 ,, 王宝祥, 周振玲, 方磊, 迟铭, 李景芳, 刘金波, Bello Babatunde Kazeem, 徐大勇,*连云港市农业科学院 / 江苏省现代作物生产协同创新中心, 江苏连云港 222006
,, 王宝祥, 周振玲, 方磊, 迟铭, 李景芳, 刘金波, Bello Babatunde Kazeem, 徐大勇,*连云港市农业科学院 / 江苏省现代作物生产协同创新中心, 江苏连云港 222006Screening of germplasm resources and QTL mapping for germinability under submerged condition in rice (Oryza sativa L.)
SUN Zhi-Guang,, WANG Bao-Xiang, ZHOU Zhen-Ling, FANG Lei, CHI Ming, LI Jing-Fang, LIU Jin-Bo, Bello Babatunde Kazeem, XU Da-Yong,*Lianyungang Academy of Agricultural Sciences / Jiangsu Collaborative Innovation Center for Modern Corp Production, Lianyungang 222006, Jiangsu, China通讯作者:
收稿日期:2020-04-27接受日期:2020-07-2网络出版日期:2021-01-12
| 基金资助: |
Received:2020-04-27Accepted:2020-07-2Online:2021-01-12
| Fund supported: |
作者简介 About authors
E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (572KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
孙志广, 王宝祥, 周振玲, 方磊, 迟铭, 李景芳, 刘金波, Bello Babatunde Kazeem, 徐大勇. 水稻萌发耐淹性种质资源筛选及QTL定位[J]. 作物学报, 2021, 47(1): 61-70. doi:10.3724/SP.J.1006.2021.02030
SUN Zhi-Guang, WANG Bao-Xiang, ZHOU Zhen-Ling, FANG Lei, CHI Ming, LI Jing-Fang, LIU Jin-Bo, Bello Babatunde Kazeem, XU Da-Yong.
水稻是世界上最重要的粮食作物之一, 水稻的稳产增产事关全球的粮食安全[1,2,3]。在我国, 传统的移栽种植模式在长时间里为我国粮食增产、农民增收发挥了十分重要的作用[4], 但是随着我国城镇化的有序推进, 农村劳动力大量减少, 插秧期间对劳动力的高需求阻碍着传统移栽模式的进行。与传统移栽模式相比, 直播稻整地用水量少, 连续淹水天数少、对灌溉水利用效率高, 而且便于机械化操作[5,6,7]。在此背景下, 随着适宜新品种的育成和栽培技术的进步, 资源和劳动力节约型的轻简栽培方式—直播稻得到了较大的发展[8,9]。
出苗率低是水稻直播中面临的一个主要问题, 当种子被水淹没时, 由于氧气浓度急剧降低, 种子有氧呼吸受到严重抑制, 导致出苗率显著下降[10]。因此, 在淹水条件下, 选择出苗能力好的耐淹性水稻品种将在直播稻安全生产中起到十分重要的作用。已有研究表明, 在低氧条件下萌发的水稻种子胚芽鞘长度与耐淹成苗率呈极显著的正相关[11,12,13,14]。随着分子标记技术的发展, 前人以胚芽鞘长度为指标, 利用多种遗传群体定位到多个萌发耐淹性QTL, 侯名语等[11]以水淹胁迫下暗发芽5 d后的胚芽鞘长为指标, 评价了359份水稻品种的耐低氧发芽能力, 其中11%粳稻品种芽鞘长度超过3.0 cm, 而芽鞘长度超过3.0 cm籼稻品种仅占2%, 其认为籼粳稻在耐低氧发芽能力方面存在差异。随后其利用Kinmaze/DV85 RIL群体, 在水稻1号、2号、5号和7号染色体上共检测到5个低氧发芽相关QTL位点, 可解释表型变异为10.5%~19.6%。陈孙禄等[15]通过对256份水稻核心种质进行萌发耐淹性评估, 认为籼粳稻的萌发耐淹性差异并不显著, 与侯名语的结果不一致, 这可能是两者所用试验材料和方法不同造成的。随后其利用R0380/RP2334回交自交系群体在水稻2号、3号、8号染色体上检测到4个QTL, 贡献率为9.37%~17.34%。Jiang等[16]利用USSR5/N22 F2分离群体在水稻5号和11号染色体上检测到2个萌发耐淹性QTL, 贡献率为10.99%~15.51%。然而以上试验多是在直接淹水胁迫下进行的, 为了更准确的模拟大田淹水环境下种子的低氧萌发状态, 本研究采用土壤介质下的种子萌发耐淹性鉴定方法, 以胚芽鞘长度为指标进行萌发耐淹性种质资源筛选, 并进一步构建分离群体挖掘萌发耐低氧QTL, 为培育耐淹萌发能力强的水稻新品种提供参考。
