 , 刘天慧1,2, 任玉芬2
, 刘天慧1,2, 任玉芬2 , 张红星2, 苏芝敏2, 王思琪2, 王效科2, 方文颖3
, 张红星2, 苏芝敏2, 王思琪2, 王效科2, 方文颖31. 河南理工大学资源环境学院, 焦作 454000;
2. 中国科学院生态环境研究中心城市与区域生态国家重点实验室, 北京城市生态系统研究站, 北京 100085;
3. 北京市南水北调大宁管理处, 北京 102442
收稿日期: 2019-11-19; 修回日期: 2020-01-10; 录用日期: 2020-01-10
基金项目: 国家重点研发计划项目(No.2016YFC0503004, 2017YFF0207303);国家自然科学基金(No.71533005, 31600333)
作者简介: 贺玉晓(1976-), 女, E-mail:heyuxiao@hpu.edu.cn
通讯作者(责任作者): 任玉芬, E-mail:yfren@rcees.ac.cn
摘要:浮游植物是水生生态系统的主要生产者,其群落结构与水质密切相关.为研究北京市北运河不同水体环境特征,于2018年10月(秋季)和2019年2月(冬季)在北京市北运河设置13个采样点开展浮游植物群落结构和水环境调查,共鉴定出浮游植物7门54属99种.群落结构分析表明,秋季浮游植物物种数(75种)高于冬季(58种),城市河道型水体物种数(78种)>城市湖泊型(59种)>山区河道型(29种).秋季浮游植物密度为916.04×104 cells·L-1,冬季浮游植物密度为220.52×104 cells·L-1,秋、冬季平均密度为568.28×104 cells·L-1.空间格局上,城市湖泊型水体浮游植物密度最高,绿藻门和蓝藻门为优势门类;城市河道型水体次之,大部分点位以硅藻门、绿藻门和蓝藻门为优势门类;山区河道型水体最低,硅藻门为主要优势门类.物种优势度和多样性指数分析结果显示,北京市北运河水体中以中富营养型指示物种为优势类群,水质污染类型为β-中污型.典范对应分析(CCA)排序结果表明,Ca和TN是影响秋季北京市北运河浮游植物群落结构的主要环境因子,pH是影响冬季浮游植物群落结构的主要环境因子.
关键词:浮游植物水环境污染群落结构环境因子北运河
Characteristics and influencing factors of phytoplankton community structure in autumn and winter of the North Canal, Beijing
HE Yuxiao1
, LIU Tianhui1,2, REN Yufen2, ZHANG Hongxing2, SU Zhimin2, WANG Siqi2, WANG Xiaoke2, FANG Wenying31. Institute of Resources and Environment, Henan Polytechnic University, Jiaozuo 454000;
2. Beijing Urban Ecosystem Research Station, State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085;
3. Daning Management Office of the Beijing South-to-North Water Diversion, Beijing 102442
Received 19 November 2019; received in revised from 10 January 2020; accepted 10 January 2020
Abstract: Phytoplankton is a major producer of water ecosystems, and its community structure is closely related to water quality. This study selected 13 sites samples in the North Canal to investigate the phytoplankton community and water environment in October 2018(autumn) and February 2019(winter). A total of 99 phytoplankton species were identified, which belonged to 7 phyla and 54 genera. Community structure analysis showed that the species composition of phytoplankton in autumn(75 species) was higher than that in winter(58 species). The species composition of phytoplankton in different waters showed urban river(78 species) > urban lake(59 species) > mountain river(29 species). Phytoplankton density decreased from 916.04×104 cells·L-1 in autumn to 220.52×104 cells·L-1 in winter, with the average density of 568.28×104 cells·L-1. Phytoplankton in urban lake had the highest cell abundance, dominated by Chlorophyta and Cyanophyta; urban river had the second highest abundance, dominated by Bacillariophyta, Cyanophyta and Chlorophyta in the most sites; mountain rivers had the lowest phytoplankton abundance, dominated by Bacillariophyta. The analysis of species dominance and diversity index showed that the North Canal was in the β-medium pollution level with the eutrophic indicator alga as the dominant group. A canonical correspondence analysis(CCA) showed that the main environmental factors affecting the phytoplankton community structure in the North Canal were calcium(Ca) and total nitrogen(TN) in autumn and potential of hydrogen(pH) in winter.
Keywords: phytoplanktonwater environmental pollutioncommunity structureenvironmental factorsNorth Canal
1 引言(Introduction)浮游植物作为河流和湖泊生态系统的重要组成部分, 通过光合作用合成有机物质, 构成了水生态环境中食物链的重要环节(陈倩等, 2018;Zheng et al., 2019).浮游植物对水环境变化极为敏感, 较其它物种更能及时反映水生态环境变化, 其群落结构组成、分布特征可客观反映水环境变化规律, 是评价水体状况的重要指示物种之一(Wu et al., 2019;朱忆秋等, 2019).目前, 浮游植物群落组成与多样性分析已广泛应用于各类水域的水环境评价, 成为水域生态学研究的重要内容之一(Sellami et al., 2016;Nunes et al., 2018;贾慧娟等, 2019;Kraus et al., 2019).例如, 宋勇军等(2019)通过综合营养状态指数法和藻类生物学法分析得出程海湖已达到轻度富营养水平.君珊等(2019)对拉萨河浮游植物群落组成、丰度、功能群及多样性指数进行了调查, 利用典范对应分析法(CCA)分析发现, pH、化学需氧量(CODCr)、总磷(TP)是影响浮游植物群落结构的主要环境因子.Lee等(2015)通过对温带浅层湖泊中浮游植物物种与水质的响应, 分析浮游植物群落的动态特征, 从而有效预测和管理蓝藻水华.
