 , 黄蔚1, 陈开宁1,3
, 黄蔚1, 陈开宁1,3 , 易齐涛2, 李恒鹏1
, 易齐涛2, 李恒鹏11. 中国科学院南京地理与湖泊研究所, 湖泊与环境国家重点实验室, 南京 210008;
2. 安徽理工大学, 地球与环境学院, 淮南 232001;
3. 江苏高校水处理技术与材料协同创新中心, 苏州 215009
收稿日期: 2019-09-20; 修回日期: 2020-01-13; 录用日期: 2020-01-13
基金项目: 中国科学院战略性先导科技专项(A类)(No.XDA23020403);中国科学院“一三五”计划项目(No.NIGLAS2018GH01);国家自然科学基金(No.51409166)
作者简介: 汪琪(1995-), 女, E-mail:wangqiaust@163.com
通讯作者(责任作者): 陈开宁, E-mail:knchen@niglas.ac.cn
摘要:在2015—2017年以月频率对大溪水库浮游植物群落结构及水体理化性质进行调查分析,以期为深入理解亚热带水库生态系统结构及功能提供理论依据,并为大溪水库及其同类型水体的生态保护提供基础数据.结果表明,调查期间大溪水库水质相关指标基本达到《地表水环境质量标准》(GB3838—2002)Ⅱ~Ш类标准,综合富营养化指数范围为37~54,基本处于中营养状态(除2017年9月及10月),浮游植物多样性及均匀度指数指示水体处于较为清洁的环境.2015—2017年,大溪水库共检出浮游植物7门57种,其中绿藻门种类最多,检出24种,其次是蓝藻门;且浮游植物群落中主要以蓝藻门、绿藻门及硅藻门种类占优;浮游植物群落结构季节变化有明显的规律,为硅藻门(春季)-蓝、绿藻门(夏、秋季)-硅藻门(冬季);物种丰度及生物量变化趋势为夏季高冬季低,丰度为339.87×104~33075.99×104 cells·L-1,生物量为2.41~40.45 mg·L-1.浮游植物生物量与环境因子之间的冗余分析及Pearson相关分析结果均表明,大溪水库浮游植物群落结构变化受温度影响较大.综上,在北亚热带地区低营养水平的水库中,温度可能是影响浮游植物群落结构的关键因素.
关键词:浮游植物群落结构营养状态大溪水库
Phytoplankton community structure and trophic status evaluation in Reservior Daxi
WANG Qi1,2
, HUANG Wei1, CHEN Kaining1,3, YI Qitao2, LI Hengpeng11. Sdate Key Laboratory of Lack Science and Environment, Nanjing Institute of Geography and Limnology, Chinese Academic of Sciences, Nanjing 210008;
2. School of Earth and Environment, Anhui University of Science and Technology, Huainan 232001;
3. Collaborative Innovation Center of Water Treatment Technology & Material, Suzhou 215009
Received 20 September 2019; received in revised from 13 January 2020; accepted 13 January 2020
Abstract: In order to provide a theoretical basis for understanding the structure and function of the subtropical reservoir ecosystem, and to provide basic data for the ecological protection of Reservior Daxi and the water bodies of the same type, we investigated the phytoplankton and analyzed physicochemical property of the water in Reservior Daxi from 2015 to 2017.The results showed that, during the investigating period the water quality related indexes of Reservoir Daxi basically reached the "Surface Water Environmental Quality Standard" (GB3838- 2002) II~Ш standard, the trophic level index ranged from 37 to 54, basically in the middle nutrition status (except September and October 2017), the Shannon-Wiener index and Pielou index indicated that the water body was in a relatively clean environment. From 2015 to 2017, a total of 7 phylum and 57 species of phytoplankton were detected in Reservoir Daxi, among them, the Chlorella was the major phylum, of which 24 species were detected, the Cyanophyta was the second big phylum; and the phytoplankton community was dominated by Cyanophyta, Chlorella and Diatom. The seasonal changes of phytoplankton community structure had an obvious rule, which was Diatom(in spring)-Chlorella and Cyanophyta(in summer and autumn)-Diatom(in winter). The species density and biomass change was high in summer and low in winter, the density range was 339.87×104~33075.99×104 cells·L-1, and the biomass range was 2.41~40.45 mg·L-1. Redundancy analysis and Pearson correlation analysis between phytoplankton biomass and environmental factors indicated that the change of phytoplankton community structure in Reservoir Daxi was greatly affected by temperature. In summary, in the reserviors with low nutrient levels in the northern subtropical region, the temperature may be a key factor affecting the structure of phytoplankton community.