1 材料与方法
1.1 种质资源
供试材料包括192份水稻种质资源, 其中191份为粳稻品种, 来自我国江苏、安徽、河南、山东、上海等地区, 1份为籼型常规水稻黄莉占, 来自广东省。定位群体来自以连粳15号(Lianjing 15, LJ15)为父本, 黄莉占(Huanglizhan, HLZ)为母本构建的F2:3分离群体, 该群体包含120个家系。为减少种子休眠带来的影响, 所有供试材料均在50℃高温条件下处理7 d以打破种子休眠。1.2 试验方法
模拟大田萌发耐淹鉴定: 每品种或家系挑选健康、饱满和均一的种子90粒, 分3次重复, 在2% H2O2中灭菌15 min, 随后用纯水冲洗5次, 浸种48 h后, 每重复挑选20粒吸足水分的种子, 播种于含有50个方孔的秧盘中, 秧盘中盛满细土, 播种深度1 cm, 方孔规格为5.5 cm × 5.5 cm × 5.0 cm, 每个方孔底部均有直径为0.8 cm的小圆孔, 以便排水, 秧盘置于57 cm × 38 cm × 9 cm 规格的周转箱中, 随后向周转箱中注入纯水至水面距离土壤表面5 cm, 每天注水以保持此高度不变, 以萌发7 d的胚芽鞘长度(coleoptile length, CL)作为萌发耐淹性的调查指标, 每重复随机取12粒种子测量胚芽鞘长度, 精确到1 mm。随机区组排列, 剔除极大值和极小值后, 取3次胚芽鞘长度平均值作为表型值进行品种比较分析、遗传分析和QTL检测。梯度试验: 分别设置9个不同的淹水深度对连粳15号和黄莉占进行萌发耐淹性测试, 水深分别为10、30、50、70、90、110、130、150和200 mm, 其余步骤同上。
以上试验均在温室中进行, 温度: (26±2)℃, 光∶暗 = 14∶10, 相对湿度80%左右。
1.3 遗传连锁图谱的构建及QTL分析
根据连粳15号/黄莉占F2:3分离群体的分子数据, 利用Mapmaker/EXP version 3.0软件(1.4 数据分析
利用Microsoft Excel 2010进行数据整理和频数分布直方图的绘制, SPSS 18.0软件进行方差、多重比较分析和KS-检验。2 结果与分析
2.1 萌发耐淹性种质资源筛选
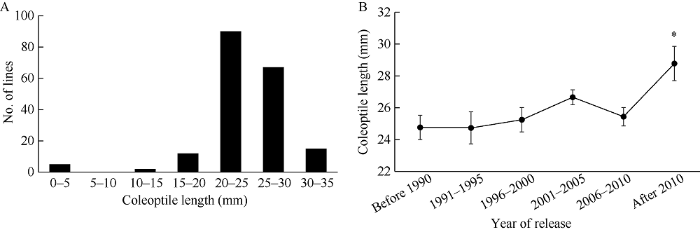
以萌发7 d的胚芽鞘长为指标, 本研究对来自不同年代和地区的191份粳稻种质资源进行萌发耐淹能力测试, 结果见附表1, 191份种质资源胚芽鞘长频数分布直方图呈偏态分布的特点(图1-A), 该群体的胚芽鞘长的变化范围为0~34.6 mm, 变异系数为21.8%, 表明这些种质资源在萌发耐淹性上具有广泛的遗传多样性。群体胚芽鞘长平均值为24.1 mm, 其中胚芽鞘长度小于20 mm种质资源总共有19份, 总占比9.9%, 处于20~25 mm之间的品种最多, 有90份, 占比47.1%, 其次为25~30 mm, 有67份, 占比35.1%, 其中 >30 mm品种有12个(表1), 占比6.3%, 这些种质表现出较强的耐低氧萌发能力, 可作为亲本材料进行萌发耐低氧新品种的培育。其中连云港地区的半野生稻种质资源穞稻表现最好, 胚芽鞘长34.6 mm, 其次为连粳15号, 胚芽鞘长34.2 mm。值得注意的是, 连粳15号(连粳7032/连粳446)的亲本之一连粳446也是由穞稻与连粳3号的杂交后代中选育而来, 推测连粳15号的萌发耐淹性大多来自穞稻, 因此, 应加大力度深入挖掘野生稻种的有利基因以增强水稻品种对生物胁迫和非生物胁迫的抗性。Table S1
附表1
附表1191份种质资源的萌发耐淹性表现
Table S1
| 编号 Number | 品种名称 Cultivar | 胚芽鞘长 Coleoptile length (mm) | 编号 Number | 品种名称 Cultivar | 胚芽鞘长 Coleoptile length (mm) |
|---|---|---|---|---|---|
| 1 | 早丰11号Zaofeng 11 | 0 | 97 | 扬农粳2号Yangnongjing 2 | — |
| 2 | 镇稻1号Zhendao 1 | 0 | 98 | 粳系103 Jingxi 103 | 24.6 |
| 3 | 早粳Zaojing | 0 | 99 | 台东育30 Taidongyu 30 | 24.6 |
| 4 | 梧桐Wutong | 0 | 100 | 盐粳11号Yanjing 11 | 24.7 |
| 5 | ITA182 | 0 | 101 | 扬粳4038 Yangjing 4038 | 24.7 |
| 6 | 临稻10号Lindao 10 | 13.4 | 102 | ZBS622 | 24.7 |
| 7 | 华粳4号Huajing 4 | 13.6 | 103 | 盐粳30192 Yanjing 30192 | 24.7 |