北运河流域是北京市人口最集中、产业最聚集、城市化水平最高的地区, 北京市境内的北运河流域面积共4348 km2, 境内的河长为89.4 km(Shen et al., 2014), 担负着城中心四区及部分郊区雨洪及废污水排泄任务, 是北京市受污染程度较高的流域, 水资源短缺、水生态系统退化严重(马思琦等, 2019;张家铭等, 2019).目前有关北京市北运河浮游植物群落特征及其影响因子的研究相对较少, 对浮游植物群落的研究大多集中于城区部分河段或者单个流域(王晓凤等, 2011;高彩凤等, 2012).本研究将北运河划分为山区河道型、城市河道型和城市湖泊型3种类型, 于2018年10月和2019年2月开展秋、冬季不同类型水体浮游植物群落分布与组成差异调查.在浮游植物群落组成和水质数据测定的基础上, 运用典范对应分析(CCA)研究浮游植物和环境因子的关系, 分析不同类型水体中浮游植物群落结构特征及其影响因子, 为北京市北运河水质监测与评价、水质提升和水生态系统恢复提供基础数据和技术支撑.
2 材料与方法(Materials and methods)2.1 采样点设置采样点主要设置在北京市城市边界内, 沿着京密引水渠和北京市五环的河道分布.根据其水体类型的多样性和生境特点, 将北运河按山区河道型、城市河道型和城市湖泊型划分为3个采集区域, 共设置13个采样点位(图 1、表 1).其中, 锥石口沟上游(S1)和锥石口沟下游(S2)为山区河道型水体, 朝阳公园南湖(S3)和陶然亭湖(S4)为城市湖泊型水体, 是北京中心城市景观湖泊, 其余的采样点为城市河道型水体, 是北京市中心城区的主要雨污水排水河道, 主要包括清河支流(S5)、清河上游(S6)、京密引水渠(S7)、南长河(S8)、莲花河(S9)、西坝河(S10)、通惠河中游(S11)、通惠河上游(S12)及凉风灌渠(S13).采样时间为2018年10月22—23日和2019年2月22—23日, 均在上午9:00—12:00完成采样.每次采样安排在连续晴天、水位较稳定的日子.
图 1(Fig. 1)
 |
| 图 1 北运河采样点分布 Fig. 1Locations of the sampling sites in the North Canal |
表 1(Table 1)
| 表 1 采样点分布 Table 1 Distribution of sampling sites | ||||||||||||||||||||||||||||||||||||||||||||||
表 1 采样点分布 Table 1 Distribution of sampling sites
| ||||||||||||||||||||||||||||||||||||||||||||||
2.2 水体理化因子测定采用便携式YSI多参数水质检测仪现场测定水温(WT)、溶解氧(DO)、电导率(Cond)和pH值.在各采样点采集1 L水样, 置于保温箱中冷藏保存(4 ℃), 于48 h内带回实验室进行样品的水质指标测定.参照《水和废水分析方法》对水体中的总氮(TN)、叶绿素a(Chla)、总磷(TP)、化学需氧量(COD)、氨氮(NH4+-N)、总有机碳(TOC)、总无机碳(TIC)、总碳(TC)等进行测定(国家环境保护总局《水和废水分析方法》编委会, 2002).
2.3 浮游植物采集及检测方法浮游植物的采样过程严格按照《水生生物学》的相关采样方法进行(赵文, 2005).
2.3.1 定性样品采集浮游植物的定性样品用浮游生物网对各采样点的表层水体进行采集, 在水面呈“∞”状拖动浮游网数次, 最后浓缩到50 mL试剂瓶中, 用1.5%的鲁哥氏液现场固定, 在每个样点釆集定性样品1个.浮游植物计数方法为目镜视野法, 在10×40倍的光学显微镜下参考相关文献(赵文, 2005;胡鸿钧, 2006;陈贺等, 2016;徐宗学等, 2017)进行鉴定和计数.
2.3.2 定量样品采集浮游植物的定量样品用有机玻璃采水器采集1 L表层水样, 加入1.5%的鲁哥氏液固定, 实验室静置沉淀48 h后, 对上层水进行虹吸, 浓缩至100 mL.定量计数前将沉淀样品充分摇匀, 然后吸取0.1 mL样品注入0.1 mL的技术框内, 每个样品至少计数2片, 每片计数50~100个视野, 同一个样品的两次计数结果与平均值之差不大于±15%, 即视为有效结果, 取其平均值.
