 ,1,3,*, 王克如4, 李少昆4, 姜闯道,2,*
,1,3,*, 王克如4, 李少昆4, 姜闯道,2,*Photosynthetic characteristics of senescent leaf induced by high planting density of maize at heading stage in the field
WU Han-Yu1,2, ZHANG Ya-Jun2, ZHANG Wang-Feng,1,3,*, WANG Ke-Ru4, LI Shao-Kun4, JIANG Chuang-Dao,2,*通讯作者:
第一联系人:
收稿日期:2018-05-3接受日期:2018-10-8网络出版日期:2018-11-05
| 基金资助: |
Received:2018-05-3Accepted:2018-10-8Online:2018-11-05
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (553KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
吴含玉, 张雅君, 张旺锋, 王克如, 李少昆, 姜闯道. 田间密植诱导抽穗期玉米叶片衰老时的光合作用机制[J]. 作物学报, 2019, 45(2): 248-255. doi:10.3724/SP.J.1006.2019.83042
WU Han-Yu, ZHANG Ya-Jun, ZHANG Wang-Feng, WANG Ke-Ru, LI Shao-Kun, JIANG Chuang-Dao.
作物栽培中, 一定程度的密植能够有效提高单位面积生物量与产量。但是, 密植也会导致个体间的资源竞争, 尤其是光照资源。众所周知, 光不仅是光合作用的能量来源, 也是影响植物生长发育的重要环境因子。我们前期研究表明, 提高种植密度会加剧植株间的相互遮阴, 限制单株对光能的截获与利用, 影响叶片的形态结构, 并导致单叶光合速率降低, 进而影响作物的生长发育[1,2]。更重要的是, 密植条件下的弱光环境还会影响甚至加剧叶片衰老。而衰老是影响作物产量的重要因素, 延缓衰老可以一定程度上提高作物产量。
叶片衰老过程中, 尽管气孔敏感性会随之降低, 但光合速率下降的主要原因被认为是非气孔限制[3,4]。实际上, 除气孔外, 光合作用包括光能捕获、电子传递和碳同化等步骤。有研究认为, 叶片衰老过程中LHCa、LHCb、叶绿素生物合成基因(CHLM、PROB、PROC)表达量下降而叶绿素分解基因(SGR、NYC1)表达量增加[5,6]。叶绿素含量的降低会直接影响光能捕获。此外, 在叶片衰老过程中光合作用中光系统I和II相关蛋白以及叶绿体ATP酶表达量均呈现下降趋势[7]。所以, 有研究认为光合电子传递功能的衰退是叶片衰老过程中光合能力迅速下降的重要原因[8]。但是, 随着叶片衰老, Rubisco和Rubisco活化酶的含量减少[9,10,11]; 同时, 两者活性均下降[3,12]。而且, Rubisco基因表达及大小亚基含量的下降也能够抑制Rubisco合成。因此, 这些研究认为暗反应碳同化能力降低可能是导致衰老过程中光合速率下降的关键因素[13,14,15,16]。
尽管前人已经就作物衰老对光合作用的影响进行了大量研究, 但这些实验大多是室内诱导叶片衰老[3,5-6,17-18]。其中, 有些是离体条件下诱导叶片衰老[6]。由于离体叶片不能与植物体进行正常的物质和信息交换, 因此离体叶片的研究可能会反映衰老的部分规律, 但不能准确反应植物活体衰老情况。另外, 我们也注意到在活体研究中多数是使用幼苗诱导衰老。因为幼苗代谢旺盛, 其在生理、代谢和基因表达上均与成熟植株有明显差异, 所以幼苗期很可能也未必能够准确地反映田间成熟植株的衰老[19]。截至目前, 仅有少数实验揭示了成熟植株的衰老规律[16]。但这些模拟研究使用的是均一的光环境诱导叶片衰老, 与田间情况不完全一致。我们的近期研究表明, 作物密植条件下冠层光环境的异质性增强, 即冠层上部暴露在强光下, 而下部则处于弱光下[1,2]。一方面, 异质性光环境中的弱光可能导致叶片衰老; 另一方面, 植物也可能会通过调节光合作用适应异质性光环境。但田间成熟植物在这种复杂的光环境下是否会衰老或适应, 以及衰老或适应相关的光合作用调节机制尚不清楚。
玉米是主要的粮食作物之一。近年来, 我国的玉米高产研究主要以提高栽培密度为基础。实际上, 在抽穗期玉米刚刚完成营养生长并形成冠层郁闭, 此时田间密植导致的异质性光环境是否诱导了玉米抽穗期叶片衰老, 密植诱导玉米叶片衰老并引起光合速率下降的原因是什么, 是否与前人的模拟研究结果一致?因此, 本实验测定抽穗期不同种植密度下的光强、比叶重、氮素含量、叶绿素含量、气体交换以及荧光诱导动力学的变化, 旨在揭示田间抽穗期密植诱导叶片衰老的规律, 以期为我国玉米的高产稳产提供实验支持。
1 材料与方法
1.1 材料种植与实验设计
2017年在新疆奇台进行试验, 以玉米先玉335 (XY335)为试材。奇台总场海拔750~851 m, 日照时数2500~2800 h, 太阳总辐射量546.3 kJ cm-2, 年≥10℃积温3000~3200℃ d, 无霜期130~156 d, 年降雨180~480 mm。先玉335其母本为PH6WC, 父本为 PH4CV。先玉335: 熟期适宜, 产量高, 稳产性好; 容重高, 含水量低, 商品品质好; 适应性广, 抗逆性强, 适合机械化收获。近几年我国北方因先玉335迅猛扩展, 其种植面积已居全国第二。根据栽培密度与产量的关系, 设置低密度(15,000株 hm-2)、中密度(75,000株 hm-2)和高密度(135,000株 hm-2) 3种处理。通过膜下滴灌进行正常水肥管理。在抽穗期, 分别针对穗位叶和穗下第4叶进行各种测定。1.2 光强测定
用冠层分析仪(Sunscan, Delta-T Devices, 英国)在7月上午10:00到14:00测定穗位叶和穗下第4叶的光强, 每个处理至少测定5个重复。1.3 比叶重测定
选择大小一致的穗位叶和穗下第4叶, 用剪刀剪出边长为3.5 cm的叶片16片, 放入105℃烘箱杀青30 min, 85℃烘干至恒重。用电子天平测得正方形叶片的总干重, 计算比叶重(比叶重=叶干重/叶面积)。1.4 叶片氮素含量测定
用混合研磨仪把样品磨碎, 用电子天平秤取8 mg左右, 用元素分析仪(Vario EL III, elementar, 德国)测定穗位叶和穗下第4叶的氮含量和碳含量。1.5 叶绿体色素含量测定
用打孔器在穗位叶和穗下第4叶分别打5个叶圆片(直径1 cm)放入刻度试管中, 分别加入80%丙酮溶液10 mL。每个处理至少3次重复, 以80%丙酮为对照。将试管放入黑暗中浸提48 h, 每12 h振荡一次, 至叶圆片绿色完全褪去后振荡混匀, 用分光光度计(UV-1800、中国)测定663 nm、645 nm、440 nm处的吸光度, 带入公式计算色素含量[20]。1.6 气体交换参数
