关键词:大豆; 籽粒; 维生素E含量; QTL定位 Identification of QTL Associated with Vitamin E Content in Soybean Seeds ZHANG Hong-Mei1, LI Hai-Chao2, WEN Zi-Xiang2, GU He-Ping1, YUAN Xing-Xing1, CHEN Hua-Tao1, CUI Xiao-Yan1, CHEN Xin1,*, LU Wei-Guo2,* 1Institute of Vegetable Crops, Jiangsu Academy of Agricultural Sciences, Nanjing 210014, China
2 Institute of Industrial Crops, Henan Academy of Agricultural Scienecs / Zhengzhou Subcenter of National Center for Soybean Improvement / Key laboratory of Oil Crops in Huanghuaihai Plains, Ministry of Agriculture, Zhengzhou 450002, China
AbstractVitamin E has effects on human immunity, anti-cancer and prevention of cardiovascular disease. Vitamin E from soybean has the advantages of higher security and higher human body absorption rate. The objective of the present study was to map the additive, additive × additive (epistasis), additive × year and epistasis × year QTLs for vitamin E and relative tocopherol contents with the RIL population BIEX (Essex×ZDD2315) using HPLC (high performance liquid chromatography) method and software QTLNetwork 2.1. Eight additive QTLs and twelve additive × additive (epistasis) QTLs were detected for vitamin E and relative tocopherol contents. The contributions to the phenotypic variances of additive QTL and epistatic QTL pairs were 8.68% (two QTLs) and 15.57% (four pairs) for α-tocopherol, 8.59% (two QTL) and 11.57% (two pairs) for γ-tocopherol, 5.44% (one QTL) and 17.61% (three pairs) for δ-tocopherol and 11.39% (three QTL) and 9.48% (three pairs) for total vitamin E contents, respectively. Those of additive and epistatic QTLs by year interaction were not found. The accumulated contribution of the unmapped minor QTLs was 66.16%-75.32%, indicating the variance of unmapped minor QTLs accounting for more than two thirds. In genetic composition, undetected minor QTLs accounted for a considerably large part additive QTLs and epistatic QTLs were nearly equal in α-tocopherol, γ-tocopherol, δ-tocopherol and total vitamin E contents. Accordingly, in breeding for vitamin E contents, the strategy of pyramiding multiple QTLs, both additive and epistatic, by using marker-assisted selection combined with accumulating minor effect QTLs through conversional procedures should be considered.
