 ,1, 徐文正1,2, 杨锍琰1, 刘玉1, 周治国1, 王友华,1,*
,1, 徐文正1,2, 杨锍琰1, 刘玉1, 周治国1, 王友华,1,*Different response of cotton leaves to heat stress is closely related to the night starch degradation
ZHAO Wen-Qing,1, XU Wen-Zheng1,2, YANG Liu-Yan1, LIU Yu1, ZHOU Zhi-Guo1, WANG You-Hua,1,*通讯作者: * 王友华, E-mail:w_youhua@njau.edu.cn
收稿日期:2020-09-27接受日期:2021-01-21网络出版日期:2021-03-01
| 基金资助: |
Corresponding authors: * E-mail:w_youhua@njau.edu.cn
Received:2020-09-27Accepted:2021-01-21Online:2021-03-01
| Fund supported: |
作者简介 About authors
E-mail:zhaowenqing@njau.edu.cn

摘要
叶片对光合产物的转运输出是其源能力的重要体现, 高温胁迫导致的源能力不足是棉花减产降质的重要原因之一。为探究棉花对位叶光合产物输出能力对短期高温胁迫的响应及其在不同热敏感性品种间的差异, 本研究以耐热型棉花品种PHY370WR和热敏感型棉花品种苏棉15为材料, 于2015—2016年在南京农业大学牌楼试验站进行盆栽试验, 在棉花花铃期设置持续5 d的温度处理[对照日均温为26℃ (CK, 昼夜温度循环为30/22℃), 高温处理日均温为34℃ (HT, 昼夜温度循环为38/30℃)], 在高温胁迫5 d和胁迫解除后5 d对叶片光合产物输出能力和光合产物含量进行研究。结果表明, 高温胁迫5 d显著降低棉花铃重, 提高对位叶比叶重, 耐热型棉花品种PHY370WR铃重降低幅度及比叶重升高幅度均较热敏感型棉花品种苏棉15低。13C标记光合产物的结果表明, 高温5 d显著降低棉花叶片光合产物的输出效率, 与CK相比, 苏棉15的降幅为22.1%, 下降斜率为-2.48, 显著高于PHY370WR的降幅15.7%和下降斜率-1.82; 恢复5 d后棉花叶片光合产物输出效率降低幅度变小, 但品种间差异由高温5 d时的6.4%增加为胁迫解除后5 d的10.2%, 且苏棉15的恢复斜率仅为0.44, 远小于PHY370WR的0.89。此外, 与高温5 d时相比, 恢复5 d后蔗糖含量日变化幅度的品种间差异减小, 而淀粉含量日变化幅度品种间差异表现为增大, 与碳水化合物输出效率的趋势一致, 相关性分析亦表明, 高温胁迫5 d及恢复5 d时淀粉含量日变化幅度与碳水化合物输出效率相关性更显著。通过大田试验和盆栽试验确定淀粉最大、最小含量出现时间, 在此基础上进一步分析发现, 在高温胁迫5 d及胁迫恢复后5 d, 淀粉日最小(夜间)含量升高幅度品种间差异较日最大(白天)含量降低幅度更为显著, 且恢复后5 d时品种间差异较高温5 d时显著增大。综上, 热敏感性差异的棉花叶片碳水化合物输出效率对高温胁迫响应的差异不仅表现在胁迫发生时, 更表现在胁迫解除后; 耐热型棉花品种PHY370WR的叶片在高温胁迫发生时及胁迫恢复后表现出更强的抵御高温逆境能力及更好的恢复能力, 与其夜间淀粉降解能力即叶片最小(夜间)淀粉含量增加幅度较小密切相关。
关键词:
Abstract
The export rate of photosynthetic products from leaves is an important manifestation of their source capacity. Heat induced source capacity shortage is one of the major reasons for cotton yield reduction and quality deterioration. To explore the difference and mechanism of leaf carbohydrate export in response to short-term heat stress between cotton cultivars with different heat sensitivity, pot experiments were carried out using two cotton cultivars PHY370WR (heat tolerance) and Sumian 15 (heat sensitivity) as experimental materials in 2015 and 2016. Two temperature treatment (CK, average temperature 26℃; HT, average temperature 34℃) was conducted at flowering and boll forming stages lasting for five days. Results showed that cotton boll weight was significantly decreased and specific leaf weight was increased by HT. The reduction of boll weight and increase of specific leaf weight of PHY370WR were lower than that of Sumian 15. The results of 13C labeled photosynthetic products showed that the carbohydrate export efficiency (CEE) of cotton leaves was significantly reduced by HT. Compared with CK, the decrease rate of CEE in Sumian 15 was 22.1% and the decline slope was -2.48, significantly higher than that of PHY370WR by 15.7% and -1.82, respectively. The decrease after five days of recovery in CEE diminished, but the differences at five days of HT between varieties were increased from 6.4% to 10.2% compared with at five days after HT released. The recovery slope of CEE in Sumian 15 was only 0.44, far less than 0.89 of PHY370WR. In addition, compared with HT, the difference of daily variation amplitude of sucrose content was decreased and the difference of daily variation amplitude of starch content was increased between cultivars after five days recovery. The latter was consistent with the trend of CEE in response to HT. Correlation analysis revealed that starch content of daily variation amplitude was more significantly correlated with CEE than sucrose. Further analysis showed that at both five days of HT and five days after HT released, the difference of increase in the minimum (night) starch content between cultivars was more significant than that of decrease in the maximum (day) starch content. The differences between cultivars at five days after HT released were significantly larger than that at five days of HT. In conclusion, there were the differences of cotton leaf CEE with different heat sensitivities not only during high temperature stress but also after the relief of stress. The heat tolerant cultivar PHY370WR indicated stronger resistance to HT and better recovery ability after HT released, which was closely related to a better starch degradation ability with a less increase in the minimum starch content in leaves at night.