使用便携式光合作用系统(CIRAS-2, PP Systems, 美国)在7月份10:00至14:00测定穗位叶和穗下第4叶的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(E)和胞间CO2浓度(Ci)。每个处理选取至少5个叶片。测定时, 光强为2000 μmol m-2 s-1, 二氧化碳浓度控制在(380±20) μmol mol-1, 湿度控制在75%±5%, 叶室温度为环境温度。1.7 叶绿素荧光测定
参照李鹏民等[21]测定方法, 选用连续激发式荧光仪(Handy-PEA, Hansatech, 英国)测定玉米穗位叶和穗下第4叶的荧光参数。待晚上22:00 30 min暗适应后测定荧光诱导动力学, 并计算光系统II (PSII)最大光化学效率(Fv/Fm)等参数。1.8 数据处理
采用SPSS统计分析数据, SigmaPlot 12.5软件作图。2 结果与分析
2.1 密植对冠层内光强和比叶重的影响
图1-A表明, 在玉米的穗位叶和穗下第4叶, 光强随密度升高而显著下降。与低密度相比, 穗位叶光强在中密度和高密度分别降低了13.9%和29.3%, 穗下第4叶光强分别降低了63.4%和87.9%。在同一密度下, 与穗位叶相比, 低密度、中密度和高密度穗下第4叶光强分别下降了44.3%、76.4%和90.5%。说明密植明显降低冠层内的光强, 尤其是下部叶片的光强。图1
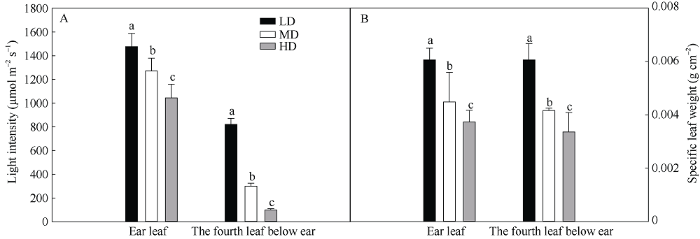 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1栽培密度对抽穗期玉米穗位叶和穗下第4叶光强(A)和比叶重(B)的影响
LD: 低密度; MD: 中密度; HD: 高密度。柱上不同字母表示不同处理差异在0.05水平显著。
Fig. 1Effects of planting density on light intensity (A) and specific leaf weight (B) of ear leaf and the fourth leaf below ear of maize at heading period
LD: low planting density; MD: medium planting density; HD: high planting density. Bars superscripted by different letters are significantly different at the 0.05 probability level among different treatments.
比叶重是衡量叶片厚度的一个重要参数。图1-B表明, 穗位叶和穗下第4叶随密度增加比叶重降低, 说明密植使叶片变薄。与低密度相比, 中密度和高密度条件下穗位叶比叶重分别下降了26.1%和38.5%, 穗下第4叶中分别下降了31.3%和44.6%。在同一种植密度下, 与穗位叶相比, 低密度、中密度和高密度穗下第4叶的比叶重分别下降了0.02%、7.1%和9.9%。上述结果说明密植降低了比叶重。
2.2 密植对叶片氮素和叶绿素含量的影响
图2-A表明, 在玉米的穗位叶和穗下第4叶, 叶片含氮量随密度升高而显著下降。与低密度相比, 穗位叶单位面积氮含量在中密度和高密度分别降低了23.9%和41.1%, 穗下第4叶氮含量分别降低了38.0%和54.5%。在同一密度下, 与穗位叶相比, 低密度、中密度和高密度穗下第4叶单位面积氮含量分别下降了5.3%、22.9%和26.8%。这些数据说明了密植明显降低叶片单位面积氮素含量。图2
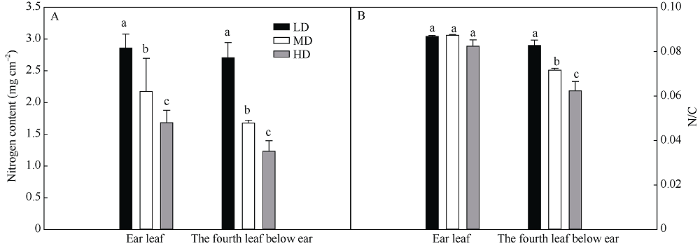 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2栽培密度对抽穗期玉米穗位叶和穗下第4叶氮素含量(A)和氮/碳比值(B)的影响
LD: 低密度; MD: 中密度; HD: 高密度。柱上不同字母表示不同处理差异在0.05水平显著。
Fig. 2Effects of planting density on nitrogen content and nitrogen/carbon ratio of ear leaf and the fourth leaf below ear of maize at heading period
LD: low planting density; MD: medium planting density; HD: high planting density. Bars superscripted by different letters are significantly different at the 0.05 probability level among different treatments.
图2-B表明, 在玉米的穗位叶, N/C随密度增加没有发生显著变化。在穗下第4叶, N/C随密度升高而显著下降。与低密度相比, 穗下第4叶N/C分别降低了13.4%和24.6%。在同一密度下, 与穗位叶相比, 低密度、中密度和高密度穗下第4叶N/C分别下降了3.8%、17.9%和24.3%。
除叶片氮含量外, 叶绿素含量也发生了明显改变。图3表明, 随密度增加穗位叶和穗下第4叶叶绿素a (图3-A)、叶绿素b (图3-B)、类胡萝卜素含量(图3-C)、叶绿素总量(图3-D)均降低。在穗位叶中, 与低密度相比, 中密度和高密度的单位面积叶绿素a的含量分别下降了14.4%和23.7%, 单位面积总叶绿素含量分别下降了17.1%和25.2%; 在穗下第4叶中, 与低密度相比, 中密度和高密度的单位面积叶绿素a含量分别下降了25.2%和25.8%, 单位面积总叶绿素含量分别下降了23.7%和26.3%。在同一种植密度下, 与穗位叶相比, 低密度、中密度和高密度穗下第4叶的叶绿素a含量分别下降了6.3%、18.1%和8.9%, 穗下第4叶叶绿素总量分别下降了5.5%、13.0%和6.9%。说明密植降低了叶绿素含量。
图3
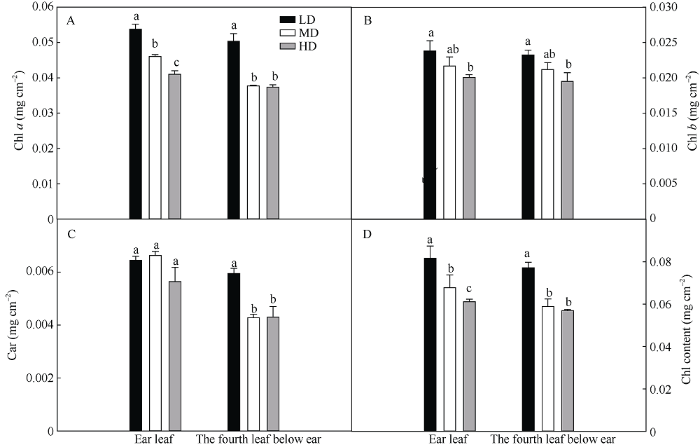 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3栽培密度对抽穗期玉米穗位叶和穗下第4叶的叶绿素a (A)、叶绿素b (B)、类胡萝卜素含量(C)和叶绿素总量(D)的影响
LD: 低密度; MD: 中密度; HD: 高密度。柱上不同字母表示不同处理差异在0.05水平显著。
Fig. 3Effects of planting density on chlorophyll a (A), chlorophyll b (B), carotenoid (C), and total chlorophyll contents (D) of ear leaf and the fourth leaf below ear of maize at heading period
LD: low planting density; MD: medium planting density; HD: high planting density. Bars superscripted by different letters are significantly different at the 0.05 probability level among different treatments.