Keywords: phytoplanktoncommunity structurenutritional statusReservior Daxi
1 引言(Introduction)浮游植物是水生态系统的初级生产者, 也是水体中溶解氧的主要来源, 同时对水环境变化有指示作用(Chapman et al., 1998; Vera et al., 1998; 邓义祥等, 1999; 王新华等, 2004).浮游植物群落的季节演替是一个复杂的生态过程, 由多因素共同参与, 其演替过程受到温度、光照、营养盐等可利用资源所产生的上行效应的影响, 同时又受到由浮游动物和滤食性鱼类等产生的下行效应的影响(Abrantes et al., 2006; Ursula et al., 2010; Sommer et al., 2012; 陈晓江等, 2016); 且浮游植物如蓝藻门、硅藻门等生长过于旺盛时会对水环境甚至人体健康造成一定的危害(Preston et al., 1980; 孔繁翔等, 2005; Steinberg et al., 2010; 乔倩, 2016).因此, 浮游植物监测是湖泊、河流和水库水环境监测的重要内容, 尤其是在饮用水源地(郭沛涌等, 2009; Katsiapi et al., 2011).水库是地球上重要的淡水资源, 是我国调蓄和供水的主要形式之一, 也是重要的水源地形式, 与人类生活息息相关, 可以减轻洪涝灾害, 维护流域生态平衡, 因此, 水库的水环境健康对保障城市安全用水有重要的意义(Guo, 2007; 韩博, 2010;侯伟等, 2016).水库水体补给来源简单, 水体环境易受来水水质影响(谢飞, 2014; 吴晓辉等, 2015).同时, 水库生态环境较为特殊, 与天然湖泊有较大差异(李秋华等, 2007).研究表明, 不同功能的水库其用水模式相差较大, 导致浮游植物生存环境有明显差别, 主要表现在种类组成和动态特征的差别(Straskraba et al., 1999; 林秋奇等, 2003; 韩亚慧, 2012).
大溪水库为太湖流域重要的水源性水库, 位于江苏省溧阳市, 属亚热带水库(刘淼等, 2017).近年来, 尽管对亚热带湖泊及水库的浮游植物生态结构特征及其影响因子的研究较多, 但基于长时间序列的研究不足, 难以揭示长时间尺度的浮游植物群落演替规律及特征, 以及影响其结构分布的环境因子(崔扬等, 2014; 刘林峰等, 2018; 吴天浩等, 2019); 且对营养水平较高的水体研究较多, 如太湖和洪泽湖(孟顺龙等, 2009; 吴天浩等, 2019; 夏莹霏等, 2019), 对营养水平较低的湖泊研究较少.自2008年以来, 随着地方经济及水库周边旅游产业的发展, 大溪水库水体环境受到一定程度的冲击, 水库的水质处于缓慢下降的状态, 因此, 对其水体生态环境进行监测和分析具有一定的现实意义(柏翔等, 2009; 刘淼等, 2016).前人对大溪水库的研究多停留在对水体营养状态和污染因子的探索上, 未曾进行过浮游植物群落及与环境因子关系的研究.基于此, 本研究对大溪水库浮游植物群落结构特征及其变化规律进行分析, 运用多元统计分析及相关性分析对其浮游植物群落结构及生物量变化与环境因子之间的相关关系进行探索, 同时在运用物理化学指标对水体水质进行评价的基础上结合生态学方法对水体进行评价, 以期为深入理解亚热带水库生态系统结构及功能提供理论依据, 并为大溪水库及其同类型水体的生态保护提供基础数据.
2 材料与方法(Materials and methods)2.1 研究区域大溪水库(31°20′34.8″~31°24′4.61″N, 119°22′1.67″~119°23′49.28″E)位于江苏省溧阳市的大溪河上游, 常州市天目湖风景区南侧, 集水面积为90 km2, 水库面积近10 km2, 属于太湖流域西区南河水系天目湖流域.水库建成于1960年6月, 是一座以防洪、城镇供水、灌溉为主, 结合旅游、水产养殖等综合利用的大型水库.大溪水库的补给主要来源为降水及地表径流, 其在东南部通过调度河(沙溪河)与沙河水库相连, 西南部有洙漕河汇入.当地气候为北亚热带季风气候, 四季分明, 雨量充沛(邬明伟等, 2010; 柏祥等, 2010; 刘淼等, 2017).流域土地利用类型以林地和农业用地为主, 以城镇居民用地和茶园为辅.
2.2 样品采集与分析在大溪水库均匀布设4个采样点(图 1), 2015年1月—2017年12月以月频率进行样品采集.浮游植物样品采用1000 mL有机玻璃采水器于水面下0.5 m处采集水样1000 mL, 现场加入鲁哥试剂15 mL并摇匀, 带回实验室静置沉淀24 h后浓缩并定容至30 mL供浮游植物定量分析.浮游植物鉴定采用视野计数法, 充分摇匀浓缩液后立即取0.1 mL样品于0.1 mL(20 mm×20 mm)浮游植物计数框内, 于10×40倍光学显微镜下进行鉴定.样品鉴定分析参照相关手册和研究方法(章宗涉等, 1991; 胡鸿钧等, 2006).由于浮游植物的比重接近于1, 故可以直接由浮游植物的体积换算为生物量(湿重), 即生物量为浮游植物的数量乘以各自的平均体积, 单位为mg · L-1, 单细胞的生物量主要根据浮游植物个体形状测量而得(章宗涉等, 1991).