2.4 数据处理与分析根据浮游植物现存量评价水体营养类型, 评价标准见表 2(孙林丹等, 2019).采用Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J)评估水体营养状态, 采用优势度指数(Y)(李建茹等, 2017)描述浮游植物群落优势种, 具体计算公式如下所示(杨俊丽等, 2019):
表 2(Table 2)
| 表 2 营养类型评价指标与标准 Table 2 Eutrophic level evaluation indicator and standards | ||||||||||
表 2 营养类型评价指标与标准 Table 2 Eutrophic level evaluation indicator and standards
| ||||||||||
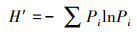 | (1) |
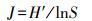 | (2) |
 | (3) |
表 3(Table 3)
| 表 3 多样性指数的评价标准 Table 3 Assessment standard of diversity indexes | ||||||||||||||||||||
表 3 多样性指数的评价标准 Table 3 Assessment standard of diversity indexes
| ||||||||||||||||||||
数据绘图使用Origin软件包;对多样性指数与环境因子用SPSS 22.0软件包进行Pearson相关分析;用单因素方差(ANOVA)检验秋、冬季不同水体间水质指标及浮游植物多样性的差异性;采用Canoco 5.0软件包对浮游植物优势种作去趋势对应分析(Detrended correspondence analysis, DCA), 若最大梯度长度大于4, 则对物种数据和相关环境因子数据进行CCA分析(安睿等, 2017), 获得浮游植物优势物种与环境因子的排序图.为消除极值影响, 在进行统计分析之前对数据进行log(x+1)转换, 使数据满足正态分布(邱阳凌等, 2018).
3 结果与分析(Results and discussion)3.1 浮游植物种类组成与优势种调查共获得浮游植物99种(包含7个变种), 隶属于7门54属.其中, 硅藻门(Bacillariophyta)和绿藻门(Chlorophyta)均为21属, 蓝藻门(Cyanophyta)7属, 裸藻门(Euglenophyta)3属, 甲藻门(Pyrrophata)、黄藻门(Xanthophyta)和金藻门(Chrysophyta)各1属.硅藻门种类最多, 有41种, 占物种总数的41.41%, 其次为绿藻门35种, 占35.35%, 此二者种类总和占本次调查总数的76.76%.此外, 检测出的蓝藻门10种, 占10.10%;裸藻门8种, 占8.08%.
北京市北运河山区河道型、城市河道型和城市湖泊型水体的浮游植物物种数存在时空差异(图 2).季节上, 秋季检测到的浮游植物种类数(75种)多于冬季(58种).空间上, 城市湖泊型水体平均物种数最多, 为21种;其次为城市河道型水体, 为14种;山区河道型水体最少, 为13种.
图 2(Fig. 2)
 |
| 图 2 浮游植物物种丰度的空间分布特征 Fig. 2Spatial distribution characteristics of phytoplankton species richness |
北京市北运河秋、冬季各采样点的浮游植物优势种(Y≥0.02)统计结果如表 4所示.在山区河道型水体中浮游植物优势种以硅藻门为主, 而城市湖泊型水体和城市河道型水体均以硅藻、绿藻和蓝藻门为主.其中, 作为中富营养型指示生物(α-β)硅藻门的针杆藻(Synedra sp., 中富营养型指示生物)在山区河道型水体、城市湖泊型水体和城市河道型水体均有发现;绿藻门的四尾栅藻(Scenedesmus quadricauda, 中营养型指示生物)在城市湖泊型水体和城市河道型水体中出现;硅藻门的谷皮菱形藻(Nitzschia palea, 中富营养型指示生物)在城市河道型水体中出现;其余藻类如绿藻门的丝藻(Ulothrix sp., 中富营养型指示生物)等在清河支流(S5)中出现.
表 4(Table 4)
| 表 4 各采样断面浮游植物优势种 Table 4 Dominant phytoplankton species in the sampling sites | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
表 4 各采样断面浮游植物优势种 Table 4 Dominant phytoplankton species in the sampling sites
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
3.2 浮游植物细胞密度调查期间, 秋、冬季各采样点的浮游植物密度存在差异, 变化范围为(38.33~4558.42)×104 cells·L-1.其中, 秋季浮游植物平均密度(916.04×104 cells·L-1)高于冬季(220.52×104 cells·L-1).从空间上看, 浮游植物密度分布也有明显差异, 山区河道型水体(S1~S2)的浮游植物物种以硅藻门为主, 浮游植物密度变化范围为(58.33~411.67)×104 cells·L-1, 均值为222.92×104 cells·L-1;城市湖泊型水体(S3~S4)物种主要包括蓝藻门和绿藻门, 其密度变化范围为(141.67~4558.42)×104 cells·L-1, 均值为1400.86×104 cells·L-1;城市河道型水体(S5~S13)以硅藻门、蓝藻门和绿藻门为主, 浮游植物密度变化范围为(38.33~2735.05)×104 cells·L-1, 均值为460.01×104 cells·L-1.结合营养类型评价标准(表 3)可知, 山区河道型水体和城区河道型水体均为中营养型, 而城市湖泊型水体为富营养型.
3.3 水体水质状况由表 5可知, 调查期间, 秋、冬季山区河道型水体水温明显低于城市河道型和城市湖泊型水体(p < 0.01);各采样点的pH冬季高于秋季(p < 0.01), 水体呈弱碱性.总氮(TN)、总磷(TP)和其他环境因子在调查期间季节性差异不明显(p>0.05).大部分采样点的总磷(TP)含量差别不大, 均符合Ⅲ类地表水标准, 而陶然亭湖(S4)和通惠河中游(S11)的总磷(TP)含量超标严重, 分别为1.33 mg·L-1和25.83 mg·L-1, 高于V类水质标准(0.4 mg·L-1), 属于劣V类水质.京密引水渠(S7)和南长河(S8)的总氮(TN)含量符合IV类水质标准, 而其它水体均高于V类水质标准(2 mg·L-1).其中, 陶然亭湖(S4)和清河上游(S6)、莲花河(S9)的总氮(TN)含量超V类水国家标准6倍.