2.3 密植对荧光诱导动力学的影响
快速叶绿素荧光诱导动力学曲线(OJIP 曲线)中蕴含着 PSII 原初光化学反应的丰富信息, 被广泛应用于 PSII 活性研究中。在标准化的OJIP曲线中(图4), 不同密度条件下穗位叶的OJIP曲线十分相似; 而穗下第4叶OJIP曲线明显变化。与低密度相比, 穗下第4叶在高密度条件下OJIP曲线的J点(2 ms)和I点(30 ms)荧光都明显上升(图4-B)。图4
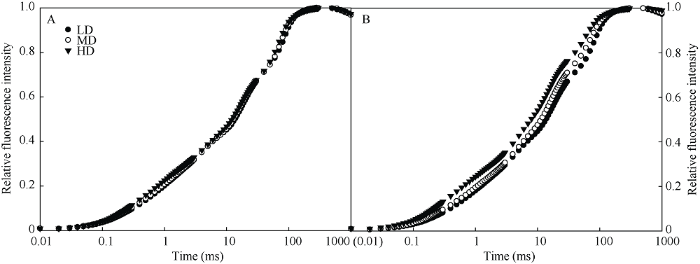 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4栽培密度对抽穗期玉米穗位叶(A)和穗下第4叶(B)叶绿素荧光诱导动力学的影响
LD: 低密度; MD: 中密度; HD: 高密度。
Fig. 4Effects of planting density on chlorophyll a fluorescence transient(OJIP) of ear leaf (A) and the fourth leaf below ear (B) of maize at heading period
LD: low planting density; MD: medium planting density; HD: high planting density.
随着密度增加, 穗位叶PSII最大光化学效率几乎未发生变化(图5-A), 穗下第4叶PSII最大光化学效率有轻微的下降(图5-A)。说明在这一时期密植导致的衰老对PSII最大光化学效率影响比较小。ψEo表示反应中心吸收的光能用于电子传递的量子产额, 能够反映PSII电子传递的效率。在穗位叶和在穗下第4叶中, 随密度增加, 电子传递效率均变化很小。与低密度相比, 中密度和高密度穗位叶的电子传递效率分别下降0.47%和2.90%, 中密度和高密度穗下第4叶的电子传递效率分别下降1.20%和8.00%。说明密植能够一定程度上影响玉米穗下第4叶的电子传递效率。
图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5栽培密度对抽穗期玉米穗位叶和穗下第4叶荧光参数的影响
LD: 低密度; MD: 中密度; HD: 高密度。柱上不同字母表示不同处理差异在0.05水平显著。
Fig. 5Effects of planting density on fluorescence parameters of ear leaf and the fourth leaf below ear of maize at heading period
LD: low planting density; MD: medium planting density; HD: high planting density. Bars superscripted by different letters are significantly different at the 0.05 probability level among different treatments.
2.4 密植对气体交换的影响
随密度增加, 穗位叶和穗下第4叶的光合速率(图6-A)、气孔导度(图6-B)和蒸腾速率(图6-C)下降。但在穗位叶中胞间二氧化碳浓度呈下降趋势(图6-D), 而在穗下第4叶中轻微升高(图6-D)。与低密度相比, 穗位叶在中密度和高密度条件下光合速率分别下降了7.6%和14.8%, 穗下第4叶分别下降了20.2%和30.0%。在同一种植密度下, 与穗位叶相比, 低密度、中密度和高密度的穗下第4叶的光合速率分别下降了13%、25%和28%。说明密植降低了碳同化能力。图6
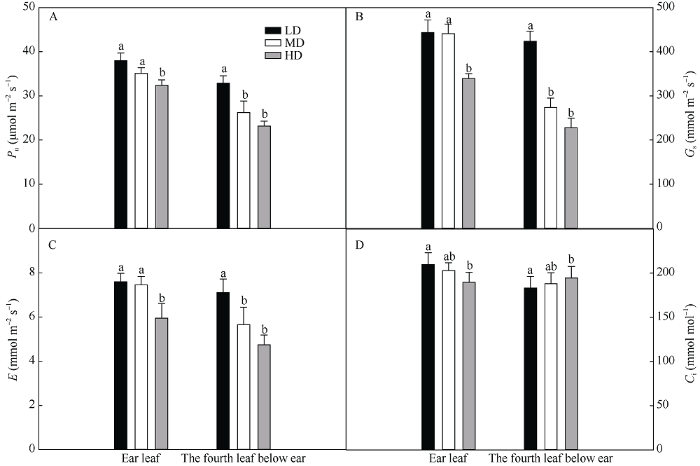 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6栽培密度对抽穗期玉米穗位叶和穗下第4叶气体交换参数的影响
(A)净光合速率; (B)气孔导度; (C)蒸腾速率; (D)细胞间隙二氧化碳浓度。LD: 低密度; MD: 中密度; HD: 高密度。柱上不同字母表示不同处理差异在 0.05 水平显著。
Fig. 6Effects of planting density on gas exchange parameters of ear leaf and the fourth leaf below ear of maize at heading period
(A): net photosynthetic rate; (B): stomatal conductance; (C): transpiration rate; (D): intercellular CO2 concentration. LD: low planting density; MD: medium planting density; HD: high planting density. Bars superscripted by different letters are significantly different at the 0.05 probability level among different treatments.