Keywords:
PDF (655KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
赵文青, 徐文正, 杨锍琰, 刘玉, 周治国, 王友华. 棉花叶片响应高温的差异与夜间淀粉降解密切相关. 作物学报[J], 2021, 47(9): 1680-1689 DOI:10.3724/SP.J.1006.2021.04220
ZHAO Wen-Qing, XU Wen-Zheng, YANG Liu-Yan, LIU Yu, ZHOU Zhi-Guo, WANG You-Hua.
温度对作物生产至关重要, 高温胁迫会显著降低棉花产量。随温室效应的加剧, 夏季极端高温天气日益频发, 成为影响棉花生产的重要自然灾害。高温胁迫导致的源能力不足是棉花减产降质的重要原因之一[1,2,3,4,5,6]。作为棉铃发育最主要的源器官, 棉铃对位叶源能力对高温胁迫的响应对棉花产量形成至关重要[7,8]。叶片源能力主要体现在碳水化合物合成及其向库器官(棉铃)的转运, 因此, 棉铃对位叶的源能力除体现在光合能力外, 还体现在对过剩光合产物的暂时储存能力和有效光合不足时对储存物质的再调动能力。
叶片中碳水化合物的含量变化不仅能反映叶片光合作用的强弱, 还能反映叶片中碳水化合物的输出转运水平。蔗糖和淀粉是棉花叶片光合作用的主要产物, 两者的含量除随生育进程的推进产生变化外, 还存在显著的昼夜变化规律。其中, 蔗糖是叶片中碳水化合物输出的主要形式, 白天叶片的蔗糖含量水平与光合作用强弱密切相关, 直接决定着碳水化合物夜间的输出速率[9,10,11], 而夜间叶片蔗糖含量除受白天蔗糖含量的影响外, 还与淀粉降解产生蔗糖的速率有关; 淀粉则起着暂时储存的作用, 白天叶片中大量不被转移利用的光合产物会在叶绿体中转化成淀粉暂时储存, 夜间再转化为蔗糖输出到相应的库器官中[12,13], 因此淀粉昼夜间含量水平的变化应能较好的反映叶片碳水化合物的输出转运能力。
高温胁迫对叶片中碳水化合物含量的影响研究较多, 研究认为高温胁迫导致叶片中蔗糖含量显著升高, 淀粉含量显著降低[14]; 夜间高温显著降低叶片蔗糖含量[15]; 高温胁迫显著降低叶片蔗糖、淀粉含量[16]等; 亦有研究表明, 棉花叶片中淀粉含量在增温条件下会显著升高[17]。可见, 不同研究的结果不尽相同, 这可能与高温胁迫发生时间、胁迫程度与持续时间密切相关。蔗糖转化率常作为衡量叶片碳水化合物输出效率的重要指标之一, 以往的研究常关注基于生育进程变化的蔗糖含量、淀粉含量及两者转化率对高温的响应, 对蔗糖、淀粉昼夜变化如何响应高温的研究较少。棉花叶片光合能力在高温胁迫解除后的研究发现, 低于40℃的胁迫对叶片光合作用的抑制是可恢复的[18,19], 但叶片物质转运能力在胁迫解除后是否具有恢复效应尚不明确。此外, 不同品种棉花叶片的光合产物输出能力存在显著差异, 较强的光合产物输出能力有利于棉花铃重的形成[20], 但不同品种间叶片耐热性差异及其机制的研究多集中于光合能力和抗氧化代谢方面[2,15,21-22], 关于不同棉花品种间叶片光合产物转运能力响应高温胁迫差异的研究尚显不足[23,24]。
综上, 我们假设短期高温胁迫会显著降低叶片碳水化合物的输出效率, 且在品种间存在显著差异。通过对高温胁迫发生时和胁迫解除后叶片碳水化合物输出效率及与物质含量的分析, 可明确品种间差异形成的机理及关键生理过程。稳定性同位素13C标记技术在国内外已广泛应用于植物生理生态学等多个领域[25], 也是国际公认的研究碳水化合物转运分配最有效的方法[26,27], 相关标记方法和检测手段十分成熟。因此, 本研究拟通过利用13C标记技术, 结合叶片中蔗糖和淀粉含量的昼夜变化, 从光合产物暂时储存和转运能力入手, 系统研究胁迫下和胁迫解除后不同热敏感性棉花叶片源能力响应花铃期短期高温胁迫的差异, 以期阐明不同基因型棉花品种对短期高温胁迫敏感性差异产生的生理机制。
1 材料与方法
1.1 试验设计
盆栽试验于2015—2016年在江苏南京(118º50′E, 32º02′N)南京农业大学牌楼试验站进行。供试土壤为黄棕壤土, 土壤晒干除杂过筛后装于聚乙烯塑料桶中(直径37 cm、高32 cm, 底部有排水孔), 每塑料桶装过筛的风干土20 kg。2015年和2016年耕层土壤分别含有机质19.3 g kg-1和20.5 g kg-1、全氮1.5 g kg-1和1.3 g kg-1、速效氮95.5 mg kg-1和102.5 mg kg-1、速效磷17.9 mg kg-1和18.8 mg kg-1、速效钾102.3 mg kg-1和110.5 mg kg-1。选择热敏感型棉花品种苏棉15和耐热型棉花品种PHY370WR为材料[28], 棉花种子分别于2015年4月5日和2016年4月10日进行营养钵育苗, 三叶期(2015年5月5日和2016年5月8日)选择均匀一致的壮苗移栽, 移栽前浇水压实桶中土壤, 每盆定植1株, 每个品种300株。在移入人工气候室前, 棉花分品种排列放于半开放式防雨棚中, 每品种6列, 每列50株。