| 8 | HP3 | 17.2 | 104 | 盐稻15 Yandao 15 | 24.8 |
| 9 | 丹东陆稻Dandongludao | 17.7 | 105 | 小黄稻Xiaohuangdao | 24.8 |
| 10 | 绍糯9714 Shaonuo9714 | 17.7 | 106 | 武运粳8号Wuyunjing 8 | 24.8 |
| 11 | 南京11号 Nanjing 11 | 18.5 | 107 | ZBS665 | 24.8 |
| 12 | 淮稻13号Huaidao 13 | 18.7 | 108 | 泗稻10号Sidao 10 | 24.8 |
| 13 | 三光稻Sanguangdao | 18.8 | 109 | 香糯8333 Xiangnuo 8333 | 24.9 |
| 14 | 盐稻10号Yandao 10 | 19.0 | 110 | 湖粳75 Hujing 75 | 25.0 |
| 15 | 凤凰稻Fenghuangdao | 19.1 | 111 | 畚禾Benhe | 25.0 |
| 16 | 矮粳10号 Aijing 10 | 19.2 | 112 | 新宁Xinning | 25.2 |
| 17 | 旱糯谷Hannuogu | 19.3 | 113 | 南粳40 Nanjing 40 | 25.2 |
| 18 | 武运粳7号Wuyunjing 7 | 19.5 | 114 | 临稻18 Lindao 18 | 25.2 |
| 19 | 冀粳14 Jijing 14 | 19.9 | 115 | 高雄育122 Gaoxiongyu 122 | 25.2 |
| 20 | 南粳36 Nanjing 36 | 20.1 | 116 | 农虎早Laohuzao | 25.3 |
| 21 | 黄枝糯Huangzhinuo | 20.2 | 117 | 连粳3号Lianjing 3 | 25.4 |
| 22 | 垦鉴稻10号Kenjiandao 10 | 20.3 | 118 | DY158 | 25.4 |
| 23 | 台东陆稻328 Taidongludao 328 | 20.8 | 119 | 中粳区6号Zhongjingqu 6 | 25.5 |
| 24 | 淮稻11号Huaidao 11 | 20.8 | 120 | 盐粳4号Yanjing 4 | 25.5 |
| 25 | 南粳37 Nanjing 37 | 20.9 | 121 | 镇稻18 Zhendao 18 | 25.5 |
| 26 | 小黄早Xiaohuangzao | 20.9 | 122 | 越粳618 Yuejing 618 | 25.6 |
| 27 | Y136 | 20.9 | 123 | 淮糯12 Huainuo 12 | 25.6 |
| 28 | 扬粳9538 Yangjing 9538 | 21.0 | 124 | 镇稻88 Zhendao 88 | 25.6 |
| 29 | 扬粳687 Yangjing 687 | 21.1 | 125 | 秀水21 Xiushui 21 | 25.7 |
| 30 | 武育粳20号Wuyujing 20 | 21.2 | 126 | 南粳43 Nanjing 43 | 25.9 |
| 31 | 通科粳Tongkejing | 21.2 | 127 | C418 | 25.9 |
| 32 | 小葱稻Xiaocongdao | 21.3 | 128 | 盐粳6号Yanjing 6 | 25.9 |
| 33 | 圣稻13 Shengdao 13 | 21.3 | 129 | 镇稻86 Zhendao 86 | 26.0 |
| 34 | 盐粳9号Yanjing 9 | 21.3 | 130 | 台东育66 Taidongyu 66 | 26.0 |
| 35 | 连粳8671 Lianjing8671 | 21.4 | 131 | 锅底黑Guodihei | 26.1 |
| 36 | 农林8号Nonglin 8 | 21.5 | 132 | 中粳区5号 Zhongjingqu 5 | 26.1 |
| 37 | 武育糯16号Wuyunuo 16 | 21.5 | 133 | 苏粳2号Sujing 2 | 26.1 |
| 38 | 京越1号Jingyue 1 | 21.6 | 134 | 桂花球Guihuaqiu | 26.2 |
| 39 | 新稻18 Xindao 18 | 21.7 | 135 | 连粳3号选系 Line of Lianjing 3 | 26.2 |
| 40 | 矮秆黄Aiganhuang | 21.8 | 136 | 黄谷粳稻Huanggujingdao | 26.2 |
| 41 | 香稻Xiangdao | 21.9 | 137 | 连粳5号Lianjing 5 | 26.3 |
| 42 | 迟粳预1号Chijingyu 1 | 22.0 | 138 | 华粳3号Huajing 3 | 26.3 |
