62.30
58.00**
59.90
58.20
Pn (μmol m-2 s-1)
18.07
4.92**
20.79
0.33**
E (mmol m-2 s-1)
1.24
0.81**
0.45
0.34**
Cleaf (mmol m-2 s-1)
193.90
95.40**
58.80
43.00**
Cint (μmol mol-1)
279.00
343.50**
239.80
436.70**
泰山4033 Taishan 4033
Chl (mg mg-1)
57.60
51.80**
57.50
47.10**
Pn (μmol m-2 s-1)
22.45
1.34**
21.56
0.28**
E (mmol m-2 s-1)
1.14
0.56**
1.82
1.27**
Cleaf (mmol m-2 s-1)
180.60
68.60**
57.20
41.30**
Cint (μmol mol-1)
229.10
406.90**
161.30
401.40**
CK1 and CK2 are the measured data of control at jointing and booting stages, respectively. LT-J and LT-B indicate low temperature at jointing and booting stages, respectively. * and ** indicate significant difference between low-temperature treatment and the control at P < 0.05 and P < 0.01, respectively. Chl: chlorophyll content; Pn: photosynthetic rate; E: traspiration ratio; Cleaf: stomatal conductance of leaf; Cint: intercellular CO2 concentration.CK1和CK2分别为对照在拔节期和孕穗期的测定值; LT-J和LT-B分别表示拔节期和孕穗期低温处理测定值。*和**分别表示低温处理与对照存在显著(P < 0.05)和极显著(P < 0.01)差异。Chl: 叶绿素含量; Pn: 光合速率; E: 蒸腾速率; Cleaf: 气孔导度; Cint: 胞间CO2浓度。 新窗口打开 光合参数变化最明显的是泰山4033, 变化幅度最小的为泰山6426。例如, 2012—2013年度, 光合速率、蒸腾速率和叶片气孔导度的降幅, 拔节期低温处理依次为74.2%、81.7%和51.8%, 孕穗期低温处理依次为79.2%、46.7%和22.3%, 而胞间CO2浓度在拔节期和孕穗期低温处理中分别比对照高171.2%和51.6%。同期, 泰山6426拔节期低温处理光合速率、蒸腾速率和叶片气孔导度分别降低67.8%、52.3%和46.8%, 孕穗期低温处理的降幅依次为76.9%、32.9%和21.9%, 胞间CO2浓度在拔节期和孕穗期低温处理中分别比对照高114.7%和32.1%。说明泰山4033的光合特性受低温影响大, 而泰山6426在低温胁迫后自我调节能力较强, 抗寒性较好。
2.3 不同时期低温处理对荧光参数的影响
不同时期低温处理后, 各品种最大荧光产量显著降低, 差异均达极显著水平(表2)。除泰山6426外, 其余2个品种的Fv/Fm也均显著降低, 如济麦22拔节期低温处理后的下降幅度分别为5.3%和9.0%, 而泰山4033下降幅度分别达到15.9%和13.8%。济麦22孕穗期下降幅度为24.1%和16.7%, 而泰山4033为20.3%和19.7%。泰山6426低温处理后的Fv/Fm下降程度最小, 说明其受低温伤害的程度小, 抗寒性最强。 Table 2 表2 表2低温处理后各品种荧光特性 Table 2Variation of fluorescent characteristics after low temperature stress
年份 Year
品种 Cultivar
参数 Parameter
CK1
LT-J
CK2
LT-B
2012-2013
济麦22 Jimai 22
最大荧光产量 Fm
849.00
682.00**
1121.00
735.00**
最小荧光产量 Fo
203.00
194.00*
239.00
294.00**
光化学猝灭系数 NPQ
1.65
0.95**
1.76
0.83**
PSII最大光化学效率Fv/Fm
0.76
0.72**
0.79
0.60**
泰山6426 Taishan 6426
最大荧光产量 Fm
728.00
707.00**
886.00
647.00**
最小荧光产量 Fo
227.00
231.00
315.00
243.00**
光化学猝灭系数 NPQ
1.37
1.30
1.27
0.90**
PSII最大光化学效率Fv/Fm
0.69
0.67
0.64
0.62
泰山4033 Taishan 4033
最大荧光产量 Fm
673.00
591.00**
771.00
694.00**
最小荧光产量 Fo
210.00
246.00**
281.00
371.00**
光化学猝灭系数 NPQ
0.70
0.48**
0.73
0.97**
PSII最大光化学效率Fv/Fm
0.69
0.58**
0.64
0.51**
2013-2014
济麦22 Jimai 22
最大荧光产量 Fm
1114.00
928.00**
1034.00
735.00**
最小荧光产量 Fo
370.00
358.00*
186.00
294.00**
光化学猝灭系数 NPQ
0.67
0.22**
0.87
0.83**
PSII最大光化学效率Fv/Fm
0.67
0.61**
0.72
0.60**
泰山6426 Taishan 6426
最大荧光产量 Fm
1101.00
1027.00**
794.00
647.00**
最小荧光产量 Fo
508.00
403.00**
287.00
243.00**
光化学猝灭系数 NPQ
0.92
0.57**
0.50
0.90**
PSII最大光化学效率Fv/Fm
0.63
0.61
0.64
0.62
泰山4033 Taishan 4033
最大荧光产量 Fm
2079.00
1419.00**
1076.00
874.00**
最小荧光产量 Fo
257.00
328.00**
345.00
444.00**
光化学猝灭系数 NPQ
0.70
0.62**
0.24
0.30*
PSII最大光化学效率Fv/Fm
0.80
0.69**
0.61
0.49**
CK1 and CK2 are the measured data of control at jointing and booting stages, respectively. LT-J and LT-B indicate low temperature at jointing and booting stages, respectively. * and ** indicate significant difference between low-temperature treatment and the control at P < 0.05 and P < 0.01, respectively.CK1和CK2分别为对照在拔节期和孕穗期的测定值; LT-J和LT-B分别表示拔节期和孕穗期低温处理测定值。*和**分别表示低温处理与对照存在显著(P < 0.05)和极显著(P < 0.01)差异。 新窗口打开 在Fm、Fv/Fm降低的同时, 泰山4033还伴随Fo的上升。在拔节期低温处理后, Fo上升幅度分别为17.1%和27.6%、而孕穗期低温处理上升幅度达32.0%和28.7%, 表明泰山4033的PSII遭受了破坏, 尤以孕穗期伤害最为严重(表2)。