在棉株第6~8果枝第1、2果节开花时(7月上旬, 棉花花铃期), 挂牌标记当日所开白花及其对位叶, 同时选取生长状况相近的棉株320株(苏棉15、PHY370WR各160株), 分别移入人工气候室进行5 d的对照处理[26℃ (30℃, 22℃), 用CK表示]和高温处理[34℃ (38℃, 30℃), 用HT表示], 每品种每处理各80株棉株。温度控制模拟自然温度的昼夜变化, 对照26℃ (30℃, 22℃)设置温度控制循环为: 2:00 (24℃)—4:00 (23℃)—6:00 (22℃)—8:00 (24℃)— 10:00 (27℃)—12:00 (29℃)—14:00 (30℃)—16:00 (29℃)—18:00 (28℃)—20:00 (27℃)—22:00 (27.5℃)— 24:00 (26℃); 高温34℃ (38℃, 30℃)设置温度控制循环为: 2:00 (32℃)—4:00 (31℃)—6:00 (30℃)— 8:00 (32℃)—10:00 (35℃)—12:00 (37℃)—14:00 (38℃)—16:00 (37℃)—18:00 (36℃)—20:00 (35℃)— 22:00 (35.5℃)—24:00 (34℃)]。连续5 d温度处理条件下, 对照日均温为25.8℃, 高温处理的日均温为34.2℃, 说明达到了试验预期温度控制目标, 具体见本实验室发表文章(文献[28]的图1)[28]。高温处理5 d后移至原半开放式防雨棚中恢复5 d。肥水和病虫害管理按棉花高产栽培进行。
图1
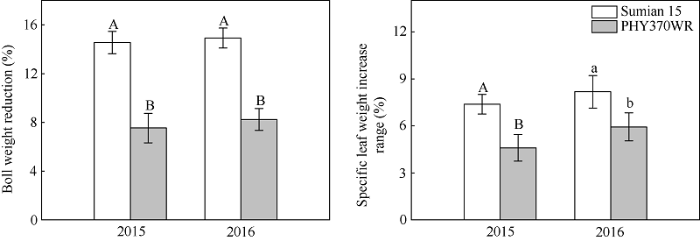 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1高温胁迫对棉花铃重及其比叶重的影响
在收获期调查了20个非取样棉花植物, 对标记部位棉铃铃重和对位叶叶片比叶重进行统计分析。图中大小写字母分别表示品种间达到极显著(P < 0.01)或显著( P < 0.05)差异。
Fig. 1Effects of high temperature stress on specific leaf weight and boll weight with different heat tolerance in cotton
The boll weight and the specific leaf weight data are derived from the investigation of 20 non-sampling cotton plants at harvest stage. Different uppercase and lowercase letters indicate significant differences at P < 0.01 and P < 0.05 between cultivars according to the shortest significant ranges (SSR) test, respectively.
为对比盆栽试验与大田中棉铃对位叶蔗糖和淀粉含量昼夜变化规律, 大田试验于2015年在江苏南京(118º50′E, 32º02′N)南京农业大学牌楼试验站进行。供试土壤为黄棕壤土。与盆栽试验一致, 在棉株第6~8果枝第1、2果节开花时(7月上旬, 棉花花铃期), 挂牌标记当日所开白花及其对位叶, 并与盆栽试验在同一天同时取样, 对比两试验条件下叶片中蔗糖和淀粉含量的日变化情况; 大田栽培条件下每1 h取样1次, 每次取5片叶; 盆栽试验条件下, 每3 h取样1次, 每次取5片叶。
1.2 测定内容与方法
1.2.1 比叶重、铃重 棉花吐絮收获时, 取挂牌标记的非取样棉花棉铃及其对位叶, 测定铃重及对位叶比叶重。对位叶用叶面积仪测定后放于烘箱, 在105℃条件下杀青30 min, 80℃烘干至恒重并称重, 计算比叶重(specific leaf weight, g m-2)。计算公式: 比叶重 = 叶片干重/叶面积。1.2.2 同位素标记及其测定 同位素标记分别于高温处理前、处理结束时及恢复5 d后进行。选取中部果枝(6~8果枝)生长状况相近的相同花后天数的挂牌标记棉铃对位叶, 于上午9时左右, 用透明密封塑料袋包裹叶片(叶片与袋子间留有足够的空隙), 用注射器注入2 mL 13CO2 (由上海化工研究院提供, 纯度为99.99%), 2 h之后移除透明塑料袋, 同时对一部分叶片进行取样, 剩余叶片则于24 h之后取样, 烘干磨碎过100目筛后使用元素分析仪-质谱联用测定其同位素丰度。
参考谷淑波等[27]测定同位素13C丰度, 称取2.0 mg左右(叶片)的样品包入锡纸中, 称重后记录样品重量, 放入元素分析仪的自动进样器后按照放样顺序在控制软件的表格中依次输入重量数据, 每隔10个试验样品要插入3个标准物质。由叶片中测得的13C同位素相对丰度值计算碳水化合物输出效率, 计算公式: 碳水化合物输出效率(%) = (100% × (饲喂2 h后叶片中的δ13C-饲喂24 h后叶片中的δ13C))/饲喂2 h后叶片中的δ13C。