| 43 | 葫芦稻Huludao | 22.1 | 139 | 桂花黄Guihuahuang | 26.4 |
| 44 | 双城糯Shuangchengnuo | 22.1 | 140 | 新粳1号Xinjing 1 | 26.4 |
| 45 | 苏州选271 Suzhouxuan 271 | 22.1 | 141 | 白壳糯 1 Baikenuo 1 | 26.5 |
| 46 | 武糯5系Wunuo 5 | 22.3 | 142 | 盐粳5号Yanjing 5 | 26.5 |
| 47 | 中粳区7 Zhongjingqu 7 | 22.3 | 143 | 连粳2号Lianjing 2 | 26.5 |
| 48 | 黄粘粳Huangzhanjing | 22.4 | 144 | DY162 | 26.6 |
| 49 | 苏联种Sulianzhong | 22.4 | 145 | 淮稻8号Huaidao 8 | 26.7 |
| 50 | HR539 | 22.6 | 146 | 临稻4号Lindao 4 | 26.7 |
| 51 | 作作稻Zuozuodao | 22.7 | 147 | 淮优粳2号Huaiyoujing 2 | 26.8 |
| 52 | 早熟香Zaoshuxiang | 22.7 | 148 | 连粳6号Lianjing 6 | 26.9 |
| 53 | 盐稻7号Yanjing 7 | 22.7 | 149 | 徐稻3号Xudao 3 | 27.0 |
| 54 | 盐粳10号Yanjing 10 | 22.8 | 150 | 江北糯1号Jiangbeinuo 1 | 27.1 |
| 55 | 国优5号Guoyou 5 | 22.8 | 151 | Kinmaze | 27.3 |
| 56 | L12-3 | 22.9 | 152 | 郑稻18 Zhengdao 18 | 27.3 |
| 57 | 浙粳61 Zhejing 61 | 22.9 | 153 | 镇稻108 Zhendao 108 | 27.4 |
| 58 | 武香粳1号Wuxiangjing 1 | 22.9 | 154 | 剑粳6号Jianjing 6 | 27.4 |
| 59 | 盐粳30237 Yanjing 30237 | 22.9 | 155 | 扬辐粳8号Yangfujing 8 | 27.5 |
| 60 | HP5 | 23.0 | 156 | 扬糯2号Yangnuo 2 | 27.5 |
| 61 | 红壳糯Hongkenuo | 23.0 | 157 | R0380 | 27.5 |
| 62 | 辽粳287 Liaojing 287 | 23.1 | 158 | DY110 | 27.7 |
| 63 | 武育粳18号Wuyujing 18 | 23.3 | 159 | 扬粳1号Yangjing 1 | 27.8 |
| 64 | 南粳46 Nanjing 46 | 23.3 | 160 | 淮稻9号Huaidao 9 | 27.9 |
| 65 | 扬粳4227 Yangjing 4227 | 23.3 | 161 | 盐稻8号Yandao 8 | 28.0 |
| 66 | L1346 | 23.3 | 162 | 淮稻7号Huaidao 7 | 28.0 |
| 67 | 盐粳16 Yanjing 16 | 23.4 | 163 | 苏香粳2号Suxiangjing 2 | 28.1 |
| 68 | 连16783 Lian 16783 | 23.4 | 164 | 泗稻12号Sidao 12 | 28.1 |
| 69 | ZBS152 | 23.5 | 165 | 早丰9号Zaofeng 9 | 28.2 |
| 70 | L168-3 | 23.5 | 166 | 盐粳2号Yanjing 2 | 28.6 |
| 71 | 浙粳66 Zhejing 66 | 23.5 | 167 | 武粳15 Wujing 15 | 28.6 |
| 72 | 淮稻2号Huaidao 2 | 23.6 | 168 | 9805.0 | 28.7 |
| 73 | 南粳44 Nanjing 44 | 23.6 | 169 | 徐稻5号Xudao 5 | 28.8 |
| 74 | 东道Dongdao | 23.7 | 170 | 徐稻6号Xudao 6 | 28.9 |
| 75 | HN363 | 23.7 | 171 | 镇稻2号Zhendao 2 | 28.9 |
| 76 | 常农粳4号Changnongjing 4 | 23.8 | 172 | 南粳41 Nanjing 41 | 29.0 |
| 77 | 秀水04 Xiushui 04 | 23.9 | 173 | 宁粳1号Ningjing 1 | 29.1 |
| 78 | 中粳区8号 Zhongjingqu 8 | 24.0 | 174 | 武运粳11号Wuyunjing 11 | 29.3 |
| 79 | 武育粳3号Wuyujing 3 | 24.0 | 175 | 临糯Linnuo | 29.3 |