图 3(Fig. 3)
 |
| 图 3 各采样点浮游植物密度时空分布 Fig. 3Temporal and spatial distribution of phytoplankton abundance at each sampling sites |
表 5(Table 5)
| 表 5 各采样点主要水化学参数(平均值±标准差) Table 5 Main water chemical parameters of sampling sites(Mean±SD) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
表 5 各采样点主要水化学参数(平均值±标准差) Table 5 Main water chemical parameters of sampling sites(Mean±SD)
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
3.4 浮游植物群落多样性指数单因素方差分析(ANOVA)结果表明, 北京市北运河浮游植物的Shannon-Wiener多样性指数(H′)和Pielou均匀度指数(J)秋、冬季节变化均不明显, 不同监测点位间的多样性差异性不明显(p>0.05).北运河山区河道型、城市河道型和城市湖泊型水体的环境因子与Shannon-Wiener多样性指数(H′)和Pielou均匀度指数(J)的Pearson相关性分析如表 6所示.结果显示, Pielou均匀度指数(J)与TC呈负相关关系(p < 0.05);Shannon-Wiener多样性指数(H′)与叶绿素a(Chla)和Pielou均匀度指数(J)均呈正相关关系(p < 0.05), 说明北运河的Shannon-Wiener多样性指数(H′)的大小是由叶绿素a(Chla)和Pielou均匀度指数(J)共同决定.季节上, 北运河的Shannon-Wiener多样性指数(H′)和Pielou均匀度指数(J)秋季高于冬季(图 4).北京市北运河浮游植物的Shannon-Wiener多样性指数(H′)为1.35~2.67, 属β-中污型, 其中, 山区河道型水体的变化范围为2.00~2.61, 均值为2.31;城市湖泊型水体的变化范围为1.51~2.67, 均值为2.10;城市河道型水体的变化范围为1.35~2.51, 均值为2.01.Pielou均匀度指数(J)为0.49~0.94, 水质评价结果为轻污型至β-中污型, 其中, 山区河道型水体的变化范围为0.83~0.94, 均值为0.90;城市湖泊型水体的变化范围为0.61~0.86, 均值为0.72;城市河道型水体的变化范围为0.49~0.91, 均值为0.80.综合Shannon-Wiener多样性指数(H′)和Pielou均匀度指数(J)可知, 山区河道型水体水质优于城市湖泊型水体和城市河道型水体.
表 6(Table 6)
| 表 6 多样性指数与环境因子的Pearson相关性分析(n=26) Table 6 Pearson correlation analysis of environmental factors and diversity index | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
表 6 多样性指数与环境因子的Pearson相关性分析(n=26) Table 6 Pearson correlation analysis of environmental factors and diversity index
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
图 4(Fig. 4)
 |
| 图 4 各采样点浮游植物Shannon-Wiener多样性指数(H′)和Pielou均匀度指数(J) Fig. 4Diversity index and evenness index of the phytoplankton at different sampling sites |
3.5 浮游植物群落结构与环境因子的CCA分析对北京市北运河秋、冬季浮游植物优势种数据分别进行去趋势对应分析(DCA), 发现最大梯度长度(Lengths of gradient)均大于4, 故采用典范对应分析法(CCA)对秋、冬季环境因子与浮游植物优势种群落进行分析.利用向前引入法(Forward selection)分别从温度、pH、溶解氧、叶绿素、COD、TN、TP、NH4+-N、TOC、TIC和TC等筛选出具有显著解释性的环境变量与采样点进行CCA分析.结果表明(表 7), 轴1的特征值及其浮游植物与环境的相关性、种类物种变量、物种与环境关系的相关性均大于轴2.前两轴的物种-环境关系累积百分比分别为60.57%和65.03%, 表明排序能够较好地反映浮游植物与环境因子间的相互关系.运用偏冗余分析(Variation partitioning analysis)对秋、冬季各个环境显著因子进行分析, 探讨主要环境因子对浮游植物分布的影响, 结果见表 8.其中, 环境因子中的Ca(F=1.8, p=0.004)、TN(F=1.5, p=0.05)和pH(F=1.6, p=0.032)对各采样点浮游植物分布的影响达到显著水平, 而其他环境变量对浮游植物的影响没有达到显著水平(p>0.05), 说明Ca和TN是影响秋季浮游植物群落结构的主要环境因子, 而pH是影响冬季浮游植物群落结构的主要环境因子.
表 7(Table 7)
| 表 7 浮游植物优势种CCA分析结果 Table 7 CCA analysis results of dominant phytoplankton species | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
表 7 浮游植物优势种CCA分析结果 Table 7 CCA analysis results of dominant phytoplankton species
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
表 8(Table 8)
| 表 8 浮游植物与显著环境因子的偏冗余分析 Table 8 Variation partitioning analysis of the significant environmental factors on the phytoplankton | |||||||||||||||||||
表 8 浮游植物与显著环境因子的偏冗余分析 Table 8 Variation partitioning analysis of the significant environmental factors on the phytoplankton
| |||||||||||||||||||
秋、冬季北运河浮游植物分布特征PCA排序如图 5所示.由图 5可知, 各采样点水环境特点存在明显差异.在秋季, COD是影响山区河道型水体(S1~S2)浮游植物分布的主要环境因子, pH是影响城市湖泊型水体(S3~S4)的主要环境因子, 而城市河道型水体(S5~S13)主要受Ca和氮、磷营养盐的影响;而对于冬季, pH是影响山区河道型水体(S1~S2)、城市湖泊型水体(S3~S4)和城市河道型水体(S5~S13)浮游植物分布的主要环境因子.