2.4 低温处理后各品种的叶表组织结构变化
低温处理后, 各品种的叶片表面组织结构均发生了一定程度的变化, 且变化程度随品种而异。主要表现为低温伤害后表皮叶肉细胞破坏严重, 排列趋于松散, 哑铃状的保卫细胞和半圆状的副卫细胞变形, 气孔变大(图2)。 显示原图|下载原图ZIP|生成PPT 图2拔节期、孕穗期低温处理各品种叶片组织结构变化 CK1和CK2分别为拔节期和孕穗期对照; LT-J和LT-B分别在拔节期和孕穗期低温处理。EP: 表皮; MC: 叶肉细胞; GC: 保卫细胞; SAC: 副卫细胞; STO: 气孔。 -->Fig. 2Leaf tissue structure of different wheat cultivars after low temperature stress at jointing and booting stages CK1 and CK2 are the controls at jointing and booting stages, respectively. LT-J and LT-B indicate low temperature at jointing and booting stages, respectively. EP: epidermis; MC: mesophyll cell; GC: guard cell; SAC: semicide-accessory cell; STO: stomata. -->
Fang SB, Yang JJ, Zhou GS.Change trend distributive characteristics of agrometeorological disasters in China in recent 30 years .J Nat Dis, 2011, 20: 69-73 [本文引用: 1]
[2]
Wang JF, Zhu YY, Liu HP.Main agrometeorological disasters and their impact in recent 28 years in Henan .Meteorol Environ Sci, 2007, 30: 9-11 [本文引用: 1]
[3]
Cao HM, Shi ZM, Zhou XB, Lei PZ, Dong SG.A review on response of plant to low temperature and its cold resistance .Chin J Agrometeorol, 2010, 31: 310-314 [本文引用: 1]
[4]
Gana JA, SuttonF, Kenefick D G. cDNA structure and expression patterns of a low-temperature-specific wheat genetacr7 . Plant Mol Biol, 1997, 34: 643-650 [本文引用: 1]
[5]
MinamiA, NagaoM, IkegamiK, KoshibaT, ArakawaK, FujikawaS, TakezawaD.Cold acclimation in bryophytes: low- temperature- induced freezing tolerance inPhyscomitrella patens is associated with increases in expression levels of stress-related genes but not with increase in level of endogenous abscisic acid . Planta, 2005, 220: 414-423 [本文引用: 1]
Liu YY, Li JZ, ChenL, Cui DQ.Effect of low temperature stress on peroxidation product of membrane lipids and activity of related enzymes in wheat seeding leaves .J Triticeae Crops, 2006, 26(4): 70-73 (in Chinese with English abstract) [本文引用: 1]
Jiang LN, Zhang DJ, SongF, LiuP, Fan TT, Yu HB, Li CX.Evaluation of cold resistance of different wheat varieties based on physiological responses of leaves to low temperature at the jointing stage .Acta Ecol Sin, 2014, 34: 4251-4261 (in Chinese with English abstract) [本文引用: 5]
Zhang ZW, Wang FH, Li SD, FengB, Si JS, ZhangB.Analysis on physiological reaction to low temperature at booting stage and their cold resistance of different types of wheat varieties .J Triticeae Crops, 2012, 32: 900-906 (in Chinese with English abstract) [本文引用: 2]
ZhangJ, LuM, Sun SG, Du WL, LiuY, WuJ, Chen XH.Changes of physiological and biochemical parameters and grain yield at joining stage of wheat under low temperature stress .Acta Agric Boreali-Occident Sin, 2014, 23(02): 73-79 (in Chinese with English abstract) [本文引用: 2]
Sun JY, Zhao YT, Liang BW, LiuF, YouY.Changes of hydroxyproline-rich glyco-protein wheat under cold stress and its relationship to cold resistance . Plant Genet Resour, 2004, 5: 6-11 (in Chinese with English abstract) [本文引用: 2]
[11]