| 80 | ZBS153 | 24.0 | 176 | 华粳6号Huajing 6 | 29.4 |
| 81 | 小香稻Xiaoxiangdao | 24.0 | 177 | 连粳4号Lianjing 4 | 29.5 |
| 82 | 新稻10号Xindao 10 | 24.1 | 178 | 盐稻9号Yandao 9 | 29.8 |
| 83 | 吾别Wubie | 24.1 | 179 | 镇稻99 Zhendao 99 | 29.8 |
| 84 | 矮城804 Aicheng 804 | 24.2 | 180 | 临稻11号Lindao 11 | 30.1 |
| 85 | 花糯Huanuo | 24.2 | 181 | 淮稻6号Huaidao 6 | 30.4 |
| 86 | 奎稻Kuidao | 24.2 | 182 | 华粳5号Huajing 5 | 30.5 |
| 87 | 盐稻6号Yandao 6 | 24.4 | 183 | 扬粳186 Yangjing 186 | 30.7 |
| 88 | 宁粳3号Ningjing 3 | 24.4 | 184 | 徐稻4号Xudao 4 | 30.9 |
| 89 | 武育粳7号Wuyujing 7 | 24.4 | 185 | 淮稻10号Huaidao 10 | 31.1 |
| 90 | 宁粳2号Ningjing 2 | 24.4 | 186 | 连粳7号Lianjing 7 | 31.1 |
| 91 | 武运粳21 Wuyunjing 21 | 24.5 | 187 | 连粳9号Lianjing 9 | 32.0 |
| 92 | 武香粳14 Wuxiangjing 14 | 24.5 | 188 | 盐粳7号Yanjing 7 | 32.2 |
| 93 | 盐糯12 Yannuo 12 | 24.5 | 189 | 南粳45 Nanjing 45 | 32.5 |
| 94 | 昆稻选7号Kundaoxuan 7 | 24.5 | 190 | 连粳15号Lianjing 15 | 34.2 |
| 95 | 农虎禾-3 Nonghuhe-3 | 24.5 | 191 | 穞稻Ludao | 34.6 |
| 96 | 五优稻1号Wuyoudao 1 | 24.5 |
新窗口打开|下载CSV
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1种质资源萌发耐淹性评价
A: 淹水条件下, 191份粳稻种质资源萌发耐淹性表型分布直方图; B: 淹水条件下, 不同年代种质资源胚芽鞘长度比较分析。
Fig. 1Evaluation for germinability of germplasm resources under submerged condition
A: Frequency distribution of 191 japonica germplasm resources for germinability under submerged condition; B: Comparative analysis of coleoptile length of germplasm resources released from different years under submerged condition.
Table 1
表1
表112份萌发耐淹性强的种质资源
Table 1
| 编号 Number | 品种名称 Cultivar | 胚芽鞘长度 Coleoptile length (mm) | 编号 Number | 品种名称 Cultivar | 胚芽鞘长度 Coleoptile length (mm) |
|---|---|---|---|---|---|
| 1 | 穞稻 Ludao | 34.6 | 7 | 淮稻10号 Huaidao 10 | 31.1 |
| 2 | 连粳15号 Lianjing 15 | 34.2 | 8 | 徐稻4号 Xudao 4 | 30.9 |
| 3 | 南粳45 Nanjing 45 | 32.5 | 9 | 扬粳186 Yangjing 186 | 30.7 |
| 4 | 盐粳7号 Yanjing 7 | 32.2 | 10 | 华粳5号 Huajing 5 | 30.5 |
| 5 | 连粳9号 Lianjing 9 | 32.0 | 11 | 淮稻6号 Huaidao 6 | 30.4 |
| 6 | 连粳7号 Lianjing 7 | 31.1 | 12 | 临稻11号 Lindao 11 | 30.1 |
新窗口打开|下载CSV
2.2 不同年代种质资源萌发耐淹性比较分析