图 1(Fig. 1)
 |
| 图 1 大溪水库采样点位示意图 Fig. 1Schematic diagram of sampling points of Reservoir Daxi |
采集浮游植物样品的同时, 鉴于1号点水深较浅, 基本处在3 m以下, 用采水器采集水面下0.5 m处的表层水样2 L, 2~4号点水深基本保持在3~10 m, 所采样样品为表层及底层水体的混合样.用于测定的水质理化指标包括:溶解氧、pH值、电导率、透明度、总氮(TN)、总磷(TP)、铵态氮(NH4+-N)、硝态氮(NO3--N)、亚硝态氮(NO2--N)、磷酸盐(PO43--P)、化学需氧量(CODMn)及叶绿素a(Chl a).其中, 溶解氧、pH值及电导率采用YSI ProPlus多参数水质监测仪现场测定, 用塞氏盘(Secchi disk)测定透明度, 水深仪SM-5A测定水深, 其余指标测定的详细方法及步骤参考相关国家标准(中国标准出版社第二室, 2010)及《水和废水检测分析方法》(国家环境保护总局《水和废水监测分析方法》编委会, 2002).因气象温度和水温之间有较好的相关性, 故本研究中所使用的温度为气象温度,数据来自江苏溧阳气象站.
2.3 数据分析2.3.1 浮游植物分析方法通过Mcnaughton优势度指数(Y)、Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J)对大溪水库浮游植物的生态学特征进行分析, 上述各项指数的计算公式(孟顺龙等, 2009)如下所示:
 | (1) |
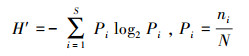 | (2) |
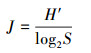 | (3) |
表 1(Table 1)
| 表 1 Shannon-Wiener指数及Pielou指数的评价标准 Table 1 Evaluation criteria of Shannon-Wiener index and Pielou index | ||||||||||||||||||||
表 1 Shannon-Wiener指数及Pielou指数的评价标准 Table 1 Evaluation criteria of Shannon-Wiener index and Pielou index
| ||||||||||||||||||||
对水库浮游植物物种生物量和所监测的环境因子进行多元统计分析.首先对数据进行去趋势对应分析(DCA), 以检验其第一轴的长度, 若梯度长度 < 3则选择冗余分析(RDA), 反之, 选择典范对应分析(CCA).
RDA是一种约束性线性直接梯度排序方法, 能够独立保持各个环境变量对生物群落变化的贡献率, 其样方排序值即反映了物种组成及生态重要值对群落的作用, 同时也反映了环境因子的影响(张元明等, 2004).同时, 对浮游植物生物量和环境因子进行Pearson相关性分析.
2.3.2 综合营养状态评价指数参考湖泊富营养化评价方法及分级标准(王明翠等, 2002), 根据Chl a、TN、TP、SD及CODMn的测定结果, 运用综合营养状态指数法对大溪水库生态系统健康状况进行评价, 具体计算方法如下:
 | (5) |
综合富营养化指数(0~100)分级标准参考《中国水资源公报》(中华人名共和国水利部, 2012), 其中, 0~30为贫营养, 30~50为中营养, 50~60为轻度富营养, 60~70为中度富营养, 70~100为高度富营养.
3 结果分析(Results analysis)3.1 水体非生物因子特征由表 2可知, 调查期间温度年变化趋势基本一致, 年际温度最高值呈逐年小幅上升趋势, 温度范围为3.32~30.18 ℃, 最高值出现在2017年7月, 为30.18 ℃; 水体pH为7.40~8.85, 处于弱碱性状态; 水体溶解氧含量呈现冬季高夏季低的趋势, 年际溶解氧最低值分别出现在2015年7月、2016年8月及2017年7月, 溶解氧含量分别为(6.72±0.24)、(4.62±0.14)、(6.73±0.29) mg · L-1, 年际最高值出现在1月及12月; 调查期间透明度为0.68~2.55 m, 电导率为125.90~195.50 μS · cm-1.