3 讨论
3.1 密植条件下异质性光环境诱导玉米叶片衰老
密植会导致玉米群体冠层中下部光强减弱, 增强光环境的异质性(图1-A)。本研究中, 穗下第4叶片的光合有效辐射更低, 在栽培密度为中密度(75,000株 hm-2)时光强约为300 μmol m-2 s-1, 而在高密度(135,000株 hm-2)时光强为100 μmol m-2 s-1左右。尽管穗位叶的比叶重有一定程度的降低, 但是其光合速率的降低幅度很小; 而且, 穗位叶的荧光诱导动力学曲线变化也很小。所以, 密植条件下玉米穗位叶能够维持很高的光合电子传递和光合能力。不过, 我们也观察到密植过程中伴随穗位叶光合速率的降低, 气孔导度和细胞间隙二氧化碳浓度也有轻微下降趋势, 所以抽穗期玉米穗位叶光合速率的降低表现为一定程度的气孔限制; 同时, 穗位叶叶绿素含量也轻微降低。这些数据暗示抽穗期穗位叶开始轻微衰老, 或者说田间条件下衰老最初表现是叶绿素降解和光合速率下降。尽管不同密度间穗位叶的光合特性差异较小, 但密植条件下穗下第4叶片差异非常显著(图6-A)。与穗位叶相比, 随着栽培密度的增加和光强减弱, 穗下第4叶的比叶重和光合速率均大幅降低, 但胞间CO2浓度未随之大幅下降, 说明密植条件下该叶片光合速率的下降主要是非气孔限制, 即叶肉限制的结果。
本研究中, 高密度下穗下第4叶片的光合速率和叶绿素含量均明显降低。大量研究表明, 弱光下植物叶片可以通过提高单位面积的叶绿素含量, 增加光能捕获, 从而维持光合能力, 进而适应弱光环境。数据表明, 在田间水肥供应充足条件下伴随栽培密度的提高穗位叶的叶绿素含量不仅没有增加, 反而有下降趋势, 穗下第4叶片的下降趋势尤为明显。这显然与植物对弱光环境的适应不符。如果植物在饱和光强下叶片也不能够捕获足够的光能用于光合作用, 会导致光合速率和叶绿素比值大幅增加, 也就意味着色素含量会限制光合作用。为排除叶绿素含量降低对光合速率的可能影响, 计算穗下第4叶片光合速率与叶绿素含量的比值(Pn/Chl)。我们发现随着栽培密度增加Pn/Chl分别为4.3、4.6和4.2, 变化很小。因此, 叶绿素含量可能不是导致穗下第4叶片饱和光合速率降低的主要因素, 可能只是光合速率降低的伴随过程。此外, 穗下第4叶片荧光诱导动力学曲线的J和I相的相对荧光产量仅略有提高。与低密度相比, 中密度和高密度的电子传递效率分别下降1.2%和8.0%, 表明PSII电子传递活性也不是穗下第4叶片光合速率下降的限制因素。
尽管密植条件下抽穗期的穗位叶和穗下第4叶都能够维持一定的光合速率, 但其单位面积的叶片氮素都降低。不过, 随着栽培密度提高穗位叶的N/C比基本没有变化, 只有穗下第4叶片该比值大幅下降, 证明后者损失了更多的氮素。氮素在光合作用, 尤其是暗反应中起非常重要的作用[22]。因此, 我们认为密植条件下抽穗期穗位叶可能以光合作用适应为主; 相反, 穗下第4叶片主要表现衰老。这也说明密植能够诱导抽穗期玉米冠层下部叶片衰老。穗下第4叶片一方面叶片年龄较穗位叶大; 另一方面, 其在密植条件下所处光强更弱。所以, 密植条件下冠层下部叶片的衰老应该与其异质性光环境有关。
3.2 密植条件下玉米植株对异质性光环境的适应
在低密度(15,000株 hm-2)条件下, 穗位叶和穗下第4叶片在比叶重、叶绿素含量、气体交换和荧光诱导动力学方面均差异非常小; 然而, 随着栽培密度的增加, 其形态和生理功能之间的差异呈增加趋势。这与其冠层光环境的异质性增强是一致的。而穗位叶饱和光合速率差异相对较小, 所以我们认为冠层下部叶片的光合功能对单株产量的形成也发挥重要作用, 而且这可能是密植条件下玉米单株产量降低的重要原因之一。但是, 在高密度条件下, 加快冠层下部叶片衰老并维持冠层上部叶片的光合功能可能也是植株个体对异质性光环境的一种适应。有研究表明, 先玉335株型紧凑、耐密、适应性广、产量高、适合机械化收获。因此, 该品种是近年来国内很多地区的密植主推品种。与其他株型松散的品种相比, 先玉335密植高产的原因可能是通过改善冠层下部叶片的光环境, 一定程度减缓了下部叶片的衰老, 从而使单株产量不至于大幅降低。
4 结论
田间密植条件下, 异质性光环境可以诱导抽穗期玉米冠层下部叶片衰老; 叶片衰老过程中, 光合作用的限制因素不是光能捕获和电子传递, 而可能是碳同化。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
URL [本文引用: 2]

比较研究了不同栽培密度对薄荷生长策略及光合特性的影响。结果表明:(1)随着密度的升高,单株的株高、茎粗、分枝数、单叶面积、叶片厚度等下降;(2)提高栽培密度时,薄荷的光合色素含量、光合面积、净光合速率均降低,而PSII的最大光化学效率不变;(3)高密度下,单株的根、茎、叶及总生物量积累下降,而群体的生物量积累变化不明显,但地下部分的生物量分配比例均显著增加。进一步分析表明密植会通过降低薄荷单位面积的色素含量而导致光合速率下降:栽培密度提高时薄荷通过增加根生物量比来优先竞争矿质营养。
URL [本文引用: 2]
比较研究了不同栽培密度对薄荷生长策略及光合特性的影响。结果表明:(1)随着密度的升高,单株的株高、茎粗、分枝数、单叶面积、叶片厚度等下降;(2)提高栽培密度时,薄荷的光合色素含量、光合面积、净光合速率均降低,而PSII的最大光化学效率不变;(3)高密度下,单株的根、茎、叶及总生物量积累下降,而群体的生物量积累变化不明显,但地下部分的生物量分配比例均显著增加。进一步分析表明密植会通过降低薄荷单位面积的色素含量而导致光合速率下降:栽培密度提高时薄荷通过增加根生物量比来优先竞争矿质营养。
DOI:10.1016/j.plaphy.2015.05.008URLPMID:26057699 [本文引用: 2]
61Photosynthesis is regulated systemically in field-grown sorghum.61The characteristics of systemic regulation of photosynthesis in the field are emphasized.61Leaf nitrogen content may not play a key role in systemic regulation of photosynthesis.61Mutual shading among plants induces optimization of photosynthetic performance.