1.2.3 碳水化合物含量测定及日变化幅度Δ 采用间苯二酚比色法[29]测定对位叶蔗糖含量, 采用蒽酮比色法[8]测定淀粉含量。淀粉/蔗糖日变化幅度Δ = (最大含量(淀粉/蔗糖) - 最小含量(淀粉/蔗糖))/最大含量(淀粉/蔗糖)×100%。
1.3 数据统计与分析方法
使用Microsoft Excel 2013、SPSS 17.0处理和分析数据, 用Origin 2015作图。2 结果与分析
2.1 棉花铃重和对位叶比叶重响应短期高温胁迫的品种间差异
与CK相比, 高温显著降低棉花铃重, 提高棉铃对位叶比叶重, 两品种表现一致(图1)。其中, 苏棉15的铃重降低幅度为14.8% (2年平均, 下同), 显著大于PHY370WR的7.9%; 苏棉15对位叶比叶重的增加幅度为7.8%, PHY370WR为5.3%, 可以看出, 耐热型棉花品种PHY370WR在遭遇高温胁迫后铃重降低幅度和比叶重增加幅度均显著小于高温敏感型棉花品种苏棉15。2.2 棉花叶片碳水化合物输出效率(能力)响应短期高温胁迫的品种间差异
2年同位素标记结果均显示, 高温胁迫显著降低棉铃对位叶光合碳水化合物的输出效率(图2)。与CK相比, 高温处理5 d苏棉15的碳水化合物输出效率降低幅度为22.1%, 下降斜率为-2.48, 显著高于PHY370WR的降低幅度15.7%和下降斜率-1.82。高温胁迫解除5 d后, 碳水化合物输出效率有所恢复, 此时苏棉15的降幅为18.2%, 恢复斜率为0.44, PHY370WR的降幅为8.0%, 恢复斜率为0.89。此外, 碳水化合物输出效率的降低幅度品种间差异由高温5 d时的6.4%上升到高温胁迫解除后5 d时的10.2%, 说明高温胁迫下棉铃对位叶光合产物转运能力(输出效率)的品种间差异不仅体现在高温胁迫发生时, 更体现在高温胁迫解除后。图2
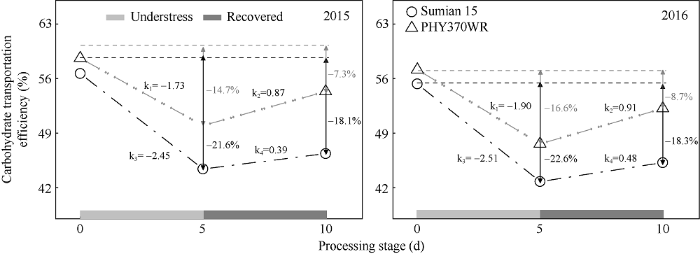 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2高温胁迫下及胁迫解除后5 d叶片碳水化合物输出能力的变化及其在品种间的差异
k1和k3分别表示高温胁迫下PHY370WR与苏棉15碳水化合物输出效率的下降斜率; k2和k4分别表示高温胁迫解除后PHY370WR与苏棉15碳水化合物输出效率的恢复斜率。
Fig. 2Differences of carbohydrate transportation efficiency in leaves of different heat-tolerant cultivars under and after high temperature
k1 and k3 represent the decline slopes of PHY370WR and Sumian 15 under heat stress, respectively; k2 and k4 represent the recovery slopes of PHY370WR and Sumian 15 after heat stress released, respectively.
2.3 棉铃对位叶淀粉、蔗糖含量日变化幅度响应高温胁迫的品种间差异及其与碳水化合物输出效率的相关性
2.3.1 棉花对位叶蔗糖、淀粉含量的昼夜变化规律高温胁迫下, 棉花叶片蔗糖、淀粉含量表现出显著的昼夜变化规律(图3-A)。尽管盆栽条件下棉花叶片蔗糖和淀粉含量的日变化幅度(35.3%, 51.1%)显著低于大田栽培条件下(40.8%, 62.5%), 但2种栽培条件下叶片中蔗糖、淀粉含量的日变化规律无显著差异(图3-B), 均表现为白天随着光合作用的增强, 蔗糖含量上升, 于12时30分左右达到最高, 之后持续降低, 22时30分左右降至最低; 淀粉含量在白天积累, 于19时左右达到最高, 之后开始降低, 至次日早上6时30分左右降至最低。因此, 本研究中盆栽试验的结果具有一定的普适性。
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3棉花叶片中蔗糖、淀粉含量的日变化规律
图A为大田栽培条件下叶片中蔗糖和淀粉含量的日变化, 每1 h取样1次。图B为盆栽试验条件下的叶片中蔗糖和淀粉含量的日变化情况, 每3 h取样1次。
Fig. 3Diurnal variation of sucrose content and starch content in cotton leaves
The picture A shows the diurnal variation of sucrose and starch content in the leaves under field cultivation conditions which is sampled every hour. The picture B shows the performance under pot experiment conditions which is sampled every three hours.