Keyword:Soybean [ Glycine max (L.) Merr.]; Seed; Vitamin E content; QTL mapping Show Figures Show Figures
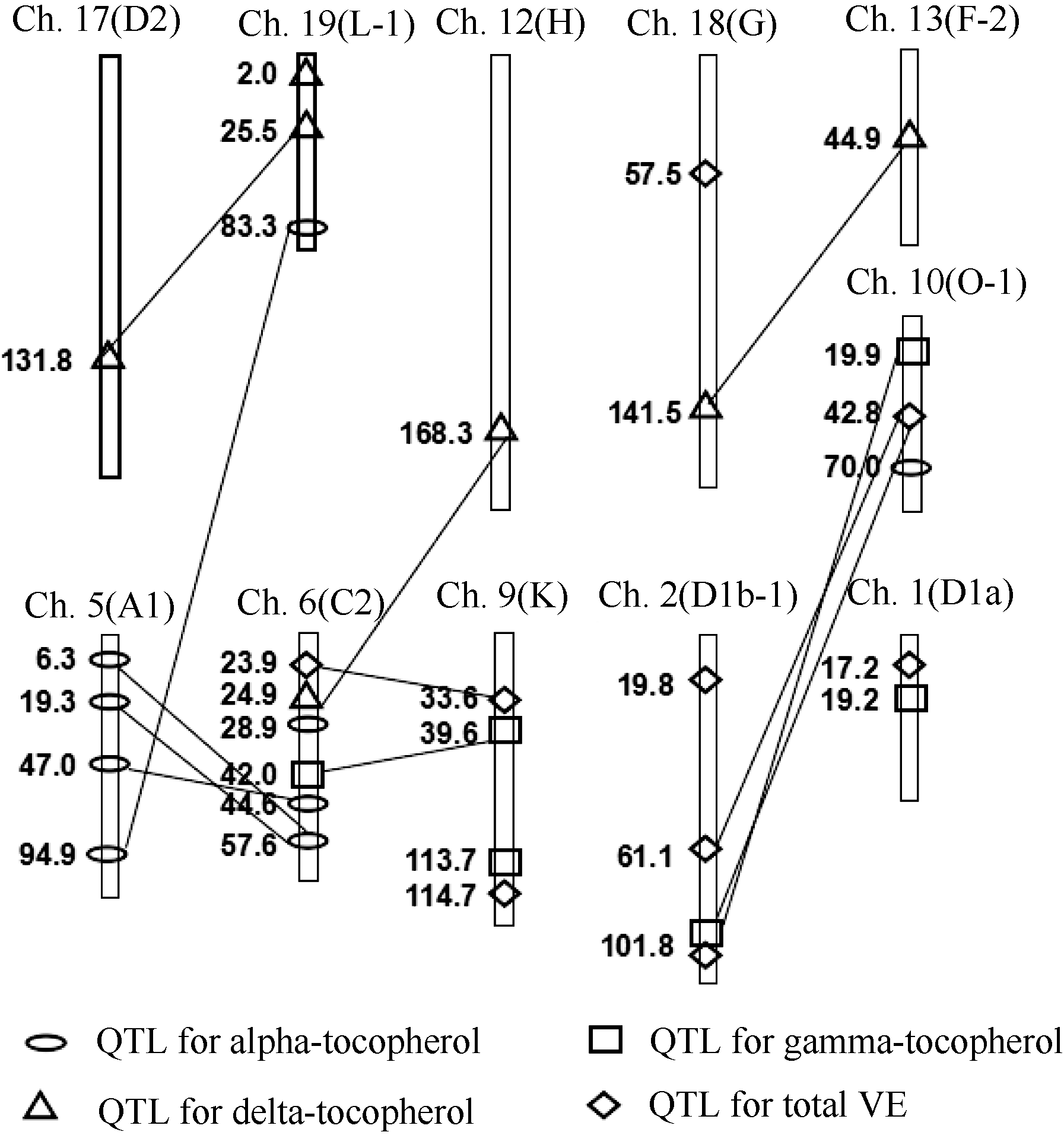
表4 BIEX群体中以 MCIM 法检测出的大豆籽粒维生素E含量的加性QTL Table 4 Additive QTL of vitamin E contents of soybean seed detected by MCIM in the BIEX population
性状 Trait
QTL
连锁群 Linkage group
染色体 Chr.
标记区间 Marker interval
位置 Position
置信区间 Confidence interval
加性QTL Additive QTL
Carto- graphera
IciMa- ppinga
加性效应a
加性遗传率h2(a) (%)
α -Toc
qα 3-1
O-1
Gm10
Sat_318-Sat_196
70.0
61.3-79.0
-1.08
2.40
2
2, m
qα 17-1
C2
Gm6
Sat_402-Satt277
28.9
25.9-33.5
1.78
6.18
1, 2, m
1, 2, m
γ -Toc
qγ 20-1
K
Gm9
Satt617-Satt001
113.7
105.5-122.7
-6.55
3.64
1, 2, m
qγ 24-1
D1a
Gm1
Sat_346-Satt436
19.2
8.2-29.2
-9.04
4.34
2, m
δ -Toc
qδ 14-1
L-1
Gm19
Sat_320-Satt313
2.0
0.0-12.0
-1.58
5.49
1, 2, m
1, 2, m
TVE
qTVE10-1
G
Gm18
Sat_164-Satt612
57.5
46.6-67.5
8.46
3.06
1, 2, m
qTVE20-1
K
Gm9
Satt617-Satt001
114.7
105.5-123.7
-7.04
2.80
2
qTVE24-1
D1a
Gm1
Sat_346-Satt436
17.2
8.2-25.2
-14.04
5.63
1, 2, m
1
Position: the distance between QTL and the first marker of the relevant linkage group; h2(a): phenotypic variation explained by the additive QTL. a The numbers 1, 2, and m indicate that the QTL was detected in 2008, 2009 and over two years, respectively. Other abbreviations are the same as those given in Table 1. 位置: QTL与相应连锁群第1个标记间的距离; h2(a): 加性QTL 解释的表型变异。a: 数字1、2和m说明该QTL能被2008、2009和两年平均检测出。其他缩写同表1。
表4 BIEX群体中以 MCIM 法检测出的大豆籽粒维生素E含量的加性QTL Table 4 Additive QTL of vitamin E contents of soybean seed detected by MCIM in the BIEX population
图1 以MCIM法检测出大豆维生素E的QTL在连锁图上的 位置椭圆: α -生育酚QTL; 方框: γ -生育酚QTL; 三角: δ -生育酚QTL; 菱形: VE含量QTL; 直线: 相连的2个加性QTL间有互作。Fig. 1 Locations of QTL for vitamin E contents detected by MCIM in soybean linkage mapOval: QTL for α -Toc; Square: QTL for γ -Toc; Triangle: QTL for δ -Toc; Rhombus: QTL for total vitamin E contents. Line: joining two additive QTLs represent epistatic interactions between them.