图 5(Fig. 5)
 |
| 图 5 采样点与环境因子的CCA排序 Fig. 5CCA diagram of the sites and variables environmental |
北京市北运河浮游植物与环境因子的关系可从CCA排序图中看出(图 6):对于秋季, 梅尼小环藻(Cyclotella meneghiniana)、钝脆杆藻(Fragilaria capucina)、舟形藻(Navicula sp.)、谷皮菱形藻、肘状针杆藻(Synedra ulna)、具星小环藻(Cyclotella stelligera)等与TN、Ca和COD呈高度正相关, 与pH和TP呈负相关关系.对于冬季, 梅尼小环藻、近缘桥弯藻(Cymbella affinis)、舟形藻等与pH呈现高度正相关;优美平裂藻(Merismopedia elegans)、颤丝藻(Ulothrix oscillarina)、变异直链藻(Melosira varians)、尖针杆藻(Synedra acus)与pH、DO、TIC、TOC和WT呈正相关;扁圆卵形藻(Cocconeis placentula)、窄异极藻延长变种(Gomphonema angustatum var. productum)、放射舟形藻(Navicula radiosa)、四尾栅藻、披针曲壳藻(Achnanthes lanceolata)以及普通黄丝藻(Tribonema vulgare)与DO、TIC、TOC和WT呈高度正相关, 与pH呈负相关, 其中, 放射舟形藻对DO相关显著;而其他浮游植物与之相反.
图 6(Fig. 6)
 |
| 图 6 浮游植物群落与环境因子CCA排序图 Fig. 6CCA diagram of phytoplankton dominant species and environment factors |
4 讨论(Discussion)4.1 浮游植物物种组成与群落特征浮游植物物种组成和群落特征会受到生境和人类活动的双重影响(任辉等, 2017).本研究发现, 北京市北运河的浮游植物物种组成存在空间差异, 山区河道型水体浮游植物群落结构组成为硅藻型, 水体类型为中营养型, 物种组成主要包括硅藻门的普通等片藻(Diatoma vulgare)、变异直链藻、短线脆杆藻二凸变种(Fragilaria brecistriata)和缢缩异极藻(Gomphonema constrictum)等, 原因在于山区水体流速较快, 更适宜硅藻生长(胡忠军等, 2017).城市湖泊型水体浮游植物群落结构组成为蓝藻-绿藻型, 水体类型为重富营养型, 物种组成以蓝藻门的小席藻(Phormidium tenus)和平裂藻属(Merismopedia sp.), 绿藻门的单角盘星藻具孔变种(Pediastrum simplex var. duodenarium)、四尾栅藻为主, 原因在于湖泊水面宽阔, 透明度低, 水体交换能力差, 水体处于富营养化, 有利于蓝藻、绿藻的一些耐污种大量繁殖(严广寒等, 2019).而城市河道型水体以硅藻门的普通等片藻、变异直链藻、尖针杆藻, 绿藻门的栅藻属(Scenedesmus sp.)为主, 其群落结构组成为绿藻-硅藻型, 表明其水体富营养化程度较高, 城市河道水体大多流经市区, 城市河流污染较为严重, 造成绿藻等一些耐污种大量繁殖(孙祥等, 2017;曹文钟等, 2019).
浮游植物的细胞密度和污染指示物种是评价水体状况的关键指标(刘绍俊等, 2019).此次调查出现的优势种群大部分为污染指示种, 代表性物种如硅藻门的梅尼小环藻和绿藻门的四尾栅藻等, 均为富营养型指示种;除此之外, 裸藻属(Euglena sp.)、囊裸藻(Trachelomonas sp.)、栅藻、舟形藻、辐节藻(Staurneis sp.)也大都属污染指示种(李晶, 2012;陆欣鑫等, 2014).结合污染指示物种和藻类密度评价标准发现, 北京市北运河大部分水体处于富营养状态.
4.2 浮游植物群落的多样性特征在浮游植物生物多样性评价中, 多样性指数和均匀度指数越高, 水生态系统越稳定, 通常以Shannon-Wiener多样性指数(H′)小于1.0和Pielou均匀度指数(J)小于0.30作为群落多样性较差的标准(杨俊丽等, 2019).北运河山区河道型水体的Shannon-Wiener多样性指数(H′)和Pielou均匀度指数(J)较高, 其原因是山区相对偏僻, 人类活动干扰相对较少, 周围植被覆盖度高, 环境较好, 水体水草较多、流动缓慢, 利于不同浮游植物生长.城市河道型水体的多样性指数及均匀度指数均较低, 该结果与水体理化指标的监测情况相符合.城市河道型水体流速较快, 且有大量的营养盐及有机物输入, 对水体水质产生了剧烈影响(吴天浩等, 2019).
根据多样性指数评价法的水质评价结果可知, 研究区域的S1和S2采样点, 集水区内乔木生长茂盛, 植被覆盖度高, 水体中沉水植物和挺水植物分布广泛, 与其他采样点相比, 水体水质较好;而S3、S4采样点较S1和S2采样点透明度低, 水质较差;S5~S13站点位于市区, 生活污水、城市面源污染一定程度地破坏水体的生态功能.