2.3.2 棉花对位叶蔗糖、淀粉含量日变化幅度响应高温胁迫的品种间差异 高温显著降低棉花对位叶淀粉含量日变化幅度(图4)。与CK相比, 高温5 d时苏棉15 和PHY370WR对位叶淀粉含量的日变化幅度分别降低了18.4%和11.9% (品种间差异6.5%); 高温恢复5 d时, 苏棉15和PHY370WR对位叶淀粉含量的日变化幅度分别降低了22.3%和13.1% (品种间差异9.2%), 说明, 与高温胁迫5 d相比, 高温胁迫恢复5 d时的淀粉含量日变化幅度品种间差异增大, 这与叶片碳水化合物输出效率响应高温的结果相一致。
图4
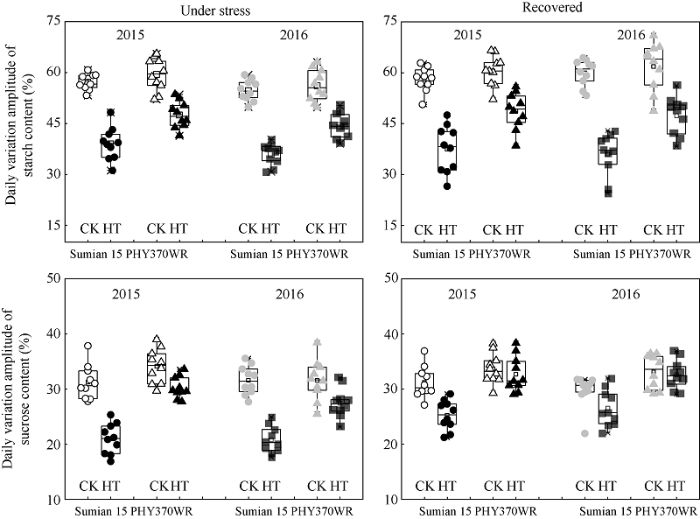 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4高温胁迫对棉花叶片中淀粉、蔗糖含量日变化幅度的影响
CK: 常温对照, 平均日均温26℃; HT: 高温胁迫, 平均日均温34℃。
Fig. 4Diurnal variation of sucrose content and starch content in cotton leaves
CK: normal temperature control and average temperature 26℃; HT: heat stress, and average temperature 34℃.
高温5 d显著降低棉花对位叶蔗糖含量日变化幅度(图4)。与CK相比, 高温5 d时苏棉15 和PHY370WR对位叶蔗糖含量的日变化幅度分别降低了10.5%和4.0% (品种间差异6.5%); 高温恢复5 d时, 苏棉15 和PHY370WR对位叶蔗糖含量的日变化幅度分别降低了4.6%和0.7% (品种间差异3.8%)。因此, 与高温胁迫5 d相比, 高温胁迫恢复5 d时的蔗糖含量日变化幅度品种间差异减小。
2.3.3 高温胁迫下棉铃对位叶光合产物含量日变化与碳水化合物输出能力的相关性 高温胁迫5 d和恢复5 d时, 棉铃对位叶中蔗糖和淀粉含量的日变化幅度与叶片碳水化合物输出能力存在显著正相关关系(图5), 且淀粉含量日变化幅度与叶片碳水化合物输出能力的相关性更高。此外, 与高温胁迫5 d时相比, 高温胁迫解除5 d时两品种淀粉含量日变化幅度与叶片碳水化合物输出能力的相关性均增强, 结合2.3.2的结果, 说明淀粉日变化幅度更能反映叶片碳水化合物输出能力。
图5
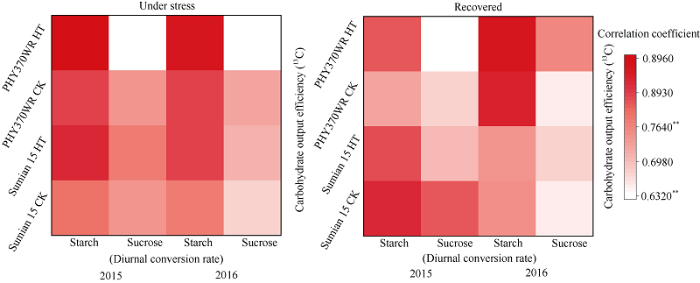 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5高温胁迫5 d及恢复5 d时棉铃对位叶蔗糖、淀粉含量日变化幅度与碳水化合物输出效率的相关性
用同位素13C标记结果计算所得的碳水化合物输出效率与叶片中淀粉含量和蔗糖含量的日最大变化幅度进行相关性分析, 对应色块的颜色越深表明相关性越强。**表示1%显著水平。
Fig. 5Correlation between diurnal variation of sucrose and starch content and carbohydrate transportation efficiency in cotton leaves after five days of high temperature stress and five days of recovery
The darker the color of the block, the stronger the correlation between carbohydrate transportation efficiency and the diurnal conversion rates of starch content and sucrose content in subtending leaves. ** Significant at the 1% probability level.