DOI:10.1111/ppl.12587URLPMID:28543370 [本文引用: 3]
Abstract Rubisco activase (RCA) is an important enzyme that can catalyze the carboxylation and oxygenation activities of ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco), which is involved in the photosynthetic carbon reduction cycle. Here, we studied the effects of changes in RCA activity on photosynthesis, growth and development, as well as the chilling tolerance of RCA overexpressing transgenic cucumber (Cucumis sativus) plants. CsRCA overexpression increased the plant height, leaf area and dry matter, and decreased the root/top ratio in transgenic cucumber plants compared with the wild type (WT) plants. Low temperature stress led to decreases in the CsRCA expression and protein levels, the photosynthetic rate (Pn) and the stomatal conductance, but an increase in the intercellular CO2 concentration in cucumber leaves. The actual photochemical efficiency and maximal photochemical efficiency of Photosystem II in cucumber seedlings also declined, but the initial fluorescence increased during chilling stress. Transgenic plants showed a lower decrease in the CsRCA expression level and actual and maximal photochemical efficiencies, as well as increases in the intercellular CO2 concentration and initial fluorescence relative to the WT plants. Low temperature stress resulted in a significant increase in the malondialdehyde (MDA) content; however, this increase was reduced in transgenic plants compared with that in WT plants. Thus, the overexpression of CsRCA may promote the growth and chilling tolerance of cucumber plants in solar greenhouses.
DOI:10.1093/jxb/eri279URLPMID:16157653 [本文引用: 1]
Although leaf senescence results in a loss of photosynthetic carbon fixation, the senescence-dependent release of nutrients, especially of nitrogen, is important for the growth of young leaves and for reproduction. Environmental regulation of senescence is therefore a vital factor in the carbon and nitrogen economy of plants. Leaf senescence is a highly plastic trait that is affected by a range of different environmental factors including light, nutrient supply, CO2 concentration, and abiotic and biotic stress. In this review, the focus is on the impact of environmental conditions on sugar accumulation and sugar signalling during senescence. By signalling a high availability of carbon relative to nitrogen in the old leaves, sugar accumulation can trigger leaf senescence. Sugar-induced senescence is therefore particularly important under low nitrogen availability and may also play a role in light signalling. Whether or not sugars are involved in regulating the senescence response of plants to elevated CO2 remains unresolved. Senescence can be delayed or accelerated in elevated CO2 and no clear relationship between sugar accumulation and senescence has been found. Plasticity in the response to environmental factors, such as daylength and sugar accumulation, varies between different Arabidopsis accessions. This natural variation can be exploited to analyse the genetic basis of the regulation of senescence and the consequences for growth and fecundity. Different evolutionary strategies, i.e. early senescence combined with a high reproductive effort or late senescence combined with a low reproductive effort, may be an important adaptation of Arabidopsis accessions to their natural habitat.
DOI:10.1093/jxb/eru060URLPMID:4106438 [本文引用: 2]
Phytochrome is thought to control the induction of leaf senescence directly, however, the signalling and molecular mechanisms remain unclear. In the present study, an ecophysiological approach was used to establish a functional connection between phytochrome signalling and the physiological processes underlying the induction of leaf senescence in response to shade. With shade it is important to distinguish between complete and partial shading, during which either the whole or only a part of the plant is shaded, respectively. It is first shown here that, while PHYB is required to maintain chlorophyll content in a completely shaded plant, only PHYA is involved in maintaining the leaf chlorophyll content in response to partial plant shading. Second, it is shown that leaf yellowing associated with strong partial shading in phyA-mutant plants actually correlates to a decreased biosynthesis of chlorophyll rather than to an increase of its degradation. Third, it is shown that the physiological impact of this decreased biosynthesis of chlorophyll in strongly shaded phyA-mutant leaves is accompanied by a decreased capacity to adjust the Light Compensation Point. However, the increased leaf yellowing in phyA-mutant plants is not accompanied by an increase of senescence-specific molecular markers, which argues against a direct role of PHYA in inducing leaf senescence in response to partial shade. In conclusion, it is proposed that PHYA, but not PHYB, is essential for fine-tuning the chlorophyll biosynthetic pathway in response to partial shading. In turn, this mechanism allows the shaded leaf to adjust its photosynthetic machinery to very low irradiances, thus maintaining a positive carbon balance and repressing the induction of leaf senescence, which can occur under prolonged periods of shade.
DOI:10.1007/s11099-009-0093-5URL [本文引用: 3]
Ten light-harvesting complex (Lhc) proteins were investigated to determine which was the most appropriate protein marker of senescence in detached rice leaves. The levels of Lhc proteins were monitored by immunoblot analysis, which was conducted using commercially available antibodies raised against each Lhc protein. Among the Lhc proteins evaluated in this study, Lhca1, Lhcb1, Lhcb2, Lhcb3, and Lhcb5 were not appropriate to be used as senescence markers while others can be used after optimization of the procedure.
URL [本文引用: 1]
大田烟叶生产过程中因打顶打叉的处理,改变了烟叶正常的衰老模式。为研究这一特殊的衰老机制,我们自旺长期开始,对‘云烟87'不同发育阶段烟株的中部叶片,进行形态观测、生理生化分析及蛋白质组学检测。结果显示:随着烟叶的逐渐成熟和衰老,烟草的叶色逐渐变黄,叶片逐渐变短、变窄,厚度减少;解剖结构清晰看到栅栏组织和海绵组织从最初的整齐排列到逐渐排列紊乱,组织细胞间轮廓不明显,细胞间隙明显增大;亚显微观测表明,淀粉粒在叶绿体中逐渐积累,类囊体片层结构被挤散,叶绿体膜被撑破。生理与生化分析表明衰老过程伴随着光合作用速率下降,光合色素降解加速,呼吸代谢的增加,这可能与衰老叶片中叶绿体逐渐崩塌和细胞膜透性增加相一致。iTRAQ标记方法共检测到不同发育阶段432个差异表达蛋白质,其中注释到308个与多种生命过程相关。蛋白差异富集分析表明,烟草叶片衰老过程中与光合作用等合成代谢相关蛋白多下调表达,而逆境反应及呼吸作用等分解代谢相关蛋白多上调表达。
URL [本文引用: 1]
大田烟叶生产过程中因打顶打叉的处理,改变了烟叶正常的衰老模式。为研究这一特殊的衰老机制,我们自旺长期开始,对‘云烟87'不同发育阶段烟株的中部叶片,进行形态观测、生理生化分析及蛋白质组学检测。结果显示:随着烟叶的逐渐成熟和衰老,烟草的叶色逐渐变黄,叶片逐渐变短、变窄,厚度减少;解剖结构清晰看到栅栏组织和海绵组织从最初的整齐排列到逐渐排列紊乱,组织细胞间轮廓不明显,细胞间隙明显增大;亚显微观测表明,淀粉粒在叶绿体中逐渐积累,类囊体片层结构被挤散,叶绿体膜被撑破。生理与生化分析表明衰老过程伴随着光合作用速率下降,光合色素降解加速,呼吸代谢的增加,这可能与衰老叶片中叶绿体逐渐崩塌和细胞膜透性增加相一致。iTRAQ标记方法共检测到不同发育阶段432个差异表达蛋白质,其中注释到308个与多种生命过程相关。蛋白差异富集分析表明,烟草叶片衰老过程中与光合作用等合成代谢相关蛋白多下调表达,而逆境反应及呼吸作用等分解代谢相关蛋白多上调表达。
DOI:10.17957/IJABURL [本文引用: 1]
DOI:10.1111/j.1365-3040.2011.02474.xURLPMID:22171633 [本文引用: 1]
Plants often have to cope with altered light conditions, which in leaves induce various physiological responses ranging from photosynthetic acclimation to leaf senescence. However, our knowledge of the regulatory pathways by which shade and darkness induce leaf senescence remains incomplete. To determine to what extent reduced light intensities regulate the induction of leaf senescence, we performed a functional comparison between Arabidopsis leaves subjected to a range of shading treatments. Individually covered leaves, which remained attached to the plant, were compared with respect to chlorophyll, protein, histology, expression of senescence-associated genes, capacity for photosynthesis and respiration, and light compensation point (LCP). Mild shading induced photosynthetic acclimation and resource partitioning, which, together with a decreased respiration, lowered the LCP. Leaf senescence was induced only under strong shade, coinciding with a negative carbon balance and independent of the red/far-red ratio. Interestingly, while senescence was significantly delayed at very low light compared with darkness, phytochrome A mutant plants showed enhanced chlorophyll degradation under all shading treatments except complete darkness. Taken together, our results suggest that the induction of leaf senescence during shading depends on the efficiency of carbon fixation, which in turn appears to be modulated via light receptors such as phytochrome A.