根据品种审定年份将83份种质资源按照5年一个阶段划分为6组, 分别为1990年前、1991—1995年、1996—2000年、2001—2005年、2006—2010年和2010年后, 通过萌发耐淹性鉴定, 6个时间段品种的平均胚芽鞘长分别为24.8、24.7、25.3、26.7、25.4和28.8 mm (图1-B), 经过多重比较分析发现, 前5个时间段, 品种之间的胚芽鞘长没有显著性差异, 但均与2010年以后品种的胚芽鞘长达到显著性差异, 这表明育种家们已开始加强对萌发耐淹性品种的选育。2.3 连粳15号萌发耐淹性验证
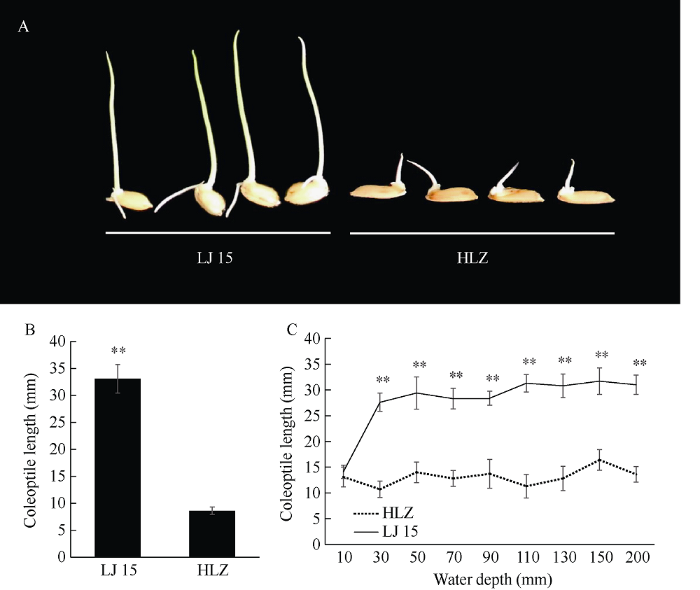
通过对191份种质资源进行筛选鉴定, 粳稻品种连粳15号在淹水胁迫下的胚芽鞘长为34.2 mm, 表现出较强的耐低氧发芽力, 为了验证该表型, 我们重新对其进行鉴定, 连粳15号胚芽鞘长为33.1 mm, 籼稻品种黄莉占胚芽鞘长为8.6 mm, 显著低于连粳15号(图2-A, B); 同时, 梯度试验表明, 在水深10 mm时, 连粳15号与黄莉占的胚芽鞘长无显著性差异, 因为此时淹水深度较浅, 2个品种的胚芽鞘均能很快伸出水面接触到氧气, 完成正常生长, 而在其他淹水处理下, 连粳15号的胚芽鞘长均显著高于黄莉占(图2-C)。因此, 连粳15号和黄莉占可作为理想的材料进行萌发耐淹性QTL检测。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2连粳15号和黄莉占的萌发耐淹性表现
A: 在淹水条件下, 连粳15号与黄莉占胚芽鞘长表型图; B: 在淹水条件下, 连粳15号与黄莉占的平均胚芽鞘长; C: 连粳15号与黄莉占在不同淹水深度条件下的胚芽鞘长度比较分析。
Fig. 2Germinability of Lianjing15 (LJ15) and Huanglizhan (HLZ) under submerged condition
A: Phenotypic image of coleoptile length between LJ15 and HLZ under submerged condition; B: Average coleoptile length of LJ15 and HLZ under submerged condition; C: Comparative analysis of coleoptile length between LJ15 and HLZ under different water depth.
2.4 萌发耐淹性QTL检测
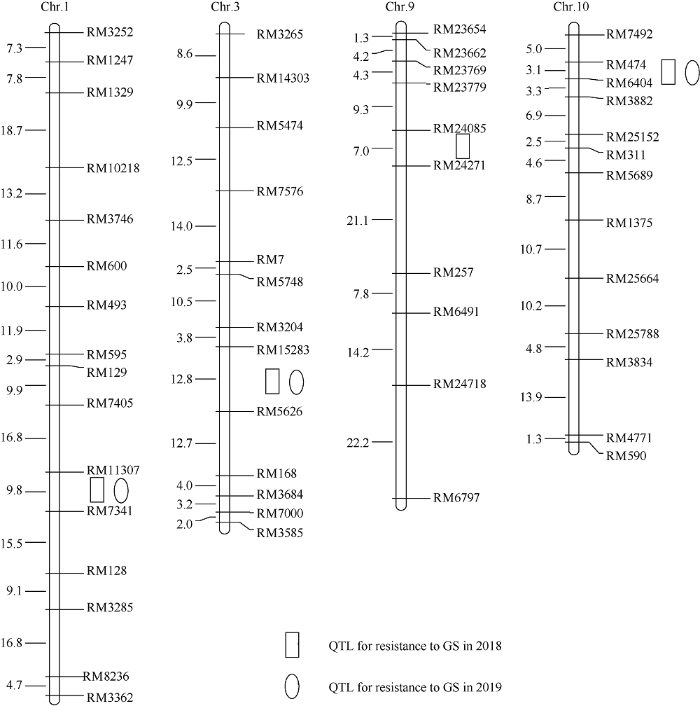
利用512对分布于12条水稻染色体上的SSR (simple sequence repeat, SSR)标记进行分离群体双亲间的多态性分析, 其中在连粳15号与黄莉占之间呈现良好多态性的SSR标记共有146对, 占所有检测标记的28.5%, 随后利用这些标记构建连粳15号/黄莉占F2:3群体的分子连锁图谱, 图谱全长为1262.4 cM, 标记之间的平均遗传距离为8.6 cM, 符合QTL检测要求。为了检测萌发耐淹性QTL, 本研究于2018年对连粳15号、黄莉占及其F2:3群体进行萌发耐淹性鉴定, 连粳15号和黄莉占的胚芽鞘长分别为31.9 mm和5.0 mm, 两者之间存在极显著差异(图3-A), 连粳15号/黄莉占F2:3分离群体胚芽鞘长的变化范围为0~31.5 mm, 在20~25 mm处出现1个峰值, 呈典型的正态分布特征(KS-检验, P > 0.05), 表明该群体的萌发耐淹性由多个QTL共同控制。以平均胚芽鞘长为表型值, 利用Windows QTL Cartographer 2.5软件采用复合区间作图法进行分析, 在水稻1号、3号、9号和10号染色体上各检测到1个QTL, 分别命名为qGS1、qGS3、qGS9和qGS10, 分别坐落于标记RM11307~RM7341、RM15283~RM5626、RM24085~RM24271和RM474~ RM6404之间, LOD值分别为6.7、3.6、3.3和6.1, 贡献率分别为24.0%、14.7%、11.7%和20.5%, 共解释表型变异的70.9% (表2和图4), 加性效应分析表明, 这些增强萌发耐淹性的QTL均来自粳稻品种连粳15号。
图3
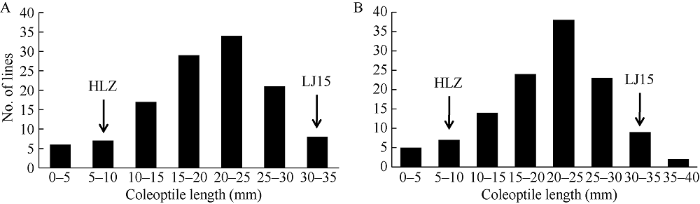 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3连粳15号/黄莉占F2:3群体萌发耐淹性表型分布直方图
A: 2018年; B: 2019年。
Fig. 3Frequency distribution of germinability under submerged condition (GS) in an F2:3 population derived from the cross between Lianjing 15 (LJ15) and Huangzhan (HLZ)
GS: germinability under submerged condition. A: in 2018; B: in 2019.