2.3.4 棉铃对位叶淀粉含量日最大值、最小值响应高温胁迫的品种间差异 高温胁迫降低了棉铃对位叶日最大淀粉含量, 提高了日最小淀粉含量, 因而减小了淀粉含量日变化幅度(图6)。高温胁迫5 d时, 与CK相比, 苏棉15和PHY370WR日最大淀粉含量降低幅度分别为10.3%和8.0% (品种间差异为2.3%), 日最小淀粉含量增加幅度分别为26.9%和20.5% (品种间差异为6.4%); 胁迫解除5 d后日最大淀粉含量降低幅度分别为9.5%和8.1% (品种间差异为1.4%), 日最小淀粉含量增加幅度分别为27.6%和16.1% (品种间差异为11.5%)。因此, 与高温胁迫5 d相比, 高温胁迫恢复5 d时的日最小淀粉含量变化幅度品种间差异增大。由上可知, 高温胁迫对日最小淀粉含量的影响较对日最大淀粉含量的影响更显著, 且其变化趋势与高温胁迫与胁迫解除后淀粉含量日变化的趋势一致, 因此, 高温胁迫下两品种对位叶中淀粉含量日变化幅度的差异主要由夜间最小淀粉含量的差异造成。
图6
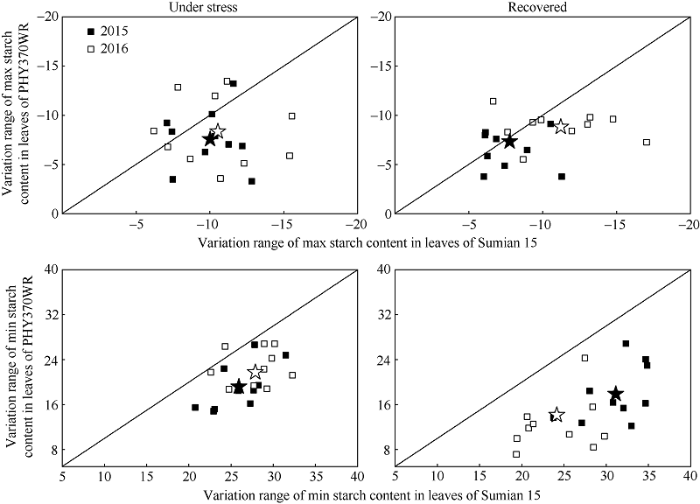 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6不同热敏感性品种棉铃对位叶淀粉昼夜极值响应高温胁迫的差异
纵轴为PHY370WR叶片中日最大/最小淀粉含量的变化幅度, 横轴为苏棉15叶片中日最大/最小淀粉含量的变化幅度, 图中点的分布距离斜线越远, 表示品种间差异越大。叶片中日最大/最小淀粉含量变化幅度 = (高温胁迫处理叶片日最大/最小淀粉含量 - 对照叶片日最大/最小淀粉含量)/(对照叶片日最大/最小淀粉含量)。
Fig. 6Variation of high temperature stress to diurnal variation of starch content in leaves of cotton boll cultivars
The vertical axis is the variation of maximum/minimum starch content in the leaves of PHY370WR, while the horizontal axis is the variation of maximum/minimum starch content in the leaves of Sumian 15. The farther the distribution of points in the figure is from the oblique line, the greater the difference between cultivars. Variation of maximum/minimum starch content = (maximum/minimum starch content HT - maximum/minimum starch content CK)/(maximum/minimum starch content CK).
3 讨论
花铃期5 d的短期高温处理后, 棉花铃重显著下降, 比叶重却显著增加。棉铃与对位叶是一个最基本的源库单位 [30], 高温胁迫导致铃重降低的同时叶片比叶重增加, 说明高温胁迫导致产量降低的原因不仅是光合产物的合成受到了抑制, 光合产物在源库器官间的分配也发生了改变。尽管开花时棉铃对位叶叶片大小基本定型, 但在后续生长发育中, 叶片厚度和重量还会有一定量的变化, 本研究中短期高温胁迫条件下比叶重有所增加, 与光合产物从叶片向棉铃的输出受阻有一定的关系。比较2个不同热敏感性的品种发现, 铃重大幅减少的苏棉15, 其叶片比叶重的增加也更为显著。光合产物在叶片与棉铃间的分配与叶片光合产物的输出转运密不可分, 2个品种叶片光合产物在叶片与棉铃间分配比例变化的差异, 表明其叶片光合产物输出效率(能力)对短期高温胁迫的响应也存在着显著差异。高温胁迫导致的源能力不足是棉花减产降质的重要原因之一[1,2,3,4,5,6], 叶片光合作用和光合产物的转运输出是既相互紧密联系又在某种程度上独立调控的2个生理代谢过程, 叶片中光合产物的转运和储存能力也是叶片源能力的重要方面。以往关于叶片光合产物输出能力的研究, 主要通过分析源、库器官间光合产物分配比例的相对变化来实现, 而碳同位素标记是研究叶片中碳水化合物在器官间转运分配的常用方法[31], 可实现对单张叶片内光合产物输出比例的量化分析。本研究采用13CO2对叶片中光合产物进行标记, 用13C的相对丰度值计算得到棉花叶片内光合产物24 h内的转运效率, 约为60%, 与前人用放射性同位素(14CO2)的标记结果一致[32]。与CK相比, 高温胁迫显著抑制了叶片的碳水化合物输出能力, 高温胁迫下24 h内耐高温型棉花品种PHY370WR叶片碳水化合物输出效率下降幅度2年平均值为15.7%, 显著小于苏棉15的22.1%; 胁迫解除后, PHY370WR和苏棉15叶片碳水化合物输出效率下降幅度2年平均值分别为8.0%和18.2%。可见, 不同热敏感性品种棉铃对位叶碳水化合物输出能力响应高温胁迫的差异在胁迫解除后更加显著。因此, 高温胁迫对棉铃对位叶光合产物转运能力的影响在品种间存在显著差异, 差异不但体现在高温胁迫发生时, 更体现在高温胁迫解除后。本研究中, 耐高温型棉花品种PHY370WR在高温胁迫发生时及胁迫恢复后碳水化合物输出效率表现出更强的抵御高温逆境能力, 即更高的保持稳定的能力, 因而高温下PHY370WR叶片比叶重的增加幅度、铃重的降低幅度均相对较小。
蔗糖和淀粉是棉花叶片光合作用的主要产物, 两者含量水平高低和昼夜变化幅度与叶片碳水化合物输出能力密切相关。本研究根据前人研究结果[33], 结合本试验的动态测定结果, 确定了棉花叶片中淀粉和蔗糖含量的昼夜变化规律, 计算出两者昼夜间变化幅度, 并研究其与碳水化合物输出效率之间的关系。结果表明, 高温胁迫显著降低棉花叶片淀粉、蔗糖含量昼夜变化幅度, 两品种间降低幅度差异均为6.5%; 胁迫恢复后5 d, 淀粉含量昼夜变化幅度仍然显著降低且两品种间差异增加为9.2%, 而蔗糖含量昼夜变化幅度降低程度减小, 两品种间差异减小为3.8%。相关性结果亦表明, 高温胁迫5 d及恢复后5 d蔗糖、淀粉昼夜变化幅度均与碳水化合物输出能力(同位素标记结果)存在显著的正相关关系, 其中, 淀粉含量昼夜变化幅度与碳水化合物输出效率的相关性更强。蔗糖作为碳水化合物输出的主要形式, 其白天的含量水平参与夜间输出速率的建成, 而淀粉作为叶片中光合产物的暂时储存形式, 白天合成积累, 夜间再转化为蔗糖输出到库器官中, 因此叶片中蔗糖含量昼夜变化幅度与碳水化合物输出速率紧密相关[34], 淀粉含量昼夜变化幅度应能反映出叶片中光合产物输出能力的大小[12, 35]。本研究中, 在高温条件下, 白天叶片蔗糖含量显著降低, 其输出效率受到显著抑制, 此时, 能更多地储存淀粉并在夜间更多的转化为蔗糖就显得极为重要。