表5 BIEX群体中以MCIM法检测出的大豆籽粒维生素E含量的互作QTL对 Table 5 Epistatic QTL pairs of VE contents of soybean seed detected by MCIM in the BIEX population
性状 Trait
互作QTL对 QTL pair
QTL
连锁群 Linkage group
标记区间 Marker region
位置 Position
上位效应 Epistatic effects
贡献率 h2(aa) (%)
α -Toc
α -Toc-e1
α -Toc-e1, 1
L-1
Satt143-Sat_301
83.3
-1.36
5.29
α -Toc-e1, 2
A1
Sat_384-Sat_385
94.9
α -Toc-e2
α -Toc-e2, 1
A1
Satt225-Satt200
6.3
-1.07
3.67
α -Toc-e2, 2
C2
Sat_142-Satt658
57.6
α -Toc-e3
α -Toc-e3, 1
A1
Satt200-Satt174
19.3
-0.81
4.12
α -Toc-e3, 2
C2
Sat_142-Satt658
57.6
α -Toc-e4
α -Toc-e4, 1
A1
Satt619-Satt385
47.0
1.57
3.03
α -Toc-e4, 2
C2
Sat_142-Satt658
44.6
γ -Toc
γ -Toc-e1
γ -Toc-e1, 1
O-1
Satt633-Satt259
19.9
12.26
7.79
γ -Toc-e1, 2
D1b-1
AI856415-Satt634
101.8
γ -Toc-e2
γ -Toc-e2, 1
C2
Satt489-Sat_142
42.0
-13.83
4.35
γ -Toc-e2, 2
K
Satt260-Sat_399
39.6
δ -Toc
δ -Toc-e1
δ -Toc-e1, 1
D2
Satt301-Satt186
131.8
-2.20
6.19
δ -Toc-e1, 2
L-1
Sat_320-Satt313
25.0
δ -Toc-e2
δ -Toc-e2, 1
F-2
Sat_262-Satt343
44.9
1.77
4.85
δ -Toc-e2, 2
G
Satt610-Sat_141
141.5
δ -Toc-e3
δ -Toc-e3, 1
H
Satt353-Sat_214
168.3
-2.03
7.20
δ -Toc-e3, 2
C2
Sat_402-Satt277
24.9
TVE
TVE-e1
TVE-e1, 1
O-1
Satt653-BF008905
42.8
9.00
1.34
TVE-e1, 2
D1b-1
AI856415-Satt634
101.8
TVE-e2
TVE-e2, 1
O-1
Satt653-BF008905
42.8
-8.77
2.32
TVE-e2, 2
D1b-1
Satt290-Satt266
61.1
TVE-e3
TVE-e3, 1
C2
Sat_402-Satt277
23.9
-15.40
4.36
TVE-e3, 2
K
Satt260-Sat_399
33.6
This epistatic effect of parental two-locus genotypes is positive while that of non-parental two-locus genotypes (recombinants) is negative. Abbreviations are the same as those given in Table 1. 上位性效应正值表示亲本型有正向效应, 而重组型有负向效应; 负值则相反。缩写同表1。
表5 BIEX群体中以MCIM法检测出的大豆籽粒维生素E含量的互作QTL对 Table 5 Epistatic QTL pairs of VE contents of soybean seed detected by MCIM in the BIEX population
Hincha DK. Effects of α-tocopherol (vitamin E) on the stability and lipid dynamics of model membranes mimicking the lipid composition of plant chloroplast membranes. FEBS Lett, 2008, 582: 3687-3692[本文引用:1][JCR: 3.582]
[3]
Abbasi AR, HajirezaeiM, HofiusD, SonnewaldU, Voll LM. Specific roles of α- and γ-tocopherol in abiotic stress responses of transgenic tobacco. Plant Physiol, 2007, 143: 1720-1738[本文引用:1][JCR: 6.555]
[4]
Prasad KN, KumarA, KochupillaiV, Cole WC. High doses of multiple antioxidant vitamins: essential ingredients in improving the efficiency of stand ard cancer therapy. J Am Coll Nutr, 1999, 18: 13-25[本文引用:1][JCR: 1.738]
[5]
Bramley PM, ElmadfaI, KafatosA, Kelly FJ, ManiosY, Roxborough HE, SchuchW, SheehyP J A, WagnerK H, 2000: Vitamin E. J Sci Food Agric, 2000, 80: 913-938[本文引用:1]
[6]
KanwischerM, PorfirovaS, BergmullerE, DörmannP. Alterations in tocopherol cyclase activity in transgenic and mutant plants of Arabidopsis affect tocopherol content, tocopherol composition, and oxidative stress. Plant Physiol, 2005, 137: 713-723[本文引用:1][JCR: 6.555]