DOI:10.1007/s12298-012-0142-6URLPMID:24381436 [本文引用: 1]
AbstractNatural leaf senescence was investigated in four rainfed lowland rice cultivars, FR 13A (tolerant to submergence), Sabita and Sarala (adapted to medium depth, 0 50 cm stagnant flooding) and Dengi (conventional farmers cultivar). Changes in the levels of pigment content, CO photosynthetic rate, photosystem II photochemistry and anti-oxidant enzyme activities of flag leaves during grain-filling stage were investigated. Chlorophyll content, photochemical efficiency of photosystem II and CO photosynthetic rate decreased significantly with the progress of grain-filling. Likely, the activities of antioxidant enzymes namely, superoxide dismutase, catalase, guaiacol peroxidase and ascorbate peroxidase decreased with progress of grain-filling. A substantial difference was observed among the four cultivars for the sustainability index (SI) of different photosynthetic parameters and antioxidant enzyme activities; SIs of those parameters, in general, were lower in low yielding cultivar FR 13A compared to the other three cultivars. Among the four cultivars Sabita gave maximum grain yield. Yet, SI of Pn was greater in Sarala and Dengi compared to the Sabita. SIs of electron transport (ETo/CS), maximal photochemical efficiency (Fv/Fm), area above Fo and Fm, catalase and ascorbate peroxidase were also greater in Sarala and Dengi. The data showed that among the different Chl a fluorescence parameters, PI could be used with greater accuracy to distinguish slow and fast senescence rice cultivars during grain-filling period. It was concluded that maintaining the vitality of rice plants during grain-filling gave guarantee to synthesize carbohydrate, however greater yield could be realized provided superior yield attributing parameters are present.
.
DOI:10.1111/j.1744-7909.2005.00098.xURL [本文引用: 1]
Abstract: Gas exchange, chlorophyll (Chl) fluorescence, photosynthetic pigments, and the expression of Rubisco and Rubisco activase in flag leaves of rice (Oryza sativa L.) from the heading to mature grain stage were investigated. The results showed that the photosynthetic capacity declined after full expansion of flag leaves. The decline of photosynthetic rate (Pn) in two cultivars during natural senescence was accompanied by a decrease in Chl content, as well as in the Rubisco and Rubisco activase contents. The Rubisco and Rubisco activase contents in Zhenong 966 decreased faster than that in Zhenong 952. The Pn diminished without a decrease in intercellular CO2 concentration during the early senescence of flag leaves, indicating that lower photosynthetic capacity is not caused by stomatal limitation but by reduced carboxylation efficiency. During the senescence of flag leaves, the correlation between the change in photosynthetic capacity and the variation in the abundance of Rubisco and Rubisco activase suggested that the decline in Pn of flag leaves could be attributed to the lower level of rbcS and rca transcripts. The PSII and the electron transport rate appeared in the same rate as Pn. However, excitation pressure (1-qP) showed a different pattern and there was an inverse linear correlation between increased excitation pressure and the reduced Rubisco. Therefore, it is likely that the increased excitation pressure resulted from saturation of the electron transport chain, owing to a limitation of the reductant used by the Calvin cycle; in addition, the change in excitation pressure could further mediate the expression of the rbcS and rca genes, resulting in a fast reduction in Rubisco content. (Managing editor: Ping HE)
DOI:10.1104/pp.109.145862URLPMID:19880611 [本文引用: 1]
While interspecific variation in the temperature response of photosynthesis is well documented, the underlying physiological mechanisms remain unknown. Moreover, mechanisms related to species-dependent differences in photosynthetic temperature acclimation are unclear. We compared photosynthetic temperature acclimation in 11 crop species differing in their cold tolerance, which were grown at 15°C or 30°C. Cold-tolerant species exhibited a large decrease in optimum temperature for the photosynthetic rate at 360 μL L6301 CO60 concentration [Opt (A616468)] when growth temperature decreased from 30°C to 15°C, whereas cold-sensitive species were less plastic in Opt (A616468). Analysis using the C61 photosynthesis model shows that the limiting step of A616468 at the optimum temperature differed between cold-tolerant and cold-sensitive species; ribulose 1,5-bisphosphate carboxylation rate was limiting in cold-tolerant species, while ribulose 1,5-bisphosphate regeneration rate was limiting in cold-sensitive species. Alterations in parameters related to photosynthetic temperature acclimation, including the limiting step of A616468, leaf nitrogen, and Rubisco contents, were more plastic to growth temperature in cold-tolerant species than in cold-sensitive species. These plastic alterations contributed to the noted growth temperature-dependent changes in Opt (A616468) in cold-tolerant species. Consequently, cold-tolerant species were able to maintain high A616468 at 15°C or 30°C, whereas cold-sensitive species were not. We conclude that differences in the plasticity of photosynthetic parameters with respect to growth temperature were responsible for the noted interspecific differences in photosynthetic temperature acclimation between cold-tolerant and cold-sensitive species.