4.3 浮游植物群落与环境因子的关系浮游植物能对水环境变化做出响应, pH、水温、营养盐、金属元素及浮游动物牧食等通常是影响浮游植物生长的环境因子(Zhou et al., 2010;李磊等, 2015;Han et al., 2016;Pramanik et al., 2016).本研究中, 浮游植物优势种与环境因子之间的典范对应分析结果表明, Ca和TN是影响秋季北运河浮游植物分布的主要环境因子, 而pH是影响冬季浮游植物分布的主要环境因子.
水体中的氮、磷既是浮游植物重要的营养元素, 也是水环境质量的重要指标之一(Turner et al., 2013).典范对应分析发现, 浮游植物中的针杆藻(Synedra sp., 中富营养型指示生物)、谷皮菱形藻(中富营养型指示生物)、栅藻、舟形藻等污染指示种与TP、TN呈正相关, 表明N、P等营养物会影响浮游植物的生长(Reed et al., 2016;Sitta et al., 2018).水质监测结果显示, 秋季TN平均浓度为6.50 mg·L-1, TP平均浓度为0.16 mg·L-1, TN/TP值平均为114;冬季TN、TP平均浓度分别为5.74 mg·L-1和2.04 mg·L-1, TN/TP值平均为293.依据Redfield定律, 磷可被考虑为北运河浮游植物生长的限制性环境因素(Redfield, 1958;Perkins et al., 2000;贾海燕等, 2019).因此, 控制水体中磷的含量及外源输入, 是预防北运河流域水体富营养化的主要途径之一.
水温是影响浮游植物光合作用、呼吸作用的重要因子, 通过影响细胞代谢活动相关酶的效率而影响浮游植物的生长(李红飞等, 2019).本次调查发现, 秋季浮游植物种类数和密度均高于冬季, 温度是造成城市湖泊型、城市河道型和山区河道型水体浮游植物群落组成及细胞密度秋、冬差异的主要原因.
一般认为, 水中的金属元素对藻类的繁殖和群体形成存在适宜浓度, 当该种金属元素浓度低于这一浓度阈值时, 金属元素浓度的增长会促进浮游植物形成群体, 反之则使浮游植物群体解聚死亡(赵荣芳, 2017).一些研究发现, 水体中的金属Ca是维持浮游植物正常生长的重要营养元素之一, 是藻类生长发育的重要调节因子, 能够维持细胞壁、细胞膜及膜结合蛋白的稳定性, 可以调节浮游植物的多种代谢过程, 适当增加Ca浓度可以提高浮游植物细胞凝集素的活力(李林等, 2012;闵海丽等, 2012;白宁静等, 2019).Ca还可降低其他重金属对硅藻的毒害作用, 促进浮游植物的生长(李文君, 2012).水体中钙离子浓度的降低将降低藻类的生长速率, 但过高浓度的钙离子也会抑制藻类的生长(Shi et al., 2013).另外, 不同门类的浮游植物对于水体中不同钙离子浓度的响应不同(陶志英等, 2017).通过典范对应分析(CCA)发现, 谷皮菱形藻、肘状针杆藻等与Ca浓度高度正相关, 其密度随Ca浓度的增加而增加, 说明Ca对浮游植物群落有重要影响, 这与前人的研究结果一致(李文君, 2012).
除营养盐、水温和金属元素外, pH也对浮游植物分布有重要影响(Kim et al., 2013), 浮游植物生长繁殖有其不同的pH范围(王为民, 2013), pH的高低会影响浮游植物群落结构的变化及分布, 反映出水体中藻类生长与水环境变化的状况, 碱性环境有利于浮游植物进行光合作用, 在碱性水体中浮游植物具有较高的藻类生产力(Jakobsen et al., 2015).通过CCA排序图发现, 桥弯藻、舟形藻、普通等片藻等对pH响应显著, 也说明碱性水体具有较高的藻类生产力.
5 结论(Conclusions)1) 北京市北运河浮游植物种类和密度在空间上呈现城市湖泊型水体>城市河道型水体>山区河道型水体, 时间上秋季浮游植物种类和密度显著高于冬季.
2) 秋、冬季节北京市北运河山区河道型水体主要优势种为硅藻门, 部分为中富营养型指示物种;城市河湖泊型和城市河道型水体主要优势种为蓝藻门和绿藻门的耐污种.综合Shannon-Wiener指数(H′)和Pielou均匀度指数(J)表明, 北京市北运河城市湖泊型、城市河道型和山区河道型水体均为β-中污型.
3) 北京市北运河城市湖泊型、城市河道型和山区河道型水体的浮游植物分布受环境影响较为显著.其中, Ca和TN是影响秋季浮游植物分布的主要环境因子, 而pH是影响冬季浮游植物群落分布的主要环境因子.