[7]
米歇尔·琼斯, 田颖. 天然维生素E. 日用化学品科学, 2002, (4): 47-48MichelleJ, TianY. Naturals vitamin E. Deterg Cosmet, 2002, (4): 47-48 (in Chinese with English abstract)[本文引用:1][CJCR: 0.3009]
[8]
UjiieA, YamadaT, FujimotoK, EndoY, KitamuraK. Identification of soybean varieties with high α-tocopherol content. Breed Sci, 2005, 55: 123-125[本文引用:1][JCR: 1.04]
[9]
李卫东, 卢为国, 梁慧珍, 王树峰, 苑保军, 耿臻, 王素阁, 范彦英, 刘亚非, 王令涛. 大豆籽粒维生素E含量与生态因子关系的研究. 作物学报, 2007, 33: 1094-1099Li WD, Lu WG, Liang HZ, Wang SF, Yuan BJ, GengZ, Wang SG, Fan YY, Liu YF, Wang LT. Effects of eco-physiological factors on vitamin E content in soybean seed. Acta Agron Sin, 2007, 33: 1094-1099 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[10]
Dwiyanti MS, UjiieA, ThuyL T B, YamdaT, KitamuraK. Genetic analysis of high α-tocopherol content in soybean seeds. Breed Sci, 2007, 57: 23-28[本文引用:2][JCR: 1.04]
[11]
Li HY, Liu HC, Han YP, Wu XX, Teng WL, Liu GF, Li WB. Identification of QTL underlying vitamin E contents in soybean seed among multiple environments. Theor Appl Genet, 2010, 120: 1405-1413[本文引用:4][JCR: 3.658]
[12]
Song QJ, Marek LF, Shoemaker RC, Lark KG, Concibido VC, DelannayX, Specht JE, Cregan PB. A new integrated genetic linkage map of the soybean. Theor Appl Genet, 2004, 109: 122-128[本文引用:1][JCR: 3.658]
[13]
王宇峰. 大豆基因组SSR分布特征和高密度遗传图谱的构建、整合与应用. 南京农业大学博士学位论文, 江苏南京, 2009Wang YF. Genomic Characterization of Simple Sequence Repeats and Establishment, Integration and Application of High Density Genetic Linkage Map in Soybean. PhD Dissertation of Nanjing Agricultural University, Jiangsu, China, 2009 (in Chinese with English abstract)[本文引用:1]
[14]
YangJ, ZhuJ, Williams RW. Mapping the genetic architecture of complex traits in experimental populations. Bioinformatics, 2007, 23: 1527-1536[本文引用:1][JCR: 5.323]
[15]
Piepho HP, GauchH G J. Marker pair selection for mapping quantitative trait loci. Genetics, 2001, 157: 433-444[本文引用:1][JCR: 4.389]
Wang SC, Basten CJ, Zeng ZB. Windows QTL Cartographer 2. 5. North Carolina State University, Raleigh, NC, USA, 2001-2006[本文引用:1]
[18]
Li HH, Ye GY, Wang JK. A modified algorithm for the improvement of composite interval mapping. Genetics, 2007, 175: 361-374[本文引用:1][JCR: 4.389]
[19]
邢光南, 周斌, 赵团结, 喻德跃, 邢邯, 陈受宜, 盖钧镒. 大豆抗筛豆龟蝽Megacota cribraria (Fabricius)的QTL分析. 作物学报, 2008, 34: 361-368Xing GN, ZhouB, Zhao TJ, Yu DY, XingH. ChenS Y, Gai JY. Mapping QTLs of resistance to Megacota cribraria (Fabricius) in soybean. Acta Agron Sin, 2008, 34: 361-368 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[20]
ChantretN, MingeotD, SourdilleP, BernardM, Jacquemin JM, DoussinaultG. A major QTL for powdery mildew resistance is stable over time and at two development stages in winter wheat. Theor Appl Genet, 2001, 103: 962-971[本文引用:1][JCR: 3.658]