DOI:10.1016/S2095-3119(13)60670-XURL [本文引用: 1]
Low light stress is one of the most important factors affecting photosynthesis and growth in winter production of cucumber (Cucumis sativus L.) in solar greenhouses in northern China. Here, two genotypes of cucumber (Deltastar and Jinyan 2) are used to determine the effect of low light stress on Rubisco expression and photosynthesis of leaves from emergence to senescence. During leaf development, the net photosynthetic rate (PN), stomatal conductance (gs), Rubisco initial activity and activation state, transcript levels of rbcL and rbcS, and the abundance of rbcL and rbcS DNA in these two genotypes increase rapidly to reach maximum in 10-20 d, and then decrease gradually. Meanwhile, the actual photosystem II efficiency (肖PSII) of cucumber leaves slowly increased in the early leaf developing stages, but it declined quickly in leaf senescent stages, accompanied by an increased non-photochemical quenching (NPQ). Moreover, PN, gs, initial Rubisco activity, and abundance of protein, mRNA and DNA of Rubisco subunits of leaves grown under 100 mol m-2 s-1 are lower, and require more time to reach their maxima than those grown under 600 mol m-2 s-1 during leaf development. All these results suggest that lower photosynthetic capacity of cucumber leaves from emergence to senescence under low light stress is probably due to down-regulated Rubisco gene expression in transcript and protein levels, and decreased initial and total activity as well as activation state of Rubisco. Deltastar performs better than Jinyan 2 under low light stress.
DOI:10.11869/j.issn.100-8551.2017.06.1200URL [本文引用: 1]
为进一步探讨黄瓜耐弱光性的特性,对戴多星和津研2号2个黄瓜品种进行弱光胁迫处理(光照强度75~100μmol·m~(-2)·s~(-1)),以600μmol·m~(-2)·s~(-1)光照强度为对照,研究弱光胁迫下其生理生化指标、光合活性、荧光特性、Rubisco活性、Rubisco大小亚基蛋白含量及基因表达的变化。结果表明,弱光处理后,黄瓜气孔发育受到抑制,气孔开张度、气孔密度和气孔指数均有所降低,且津研2号表现更为明显,所有指标均降低20%以上。黄瓜叶片叶绿素含量、植株净光合速率(Pn),气孔导度(Gs),PSⅡ实际量子效率(ΦPSⅡ),Rubisco活力、Rubisco大小亚基蛋白含量及基因表达均明显降低,其中津研2号所有指标均下降30%以上。与津研2号相比,戴多星的色素含量较高,光能捕获和利用能力较强,分配于光合作用的量子产量较高,Rubisco活性、Rubisco大小亚基蛋白含量及大小亚基基因表达较高,因而Pn相对降低较少(30.9%,津研2号下降54.5%),表现出较强的耐弱光性。本研究结果为黄瓜耐弱光机理的深入研究及耐弱光黄瓜品种的选育栽培提供了参考依据。
DOI:10.11869/j.issn.100-8551.2017.06.1200URL [本文引用: 1]
为进一步探讨黄瓜耐弱光性的特性,对戴多星和津研2号2个黄瓜品种进行弱光胁迫处理(光照强度75~100μmol·m~(-2)·s~(-1)),以600μmol·m~(-2)·s~(-1)光照强度为对照,研究弱光胁迫下其生理生化指标、光合活性、荧光特性、Rubisco活性、Rubisco大小亚基蛋白含量及基因表达的变化。结果表明,弱光处理后,黄瓜气孔发育受到抑制,气孔开张度、气孔密度和气孔指数均有所降低,且津研2号表现更为明显,所有指标均降低20%以上。黄瓜叶片叶绿素含量、植株净光合速率(Pn),气孔导度(Gs),PSⅡ实际量子效率(ΦPSⅡ),Rubisco活力、Rubisco大小亚基蛋白含量及基因表达均明显降低,其中津研2号所有指标均下降30%以上。与津研2号相比,戴多星的色素含量较高,光能捕获和利用能力较强,分配于光合作用的量子产量较高,Rubisco活性、Rubisco大小亚基蛋白含量及大小亚基基因表达较高,因而Pn相对降低较少(30.9%,津研2号下降54.5%),表现出较强的耐弱光性。本研究结果为黄瓜耐弱光机理的深入研究及耐弱光黄瓜品种的选育栽培提供了参考依据。
DOI:10.1104/pp.107.099762URLPMID:17535822 [本文引用: 1]
Little is known about the effect of hormones on the photosynthetic process. Therefore, we studied Rubisco content and expression along with gas exchange parameters in transgenic tobacco (Nicotiana tabacum) plants that are not able to sense ethylene. We also tested for a possible interaction between ethylene insensitivity, abscisic acid (ABA), and sugar feedback on photosynthesis. We measured Rubisco content in seedlings grown in agar with or without added sugar and fluridone, and Rubisco expression in hydroponically grown vegetative plants grown at low and high CO . Furthermore, we analyzed gas exchange and the photosynthetic machinery of transformants and wild-type plants grown under standard conditions. In the presence of exogenous glucose (Glc), agar-grown seedlings of the ethylene-insensitive genotype had lower amounts of Rubisco per unit leaf area than the wild type. No differences in Rubisco content were found between ethylene-insensitive and wild-type seedlings treated with fluridone, suggesting that inhibition of ABA production nullified the effect of Glc application. When larger, vegetative plants were grown at different atmospheric CO concentrations, a negative correlation was found between Glc concentration in the leaves and Rubisco gene expression, with stronger repression by high Glc concentrations in ethylene-insensitive plants. Ethylene insensitivity resulted in plants with comparable fractions of nitrogen invested in light harvesting, but lower amounts in electron transport and Rubisco. Consequently, photosynthetic capacity of the insensitive genotype was clearly lower compared with the wild type. We conclude that the inability to perceive ethylene results in increased sensitivity to Glc, which may be mediated by a higher ABA concentration. This increased sensitivity to endogenous Glc has negative consequences for Rubisco content and photosynthetic capacity of these plants.
DOI:10.1007/s11738-014-1746-yURL [本文引用: 2]
A proteomic analysis combining photosynthesis-related key enzymes measurement and mRNA level analysis was used to investigate the variations of proteins involved in the photosynthetic apparatus in flag leaves of high-yield rice LYPJ during senescence under field conditions. The stability of chlorophyll-protein complexes of thylakoid membranes was measured by BN-PAGE system. Our findings revealed that the LHCII complexes were more stable than F 0 F 1 -ATPase and Cy b 6 /f complexes. Thylakoid membranes proteins during the senescence stage were separated by two-dimensional gel electrophoresis. We identified 13 photosynthesis-related proteins (a fivefold or greater variation), including electron transport, Calvin enson cycle and chloroplast protease during senescence of flag leaves. Moreover, our data showed that Ca 2+ -ATPase was more stable during senescence of flag leaves in rice, compared to Mg 2+ -ATPase. In enzymes activities analyses, it was observed that a little increment of C4 photosynthetic enzymes activities at the beginning of flag leaves senescence might compensate, to some degree, for the larger reductions in Rubisco activity, confirmed by quantitative RT-PCR analysis.