参考文献
| 安睿, 王凤友, 于洪贤, 等. 2017. 三环泡湿地浮游动物功能群季节变化及其影响因子[J]. 生态学报, 37(6): 1851-1860. |
| 白宁静, 陈丽, 蒋伊能, 等. 2019. 阳宗海砷污染背景下浮游植物的时空分布特征及其驱动因子解析[J]. 湖泊科学, 31(1): 147-158. |
| 曹文钟, 孙玉凤, 杨双, 等. 2019. 大庆库里泡浮游植物群落结构及多样性研究[J]. 生态科学, 38(2): 206-211. |
| 陈倩, 李秋华, 胡月敏, 等. 2018. 贵州百花水库浮游藻类功能群时空分布特征及影响因子分析[J]. 环境科学研究, 31(7): 1266-1274. |
| 陈贺, 刘子方, 吕军, 等. 2016. 淡水沉积物硅藻图谱[M]. 北京: 化学工业出版社. |
| 高彩凤, 李学军, 毛战坡. 2012. 北运河浮游植物调查及水质评价[J]. 水生态学杂志, 33(2): 85-90. |
| 国家环境保护总局. 2002. 水和废水监测分析方法(第4版)[M]. 北京: 中国环境科学出版社. |
| 胡鸿钧, 魏印心. 2006. 中国淡水藻类-系统、分类及生态[M]. 北京: 科学出版社. |
| Han Y L, Wu X, Gu J G, et al. 2016. Effects of organic acids on the photosynthetic, antioxidant properties and accumulations of heavy metals of Melilotus officinalis grown in Cu tailing[J]. Environmental Science Pollution Research, 23(18): 17901-17909. |
| 胡忠军, 莫丹玫, 周小玉, 等. 2017. 千岛湖浮游植物群落结构时空分布及其与环境因子的关系[J]. 水生态学杂志, 38(5): 46-54. |
| Jakobsen H H, Blanda E, Staehr P A, et al. 2015. Development of phytoplankton communities:Implications of nutrient injections on phytoplankton composition, pH and ecosystem production[J]. Journal of Experimental Marine Biology and Ecology, 473: 81-89. |
| 贾海燕, 徐建锋, 雷俊山. 2019. 丹江口库湾浮游植物群落与环境因子关系研究[J]. 人民长江, 50(5): 52-58. |
| 贾慧娟, 赖子尼, 王超. 2019. 珠三角河网浮游植物物种丰富度的时空特征[J]. 生态学报, 39(11): 1-11. |
| 君珊, 王东波, 周健华, 等. 2019. 拉萨河流域浮游植物群落结构特征及与环境因子的关系[J]. 生态学报, 39(3): 787-798. |
| Kim H, Spivack A J, Menden-Deuer S. 2013. pH alters the swimming behaviors of the raphidophyte Heterosigma akashiwo:Implications for bloom formation in an acidified ocean[J]. Harmful Algae, 26: 1-11. |
| Kraus C N, Bonnet M-P, Nogueira I S, et al. 2019. Unraveling flooding dynamics and nutrients' controls upon phytoplankton functional dynamics in Amazonian floodplain lakes[J]. Water, 11(1): 1-17. |
| Lee T A, Rollwagen-Bollens G, Bollens S M. 2015. The influence of water quality variables on cyanobacterial blooms and phytoplankton community composition in a shallow temperate lake[J]. Environmental Monitoring and Assessment, 187(6): 1-14. |
| 李红飞, 林森杰. 2019. 南海浮游植物生态学研究进展[J]. 厦门大学学报(自然科学版), 58(1): 1-10. |
| 李建茹, 李兴. 2017. 基于SOM的乌梁素海浮游植物群落结构研究[J]. 生态环境学报, 26(4): 649-657. |
| 李晶.2012.扎龙湿地浮游植物生态特征及其环境效应研究[D].哈尔滨: 哈尔滨工业大学 |
| 李磊, 李秋华, 焦树林, 等. 2015. 阿哈水库浮游植物功能群时空分布特征及其影响因子分析[J]. 环境科学学报, 35(11): 3604-3611. |
| 李林, 朱伟, 罗永刚. 2012. 钙、镁离子在水流作用下对铜绿微囊藻生长的影响[J]. 环境科学与技术, 35(5): 9-13. |
| 李文君.2012.黄、东海春季网采浮游植物中部分金属元素组成研究[D].青岛: 中国海洋大学 |
| 刘绍俊, 吉正元, 普发贵, 等. 2019. 星云湖浮游植物群落结构及水体营养状态生物评价[J]. 安全与环境学报, 19(4): 1439-1447. |
| 陆欣鑫, 刘妍, 范亚文. 2014. 呼兰河湿地夏-秋两季浮游植物功能分组演替及其驱动因子[J]. 生态学报, 34(5): 1264-1273. |
| 马思琦, 杨柏贺, 王汨, 等. 2019. 北运河水系春、秋季轮虫群落结构特征及其与水环境因子的关系[J]. 水产科技情报, 46(6): 345-349. |
| 闵海丽, 蔡三娟, 徐勤松, 等. 2012. 外源钙对黑藻抗镉胁迫能力的影响[J]. 生态学报, 32(1): 256-264. |
| Nunes M, Adams J B, Rishworth G M. 2018. Shifts in phytoplankton community structure in response to hydrological changes in the shallow St Lucia Estuary[J]. Marine Pollutioin Bulletin, 128: 275-286. |
| Perkins R G, Underwood G J C. 2000. Gradients of chlorophyll a and water chemistry along an eutrophic reservoir with determination of the limiting nutrient by in situ nutrient addition[J]. Water Research, 34(3): 713-724. |
| Pielou E C. 1966. Species diversity and pattern diversity in the study of ecological succession[J]. Journal of Theoretical Biology, 10(2): 370-383. |