然而, 高温胁迫下, 不同耐热性棉花品种间差异到底是因为能更多的储存淀粉还是能较好的保证淀粉的转化?尚未可知。本研究进一步分析了淀粉含量的昼夜极值对高温胁迫的响应及其在品种间的差异, 发现高温胁迫下, 与CK相比, 最小(夜间)淀粉含量上升较最大(白天)淀粉含量下降幅度更大; 在高温胁迫解除5 d后, 叶片最小(夜间)淀粉含量增加幅度的品种间差异与高温5 d相比显著增加, 而最大(白天)含量降低幅度与胁迫下无显著变化。因此, 淀粉最小含量增加幅度的品种间差异是淀粉昼夜变化幅度降低的主要原因, 即淀粉夜间降解能力的减弱是其光合产物输出能力降低的重要原因。本研究中, 热敏感型棉花品种苏棉15叶片最小(夜间)淀粉含量的增加是其叶片淀粉含量昼夜变化幅度降低程度显著小于耐热型棉花品种PHY370WR的主要原因, 也是其表现出在高温下更为敏感特性的原因。综上, 我们认为叶片淀粉夜间降解能力的下降是介导短期高温胁迫后不同热敏感性品种叶片光合产物输出能力差异的重要生理过程。
4 结论
不同热敏感性棉花叶源铃库间的光合产物分配比例对高温胁迫的响应存在显著差异, 表现为耐热型棉花品种铃重降低幅度及比叶重升高幅度均较热敏感型棉花品种低。热敏感性差异的棉花品种叶片碳水化合物输出效率对高温胁迫响应的差异不仅表现在胁迫发生时, 更表现在胁迫解除后。耐热型棉花品种的叶片在高温胁迫发生时及胁迫恢复后表现出更强的抵御高温逆境能力及更好的恢复能力, 与其夜间淀粉降解能力即叶片最小(夜间)淀粉含量增加幅度较小密切相关。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 2]
[本文引用: 2]
DOIURL [本文引用: 3]
DOIURL [本文引用: 2]
DOIURL [本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
DOIURL [本文引用: 1]
DOIURL [本文引用: 2]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
PMID [本文引用: 2]

The influence of growth temperature on the free sugar and sucrose phosphate synthase content and activity of spinach (Spinacia oleracea) leaf tissue was studied. When plants were grown at 25 degrees C for 3 weeks and then transferred to a constant 5 degrees C, sucrose, glucose, and fructose accumulated to high levels during a 14-d period. Predawn sugar levels increased from 14- to 20-fold over the levels present at the outset of the low-temperature treatment. Sucrose was the most abundant free sugar before, during, and after exposure to 5 degrees C. Leaf sucrose phosphate synthase activity was significantly increased by the low-temperature treatment, whereas sucrose synthase and invertases were not. Synthesis of the sucrose phosphate synthase subunit was increased during and after low-temperature exposure and paralleled an increase in the steady-state level of the subunit. The increases in sucrose and its primary biosynthetic enzyme, sucrose phosphate synthase, are discussed in relation to adjustment of metabolism to low nonfreezing temperature and freezing stress tolerance.
[本文引用: 1]
[本文引用: 1]
PMID [本文引用: 1]
This study was undertaken to determine the role of sucrose-metabolizing enzymes in altered carbohydrate partitioning caused by heat stress. Potato (Solanum tuberosum L.) genotypes characterized as susceptible and tolerant to heat stress were grown at 19/17[deg]C, and a subset was transferred to 31/29[deg]C. Data were obtained for plant growth and photosynthesis. Enzyme activity was determined for sucrose-6-phosphate synthase (SPS) in mature leaves and for sucrose synthase, ADP-glucose pyrophosphorylase, and UDP-glucose pyrophosphorylase in developing tubers of plants. High temperatures reduced growth of tubers more than of shoots. Photosynthetic rates were unaffected or increased slightly at the higher temperature. Heat stress increased accumulation of foliar sucrose and decreased starch accumulation in mature leaves but did not affect glucose. SPS activity increased significantly in mature leaves of plants subjected to high temperature. Changes in SPS activity were probably not due to altered enzyme kinetics. The activity of sucrose synthase and ADP-glucose pyrophosphorylase was reduced in tubers, albeit less quickly than leaf SPS activity. There was no interaction of temperature and genotype with regard to the enzymes examined; therefore, observed differences do not account for differences between genotypes in heat susceptibility.