DOI:10.1111/j.1365-3040.2007.01724.xURLPMID:17986154 [本文引用: 1]
Senescence is an active process allowing the reallocation of valuable nutrients from the senescing organ towards storage and/or growing tissues. Using Arabidopsis thaliana leaves from both whole darkened plants (DPs) and individually darkened leaves (IDLs), we investigated the fate of mitochondria and chloroplasts during dark-induced leaf senescence. Combining in vivo visualization of fates of the two organelles by three-dimensional reconstructions of abaxial parts of leaves with functional measurements of photosynthesis and respiration, we showed that the two experimental systems displayed major differences during 6 d of dark treatment. In whole DPs, organelles were largely retained in both epidermal and mesophyll cells. However, while the photosynthetic capacity was maintained, the capacity of mitochondrial respiration decreased. In contrast, IDLs showed a rapid decline in photosynthetic capacity while maintaining a high capacity for mitochondrial respiration throughout the treatment. In addition, we noticed an unequal degradation of organelles in the different cell types of the senescing leaf. From these data, we suggest that metabolism in leaves of the whole DPs enters a 'stand-by mode' to preserve the photosynthetic machinery for as long as possible. However, in IDLs, mitochondria actively provide energy and carbon skeletons for the degradation of cell constituents, facilitating the retrieval of nutrients. Finally, the heterogeneity of the degradation processes involved during senescence is discussed with regard to the fate of mitochondria and chloroplasts in the different cell types.
DOI:10.1104/pp.010312URLPMID:11706170 [本文引用: 1]
It has long been known that leaf senescence can be induced in many plant species by detaching leaves and placing them in the darkness. It recently has been shown that entire Arabidopsis plants placed in the darkness are not induced to senesce, as judged by visible yellowing and certain molecular markers. Here, we show that when individual Arabidopsis leaves are darkened, but not when entire plants are darkened, senescence is induced in the covered leaves. This induction of senescence is highly localized. The phenomenon is leaf age dependent in that it occurs more rapidly and strongly in older leaves than in younger ones, as is the case with many forms of induced senescence. Whole adult plants placed in darkness, in contrast, show delayed senescence, although seedlings lacking primary leaves do not. These observations imply that the light status of the entire plant affects the senescence of individual leaves. A model summarizing the results is presented.
DOI:10.1111/pbi.12422URLPMID:26132509 [本文引用: 1]
Summary Leaf senescence is a complex process, which has dramatic consequences on crop yield. In sunflower, gap between potential and actual yields reveals the economic impact of senescence. Indeed, sunflower plants are incapable of maintaining their green leaf area over sustained periods. This study characterizes the leaf senescence process in sunflower through a systems biology approach integrating transcriptomic and metabolomic analyses: plants being grown under both glasshouse and field conditions. Our results revealed a correspondence between profile changes detected at the molecular, biochemical and physiological level throughout the progression of leaf senescence measured at different plant developmental stages. Early metabolic changes were detected prior to anthesis and before the onset of the first senescence symptoms, with more pronounced changes observed when physiological and molecular variables were assessed under field conditions. During leaf development, photosynthetic activity and cell growth processes decreased, whereas sucrose, fatty acid, nucleotide and amino acid metabolisms increased. Pathways related to nutrient recycling processes were also up-regulated. Members of the NAC, AP2-EREBP, HB, bZIP and MYB transcription factor families showed high expression levels, and their expression level was highly correlated, suggesting their involvement in sunflower senescence. The results of this study thus contribute to the elucidation of the molecular mechanisms involved in the onset and progression of leaf senescence in sunflower leaves as well as to the identification of candidate genes involved in this process.
DOI:10.1104/pp.24.1.1URLPMID:16654194 [本文引用: 1]
Publication » COPPER ENZYMES IN ISOLATED CHLOROPLASTS. POLYPHENOLOXIDASE IN BETA VULGARIS..
DOI:10.3321/j.issn:1671-3877.2005.06.001URL [本文引用: 1]
JIP-测定(JIP-test)是以生物膜能量流动为基础建立的分析方法。利用该方法可以获得有关光系统II的大量信息。文章介绍了快速叶绿素荧光诱导动力学曲线的定义、数据分析方法及相关参数的意义,并举例说明如何利用该方法分析不同环境条件对光合机构主要是PSII的供体侧、受体侧及PSII反应中心的影响。
DOI:10.3321/j.issn:1671-3877.2005.06.001URL [本文引用: 1]
JIP-测定(JIP-test)是以生物膜能量流动为基础建立的分析方法。利用该方法可以获得有关光系统II的大量信息。文章介绍了快速叶绿素荧光诱导动力学曲线的定义、数据分析方法及相关参数的意义,并举例说明如何利用该方法分析不同环境条件对光合机构主要是PSII的供体侧、受体侧及PSII反应中心的影响。
DOI:10.3724/SP.J.1006.2010.00871URL [本文引用: 1]
选用玉米品种先玉335、郑单958和吉单209,设计了6.0、7.5、9.0和10.5万株 hm-2 4个种植密度。测定了不同生育时期(吐丝期为重点)的叶片叶绿素含量、可溶性糖含量、全氮含量与碳氮比的动态变化。结果表明,3个品种的叶片叶绿素含量在灌浆期最高,可溶性糖含量在吐丝期最高,碳氮比在吐丝期与成熟期出现2个高峰;叶绿素含量、灌浆期光合速率、叶片可溶性糖、全氮、碳氮比均随种植密度增加而降低;吐丝后光合速率呈下降趋势;吐丝后生育天数与叶绿素含量、光合速率、可溶性糖、C/N呈二次曲线关系,与全氮含量呈线性关系;高种植密度主要影响叶片碳代谢;叶绿素含量不是影响吐丝后光合速率的主要因素。不同基因型耐密性不同,先玉335最好,郑单958次之,吉单209最差。
DOI:10.3724/SP.J.1006.2010.00871URL [本文引用: 1]
选用玉米品种先玉335、郑单958和吉单209,设计了6.0、7.5、9.0和10.5万株 hm-2 4个种植密度。测定了不同生育时期(吐丝期为重点)的叶片叶绿素含量、可溶性糖含量、全氮含量与碳氮比的动态变化。结果表明,3个品种的叶片叶绿素含量在灌浆期最高,可溶性糖含量在吐丝期最高,碳氮比在吐丝期与成熟期出现2个高峰;叶绿素含量、灌浆期光合速率、叶片可溶性糖、全氮、碳氮比均随种植密度增加而降低;吐丝后光合速率呈下降趋势;吐丝后生育天数与叶绿素含量、光合速率、可溶性糖、C/N呈二次曲线关系,与全氮含量呈线性关系;高种植密度主要影响叶片碳代谢;叶绿素含量不是影响吐丝后光合速率的主要因素。不同基因型耐密性不同,先玉335最好,郑单958次之,吉单209最差。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}