| Pramanik S, Gani M A, Alfasane M A, et al. 2016. Seasonality of phytoplankton and their relationship with some environmental factors in a pond of old Dhaka[J]. Bangladesh Journal of Botany, 45(1): 195-201. |
| 邱阳凌, 林育青, 刘俊杰, 等. 2018. 淮河干流及主要支流夏季浮游植物群落生物多样性评价[J]. 环境科学学报, 38(4): 1665-1672. |
| Reed M L, Pinckney J L, Keppler C J, et al. 2016. The influence of nitrogen and phosphorus on phytoplankton growth and assemblage composition in four coastal, southeastern USA systems[J]. Estuarine, Coastal and Shelf Science, 177: 71-82. |
| Redfield A C. 1958. The biological control of chemical factors in the environment[J]. American Scientist, 46(3): 205-221. |
| 任辉, 田恬, 杨宇峰, 等. 2017. 珠江口南沙河涌浮游植物群落结构时空变化及其与环境因子的关系[J]. 生态学报, 37(22): 7729-7740. |
| Sellami I, Hamza A, EI Bour M, et al. 2016. Succession of phytoplankton and zooplankton communities coupled to environmental factors in the oligo-mesotrophic Nabhana reservoir(Semiarid mediterranean area, central Tunisia)[J]. Zoological Studies, 55: 1-15. |
| Shen Z Y, Hou X S, Li W, et al. 2014. Relating landscape characteristics to non-point source pollution in a typical urbanized watershed in the municipality of Beijing[J]. Landscape and Urban Planning, 123: 96-107. |
| Shi J Q, Wu Z X, Song L R. 2013. Physiological and molecular responses to calcium supplementation in Microcystis aeruginosa(Cyanobacteria)[J]. New Zealand Journal of Marine & Freshwater Research, 47(1): 51-61. |
| Sitta K A, Reed M, Mortensen R, et al. 2018. The influences of nitrogen form and zooplankton grazing on phytoplankton assemblages in two coastal southeastern systems[J]. Limnology and Oceanography, 63(6): 2523-2544. |
| 宋勇军, 戚菁, 刘立恒, 等. 2019. 程海湖夏冬季浮游植物群落结构与富营养化状况研究[J]. 环境科学学报, 39(12): 4106-4113. |
| 孙林丹, 沈建忠, 艾桃山, 等. 2019. 放养鱼类对乌梁素海浮游植物群落结构的影响[J]. 广东海洋大学学报, 39(3): 1-9. |
| 孙祥, 朱广伟, 杨文斌, 等. 2017. 天目湖沙河水库浮游植物群落结构的时空异质性[J]. 环境科学, 38(10): 4160-4168. |
| 陶志英, 陈文静, 余智杰, 等. 2017. 太泊湖浮游植物群落结构特征及其与环境因子相关性分析[J]. 安徽农业科学, 45(13): 63-67. |
| Turner R E, Rabalais N N. 2013. Nitrogen and phosphorus phytoplankton growth limitation in the northern Gulf of Mexico[J]. Aquatic Microbial Ecology, 68(10): 159-169. |
| 王晓凤, 金桂琴, 贺晓庆, 等. 2011. 北运河(通州段)浮游植物群落与水体营养状况分析[J]. 世界科技研究与发展, 33(5): 802-804. |
| 王为民.2013.2012年夏季暴雨前后胶州湾浮游植物群落的变化[D].青岛: 中国海洋大学 |
| Wu Z S, Kong M, Cai Y J, et al. 2019. Index of biotic integrity based on phytoplankton and water quality index:Do they have a similar pattern on water quality assessment? A study of rivers in Lake Taihu Basin, China[J]. Science of the Total Environmental, 658: 395-404. |
| 吴天浩, 刘劲松, 邓建明, 等. 2019. 大型过水性湖泊-洪泽湖浮游植物群落结构及其水质生物评价[J]. 湖泊科学, 31(2): 440-448. |
| 徐宗学, 殷旭旺, 武玮, 等. 2016. 渭河流域常见水生生物图谱[M]. 北京: 中国水水利水电出版社. |
| 严广寒, 殷雪妍, 汪星, 等. 2019. 长江三口-西洞庭湖环境因子对浮游植物群落组成的影响[J]. 中国环境科学, 39(6): 2532-2540. |
| 杨俊丽, 李希磊, 于潇, 等. 2019. 2016年莱州湾扇贝养殖区浮游植物群落生态特征[J]. 中国海洋大学学报(自然科学版), 49(8): 27-36. |
| 张家铭, 李炳华, 毕二平, 等. 2019. 北运河流域(北京段)沉积物中PAHs污染特征与风险评估[J]. 环境科学研究, 32(11): 1852-1860. |
| 赵文. 2005. 水生生物学[M]. 北京: 中国农业出版社. |
| Zheng W, Sugie J. 2019. Global asymptotic stability and equiasymptotic stability for a time-varying phytoplankton-zooplankton-fish system[J]. Nonlinear Analysis:Real World Applications, 46: 116-136. |
| 朱忆秋, 吕俊, 李乔臻, 等. 2019. 亚热带水库浮游植物群落的季节变化及其与环境的关系-两种功能群分类法的比较[J]. 应用生态学报, 30(6): 2079-2086. |
| Zhou Y Q, Huang S Z, Yu S L, et al. 2010. The physiological response and sub-cellular localization of lead and cadmium in Iris pseudacorus L[J]. Ecotoxicology, 19(1): 69-76. |