[21]
Symonds VV, Godoy AV, AlconadaT, Botto JF, Juenger TE, Casal JJ, Lloyd AM. Mapping quantitative trait loci in multiple populations of Arabidopsis thaliana identifies natural allelic variation for trichome density. Genetics, 2005, 169: 1649-1658[本文引用:1][JCR: 4.389]
[22]
Tucker DM, Griffey CA, LiuS, Brown-GuediraG, Marshall DS, Saghai MaroofM A. Confirmation of three quantitative trait loci conferring adult plant resistance to powdery mildew in two winter wheat populations. Euphytica, 2007, 155: 1-13[本文引用:1][JCR: 1.643]
[23]
李广军, 李河南, 程利国, 章元明. 大豆叶绿素含量动态表达的QTL分析. 作物学报, 2010, 36: 242-248Li GJ, Li HN, Cheng LG, Zhang YM. QTL analysis for dynamic expression of chlorophyll content in soybean. Acta Agron Sin, 2010, 36: 242-248[本文引用:1][CJCR: 1.667]
[24]
Rossi ME, Orf JH, Liu LJ, DongZ, RajcanI. Genetic basis of soybean adaptation to North American vs. Asian mega- environments in two independent populations from Canadian × Chinese crosses. Theor Appl Genet, 2013, 126: 1809-1823[本文引用:1][JCR: 3.658]
[25]
PalomequeL, Li JL, LiW, HedgesB, Cober ER, RajcanI. QTL in mega-environments: II. Agronomic trait QTL co-localized with seed yield QTL detected in a population derived from a cross of high-yielding adapted x high-yielding exotic soybean lines. Theor Appl Genet, 2009, 119: 429-436[本文引用:1][JCR: 3.658]
[26]
HanY, LiD, ZhuD, LiH, LiX, TengW, LiW. QTL analysis of soybean seed weight across multi-genetic backgrounds and environments. Theor Appl Genet, 2012, 125: 671-683[本文引用:1][JCR: 3.658]
[27]
Fasoula, VasiliaA, Harris, DonnaK, BoermaH R. Validation and designation of quantitative trait loci for seed protein, seed oil, and seed weight from two soybean populations. Crop Sci, 2004, 44: 1218-1225[本文引用:1][JCR: 1.513]
[28]
KorirP, QiB, WangY, ZhaoT, YuD, ChenS, GaiJ. A study on relative importance of additive, epistasis and unmapped QTL for aluminum tolerance at seedling stage in soybean. Plant Breed, 2011, 130: 551-562[本文引用:1][JCR: 1.175]
[29]
Della PD. Progress in the dissection and manipulation of vitamin E synthesis. Trends Plant Sci, 2005, 10: 574-579[本文引用:1][JCR: 11.808]
[30]
ShintaniD, Della PD. Elevating the vitamin E content of plants through metabolic engineering. Science, 1998, 282: 2098-2100[本文引用:1]
[31]
Venkatesh TV, Karunanand aaB, Free DL, Rottnek JM, Baszis SR, Valentin HE. Identification and characterization of an Arabidopsis homogentisate phytyltransferase paralog. Planta, 2006, 223: 1134-1144[本文引用:1][JCR: 3.347]
[32]
李灵娟. 甘蓝型油菜基因组中控制种子维生素E含量QTL的检测和分析. 华中农业大学硕士学位论文, 湖北武汉, 2008Li LJ. Identification and Analysis of QTL Controlling Seed Tocopherol Content in the Genome of Brassica napus L. MS Thesis of Huazhong Agricultural University, Hubei, China, 2008 (in Chinese with English abstract)[本文引用:1]
 , 李海朝
, 李海朝



{kind=link}
{kind=link}