DOIURL [本文引用: 2]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
PMID [本文引用: 1]
This study was conducted to determine if reproductive growth in cotton (Gossypium hirsutum L.) affects concurrent leaf development. Apparent photosynthesis (AP), stomatal conductance (Cs), soluble protein (SP), ribulose bisphosphate carboxylase (RuBisCO), and chlorophyll (Chl) were monitored in four main-stem cotton leaves which emerged at approximately 2 week intervals. The leaf which emerged during vegetative growth (48 days after planting) had higher AP, SP, and RuBisCO levels than that present in any leaves which emerged during fruit development. The last leaf studied (89 days after planting) was still present after boll maturation was completed and exhibited a rejuvenation in AP, SP, RuBisCO, and Chl starting at 30 days after leaf emergence. At 96 days after planting, the P700 Chl a-protein complex (PSI) was virtually absent from the leaves that emerged at 48 and 62 days after planting. The light harvesting Chl a/b complex was still present in these leaves, indicating greater degradation of PSI. The data emphasize the influence of developing fruit on concurrently developing leaves, an effect which was alleviated after boll maturation was completed. The declining AP per unit leaf area and smaller leaf size at the top of the plant results in a reduced photosynthetic potential of successively later emerging leaves. This reduction in leaf AP is consistent with earlier reported seasonal canopy photosynthesis patterns.
DOIURL [本文引用: 1]
DOIPMID [本文引用: 1]
Numerous studies have illustrated the need for antioxidant enzymes in acquired photosynthetic thermotolerance, but information on their possible role in promoting innate thermotolerance is lacking. We investigated the hypothesis that genotypic differences in source leaf photosynthetic thermostability would be dependent upon prestress capacity for antioxidant protection of the photosynthetic apparatus in Gossypium hirsutum. To test this hypothesis, thermosensitive (cv. ST4554) and reportedly thermotolerant (cv. VH260) G. hirsutum plants were exposed to control (30/20 degrees C) or high-day temperature (38/20 degrees C) conditions during flowering and source leaf gas exchange, chlorophyll content and maximum photochemical efficiency (F(v)/F(m)) were measured for each treatment. The relationship between source leaf thermostability and prestress antioxidant capacity was quantified by monitoring the actual quantum yield response of photosystem II (PSII) (Phi(PSII)) to a range of temperatures for both cultivars grown under the control temperature regime and measuring antioxidant enzyme activity for those same leaves. VH260 was more thermotolerant than ST4554 as evidenced by photosynthesis and F(v)/F(m) being significantly lower under high temperature for ST4554 but not VH260. Under identical growth conditions, VH260 had significantly higher optimal and threshold temperatures for Phi(PSII) and glutathione reductase (GR; EC 1.8.1.7) activity than ST4554, and innate threshold temperature was dependent upon endogenous GR and superoxide dismutase (SOD; EC 1.15.1.1) activity. We conclude that maintaining a sufficient antioxidant enzyme pool prior to heat stress is an innate mechanism for coping with rapid leaf temperature increases that commonly occur under field conditions.
[本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOIURL [本文引用: 3]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
PMID [本文引用: 1]
In fully expanded leaves of greenhouse-grown cotton (Gossypium hirsutum L., cv Coker 100) plants, carbon export, starch accumulation rate, and carbon exchange rate exhibited different behavior during the light period. Starch accumulation rates were relatively constant during the light period, whereas carbon export rate was greater in the afternoon than in the morning even though the carbon exchange rate peaked about noon. Sucrose levels increased throughout the light period and dropped sharply with the onset of darkness; hexose levels were relatively constant except for a slight peak in the early morning. Sucrose synthase, usually thought to be a degradative enzyme, was found in unusually high activities in cotton leaf. Both sucrose synthase and sucrose phosphate synthetase activities were found to fluctuate diurnally in cotton leaves but with different rhythms. Diurnal fluctuations in the rate of sucrose export were generally aligned with sucrose phosphate synthase activity during the light period but not with sucrose synthase activity; neither enzyme activity correlated with carbon export during the dark. Cotton leaf sucrose phosphate synthase activity was sufficient to account for the observed carbon export rates; there is no need to invoke sucrose synthase as a synthetic enzyme in mature cotton leaves. During the dark a significant correlation was found between starch degradation rate and leaf carbon export. These results indicate that carbon partitioning in cotton leaf is somewhat independent of the carbon exchange rate and that leaf carbon export rate may be linked to sucrose formation and content during the light period and to starch breakdown in the dark.
[本文引用: 1]
[本文引用: 1]
PMID [本文引用: 1]
The partitioning of carbon in intact, mature cotton (Gossypium hirsutum L.) leaves was examined by steady-state (14)CO(2) labeling. Plants were exposed to dark periods of varying lengths, followed by similar illuminated labeling periods. These treatments produced leaves with a range of starch and soluble sugar contents, carbon exchange, and carbon export rates. Export during the illuminated periods was neither highly correlated with photosynthesis nor was export during the illuminated periods significantly different among the treatments. In contrast, the rate of subsequent nocturnal carbon export from these leaves varied widely and was found to be highly correlated with leaf starch content at the end of the illumination period (r = 0.934) and with nocturnal leaf respiration (r = 0.954). Leaves which had accumulated the highest levels of starch (about 275 micrograms per square centimeter) by the end of the illumination period exhibited nocturnal export rates very similar to those during the daylight hours. Leaves which accumulated starch to only 50 to 75 micrograms per square centimeter virtually ceased nocturnal carbon export. For leaves with starch accumulations of between 50 and 275 micrograms per square centimeter, nocturnal export was directly proportional to leaf starch at the end of the illumination period. After the nocturnal export rate was established, it continued at a constant rate throughout the night even though leaf starch and sucrose contents declined.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}