Table 2
表2
表2连粳15号/黄莉占F2:3群体中检测到的萌发耐淹性QTL
Table 2
| 年份 Year | 数量性状位点 QTL | 染色体 Chr. | 标记区间 Marker interval | LOD值 LOD scores | 贡献率 PVE (%) | 加性效应 Additive effect |
|---|---|---|---|---|---|---|
| 2018 | qGS1 | 1 | RM11307-RM7341 | 6.7 | 24.0 | -0.53 |
| qGS3 | 3 | RM15280-RM7134 | 3.6 | 14.7 | -0.43 | |
| qGS9 | 9 | RM24085-RM24271 | 3.3 | 11.7 | -0.32 | |
| qGS10 | 10 | RM474-RM6404 | 6.1 | 20.5 | -0.50 | |
| 2019 | qGS1 | 1 | RM11307-RM7341 | 5.1 | 19.2 | -0.49 |
| qGS3 | 3 | RM15280-RM7134 | 3.0 | 12.6 | -0.41 | |
| qGS10 | 10 | RM474-RM6404 | 5.6 | 19.1 | -0.50 |
新窗口打开|下载CSV
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4连粳15号/黄莉占F2:3群体中检测到的萌发耐淹性QTL
染色体右边是分子标记名称, 左边是遗传距离(cM)。
Fig. 4QTLs for germinability under submerged condition (GS) detected in LJ15/HLZ F2:3 population
Marker names are given on the right of the chromosome and map distances (cM) on the left. PVE: phenotypic variation explaned.
2.5 萌发耐淹性QTL的验证
为进一步验证以上QTL的真实性, 本研究于2019年再次对120个连粳15号/黄莉占F2:3家系及其双亲进行表型鉴定(图3-B), 结合分子连锁图谱, 利用Windows QTL Cartographer 2.5软件对以上位点进行了重新检测, 结果表明, qGS1、qGS3和qGS10均能够被重新检测到, LOD值分别为5.1、3.0和5.6, 贡献率分别为19.2%、12.6%和19.1% (表2和图4), 以上结果证明这些QTL是稳定表达真实存在的。3 讨论
水稻直播作为一种轻简、高效、节水、省工的栽培方式, 越来越受到种植者的欢迎[17]。然而, 在淹水胁迫下, 特别是当种子播在较深土层时, 由于氧气浓度过低导致出苗率急剧下降, 严重制约着直播稻的发展[18]。多项研究表明, 种子萌发耐淹性存在着品种间差异[19,20,21], 选择低氧萌发能力强的稻种是直播稻生产中提高出苗率的一个关键问题。因此, 筛选萌发耐淹性强的种质资源, 挖掘低氧耐受基因, 并解析其低氧适应机制已迫在眉睫。本研究通过对191份粳稻种质资源进行萌发耐淹性鉴定, 发现该性状的频率分布直方图呈现偏态分布特点, 说明该性状受到育种家的人工选择, 其中胚芽鞘长为20~25 mm的种质资源占比最多, 达到47.1%, 该群体变异系数为21.8%, 表明粳稻品种的萌发耐淹性存在广泛的遗传变异, 与前人研究结果一致[12,16,22]。经过筛选, 最终获得12份萌发耐淹性强的水稻种质(胚芽鞘长>30 mm) (表1), 这些资源可作为亲本材料用来选育萌发耐低氧的水稻品种。通过不同年代品种间的比较分析, 我们发现2010年后审定的品种平均胚芽鞘长度为28.8 mm, 相对于前期品种, 在低氧胁迫下的萌发能力显著性提高, 这表明随着耕作栽培制度的改变, 育种家们为了适应以直播稻为主的轻简栽培的需要, 已开始加强对品种的萌发耐淹性状的选择。同时我们也比较了不同种植区域种质资源的耐淹萌发能力, 发现并没有达到显著的地域性差异, 这说明在不同区域环境条件下, 萌发耐淹性是水稻在驯化扩散过程中的一个基本特性。
前人研究已经表明, 萌发耐淹性是由多基因控制的数量性状[14,16,23-24], 本研究通过资源鉴定, 发现粳稻品种连粳15号表现出较强的低氧萌发能力, 利用其与籼稻品种黄莉占的F2:3群体共检测到4个QTL, 再次验证了该观点。孙凯等[14]利用全基因组关联分析, 在水稻3号、4号、5号、6号、8号、10号和11号染色体上共检测到15个与胚芽鞘长度显著关联的位点, 其中位于3号染色体上的位点在本研究检测位点qGS3两端标记区间内, 这2个QTL很可能是同一个位点, 随后其通过转录组测序发现该位点区间内基因Os03g0592500对氧气处理敏感, 该基因编码一个叶绿素a-b结合蛋白, 并推测该基因对水稻种子耐淹生长具有正调控作用。王洋等[24]利用粳稻Nipponbare与籼稻Kasalath BIL群体也在水稻3号染色体上检测到1个耐低氧发芽能力QTL, qSAT-3-B, 与qGS3的物理位置十分接近, 是否为同一个QTL尚需进一步确认。目前前人已克隆2个耐淹基因, CIPK15和Sub1, 其中, CIPK15在种子萌发阶段通过糖信号途径产生更多能量促使胚芽鞘快速生长以应对淹水胁迫[25], 而Sub1则通过调节乙烯和赤霉素介导的反应, 抑制水稻淹水期的营养生长, 以节约能量促进去淹没后的恢复生长来达到提高水稻耐淹性的目的[26]。通过比对分析, 这2个基因均在水稻9号染色体上着丝粒附近, 与本研究检测到的qGS9在染色体上的距离超过1700 kb, 三者为不同的基因。王洋等[24]也在水稻9号染色体上检测到1个QTL, qSAT-9-B, 贡献率为9.5%, 与本研究检测到的qGS9有部分重叠区域, 可能为同一个QTL, 值得注意的是, qGS9未在连续的2年试验中重复检测到, 这可能是由于qGS9本身效应较小, 且受到环境影响, 表达不稳定造成。通过比对分析, 本研究检测到的位于水稻第1和10染色体上的QTL, qGS1和qGS10, 与前人报道萌发耐淹性QTL没有重叠区域, 是2个新的萌发耐淹性QTL位点, 而且这2个QTL都能被重复检测到, 2次检测中LOD值均大于5, 分别解释表型变异的19.2~24.0%, 19.1~20.5%, 是稳定表达的QTL。因此, 利用与之紧密连锁的分子标记, 通过分子标记辅助选择的手段聚合这些萌发耐淹性QTL, 将有助于加快培育耐低氧萌发水稻新品种。