Equiza MA.Morphological, anatomical and physiological responses related to differential shoot vs. root growth inhibition at low temperature in spring and winter wheat .Ann Bot, 2001, 87: 67-76 [本文引用: 2]
Liu YY, Li JZ, ChenL, Cui DQ.Effect of low temperature stress on peroxidation product of membrane lipids and activity of related enzymes in wheat seed-ling leaves . J Triticeae Crops, 2006, 26(4): 70-73 (in Chinese with English abstract) [本文引用: 1]
[13]
BohnM, LuthjeS.Plasma membrane lipid alterations induced by cold acclimation and abscisic acid treatment of winter wheat seedlings differing in frost resistance .J Plant Physiol, 2007, 164: 146-156 [本文引用: 1]
LiuW, Sun DL, WangH, Jian LC, Zhao KF.Comparison of Ca2+-ATPase activity in seedling plasmolemma of cold resistant winter wheat vs. cold-sensitive spring wheat under 2°C low temperature .Acta Agron Sin, 2002, 28: 227-229 (in Chinese with English abstract) [本文引用: 1]
Jiang G M. Plant Ecophysiology.Beijing: Higher Education Press, 2004. pp 46-53 (in Chinese) [本文引用: 1]
[16]
GentyB, Briantais JM, Baker NR.The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence .Biochim Biophys Acta, 1989, 99: 87-92 [本文引用: 1]
[17]
SchreiberU, BilgerW, NeubauerC.Chlorophyll fluorescence as a non-destructive indicator for rapid assessment of in vivo photo- synthesis .Ecol Studies, 1994, 100: 49-70 [本文引用: 1]
[18]
Krause GH, WeisE.Chlorophyll fluorescence and photosynthesis: the basis . Annu Rev Plant Physiol Plant Mol Biol, 1991, 42: 313-349 [本文引用: 1]
[19]
Van KootenO, Snel JF.The use of chlorophyll fluorescence nomenclature in plant stress physiology .Photosynth Res, 1990, 25: 147-150 [本文引用: 1]
Wang XN, Fu LS, Li ZF, YangF, Sun YL, LiuC, Wang JW, Chen YX.Comparison of chlorophyll fluorescence parameters between two winter wheat cultivars during cold acclimation and frozen period .J Triticeae Crops, 2009, 29: 83-88 (in Chinese with English abstract) [本文引用: 3]
GaoY, Tang JW, Yin GH, Han YL, HuangF, Wang LN.Effect of different periods and frequency of late spring coldness on winter wheat yield related traits .J Triticeae Crops, 2015, 35: 687-692 (in Chinese with English abstract) [本文引用: 1]
Wang SG, Wang ZL, WangP, Wang HW, HuangW, Wu YG, Yin YP.Freeze resistance analysis of different wheat cultivars based on relationships between physiological indices and grain yield . Chin J Appl Ecol, 2011, 22: 1477-1484 (in Chinese with English abstract) [本文引用: 2]
[23]
FracheboudY, HaldimannP, LeipnerJ, StampP.Chlorophyll fluorescence as a selection tool for cold tolerance of photosynthesis in maize .J Exp Bot, 1999, 50: 1533-1540 [本文引用: 2]
[24]
Xu CC, Zhang JH.Effect of drought on chlorophyll fluorescence and xanthophylls cycle components in winter wheat leaves with different ages . Acta Phytophysiol Sin, 1999, 25: 29-37 [本文引用: 2]
[25]
Kratsh HA, Wise RR.The ultrastructure of chilling stress .Plant Cell Environ, 2000, 23: 337-350 [本文引用: 1]
Chen SS, Li CY, YangJ, XuW, Zhu XK, Guo WS, Feng CN.Effect of low temperature at jointing stage on photosynthetic characteristics and yield in wheat cultivar Yangmai 16 . J Yangzhou Univ (Agric Life Sci Edn), 2014, 35(3): 59-64 (in Chinese with English abstract) [本文引用: 1]

 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}