4 结论
从191份粳稻中筛选获得12份萌发耐淹性强的种质资源, 并利用连粳15号/黄莉占F2:3群体, 共检测到4个萌发耐淹性QTL, qGS1、qGS3、qGS9和qGS10, 分别位于水稻1号、3号、9号和10号染色体上, 贡献率分别为24.0%、14.7%、11.7%和20.5%, 其中qGS1和qGS10真实可靠, 且贡献率较高, 这些QTL的定位为进一步开展相关基因的图位克隆和分子辅助育种奠定基础。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s13593-014-0239-0URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.1111/j.1438-8677.2008.00150.xURLPMID:19538394 [本文引用: 2]

Rice (Oryza sativa L.) seeds can germinate under anoxia and can show coleoptile elongation. The anoxic coleoptile is usually longer than aerobic coleoptiles. Although several hypotheses have been proposed to explain the ability of rice to elongate coleoptiles under anoxia, conclusive experimental evidence explaining this physiological trait is lacking. In order to investigate whether metabolic and molecular markers correlate with anoxic coleoptile length, we screened 141 Italian and 23 Sri Lankan rice cultivars for their ability to elongate coleoptiles under anoxia. Differences in anoxic coleoptile length were used to evaluate whether a correlation exists between coleoptile length and biochemical and molecular parameters. The expression of genes coding for glycolytic and fermentative enzymes showed a very low correlation with anoxic coleoptile length. Although differences were found in carbohydrate content between the varieties tested, this parameter also does not appear to be critical in terms of coleoptile elongation. Efficient ethanol fermentation does, however, correlate well with the elongation of coleoptiles under anoxic conditions.
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
DOI:10.1126/scisignal.2000333URLPMID:19809091 [本文引用: 1]
Flooding is a widespread natural disaster that leads to oxygen (O(2)) and energy deficiency in terrestrial plants, thereby reducing their productivity. Rice is unusually tolerant to flooding, but the underlying mechanism for this tolerance has remained elusive. Here, we show that protein kinase CIPK15 [calcineurin B-like (CBL)-interacting protein kinase] plays a key role in O(2)-deficiency tolerance in rice. CIPK15 regulates the plant global energy and stress sensor SnRK1A (Snf1-related protein kinase 1) and links O(2)-deficiency signals to the SnRK1-dependent sugar-sensing cascade to regulate sugar and energy production and to enable rice growth under floodwater. Our studies contribute to understanding how rice grows under the conditions of O(2) deficiency necessary for growing rice in irrigated lowlands.
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}