表 2(Table 2)
| 表 2 大溪水库水体水质指标 Table 2 Water quality index of Reservoir Daxi | ||||||||||||||||||||||||||||||||||||
表 2 大溪水库水体水质指标 Table 2 Water quality index of Reservoir Daxi
| ||||||||||||||||||||||||||||||||||||
2015—2017年大溪水库水体水质化学指标变化见图 2.由图可知, 2015年TN含量变化范围较大, 2016、2017年波动趋势大致相同, 其变化范围在调查期间各年份基本相似, 年际含量均值有逐年上升趋势; TP含量变化无明显规律, 2017年波动幅度较其余两年略大且其含量有上升的趋势, 3年TP含量均值分别为(0.0216±0.0097)、(0.0209±0.0096)及(0.0362±0.0202) mg · L-1; CODMn年变化趋势大致相同, 均为夏季较高; 调查期间, Chl a含量有逐年上升趋势, 年际最高值均出现在9月, 分别为(11.53±3.10)、(19.20±4.05)、(21.08±10.26) μg · L-1; NH4+-N、NO3--N、NO2--N及PO43--P在调查期间的平均含量分别为(0.14±0.12)、(0.29±0.17)、(0.0103±0.0085)及(0.0038±0.0030) mg · L-1.
图 2(Fig. 2)
 |
| 图 2 大溪水库水体水质化学指标 Fig. 2Chemical index of water quality in Reservoir Daxi |
3.2 浮游植物群落结构3.2.1 浮游植物种类组成及优势种在调查期间共监测到浮游植物7门57种, 其中, 绿藻门24种、蓝藻门11种、硅藻门12种、裸藻门3种、甲藻门3种、隐藻门2种、金藻门及黄藻门各1种.
优势种种类个数为31种, 其中, 绿藻门11种、蓝藻门10种、硅藻门7种、裸藻门、隐藻门及金藻门均为1种.绿藻门的栅列藻(Scenedesmus)占优频率最高, 其次为蓝藻门的席藻(Phormidium)和绿藻门的十字藻(Crucigenia), 说明这3种藻类在大溪水库适应能力较强; 排位第四的为硅藻门的曲壳藻(Achnanthe)和小环藻(Achnanthes).浮游植物中, 2015年春季(3—5月)小环藻均占据第一优势地位, 且优势度越来越显著, 2016年及2017年春季小环藻在3、4月占据绝对优势, 5月其优势不明显, 分别转为微囊藻(Microcystis)及曲壳藻占据第一优势地位, 调查期间春季水体浮游植物优势种以硅藻门和绿藻门种类为主, 5月有向蓝藻门转变的趋势.夏季(6—9月)第一优势种除2017年6月为绿藻门空星藻(Achnanthe)外, 其余均为蓝藻门种类, 微囊藻始终占据优势地位, 调查期间蓝藻门种类在夏季的优势度有逐年上升的趋势.秋季(9—11月)第一优势种为蓝藻门微囊藻、席藻及束丝藻(Aphanizomenon), 9、10月其优势地位极为明显, 11月有所降低, 硅藻门及绿藻门种类优势地位逐渐上升.冬季(12月、1月、2月)第一优势种有从硅藻门向绿藻门转变的趋势, 曲舟藻(Pleurosigma)及蓝隐藻仅在2016年冬季出现且占据优势地位.监测期间各月浮游植物优势属前三位及优势度见表 3.
表 3(Table 3)
| 表 3 大溪水库浮游植物种类组成 Table 3 Species composition of phytoplankton in Reservoir Daxi | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
表 3 大溪水库浮游植物种类组成 Table 3 Species composition of phytoplankton in Reservoir Daxi
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
3.2.2 浮游植物丰度及生物量变化调查期间浮游植物种类丰度及生物量变化趋势见图 4.浮游植物丰度为339.87×104~33075.99×104 cells · L-1, 呈一定的季节性变化, 除2016年8月浮游植物丰度较低外, 其余均呈现夏、秋(6—11月)大于冬、春(1—5、12月)的趋势, 藻类丰度最高值出现在2017年8月, 以蓝藻门为主.2015—2017年, 藻类年均丰度呈逐年上升趋势, 分别为2216.95×104、3143.22×104、6832.51×104 cells · L-1.调查期间以绿藻门、蓝藻门及硅藻门占主要优势, 优势种类变化规律为硅藻门-蓝、绿藻门-硅藻门, 且大部分月份为蓝藻门占主要优势.蓝藻门丰度在调查期间季节演替规律较为明显, 为夏、秋季高, 春、冬季低, 且2017年丰度大幅上升, 其峰值出现在2017年8月; 硅藻门年均丰度及其波动幅度呈逐年上升趋势; 绿藻门丰度在2015年处于相对较低水平, 各个月份之间分布较为平均, 2016年有所上升且在夏季达到丰度最大值1429.79×104 cells · L-1, 调查期间丰度均值分别为317.68×104、454.69×104、527.66×104 cells · L-1, 呈逐年上升趋势; 裸藻门变化趋势与绿藻门相同; 甲藻门年均丰度在2016年最低, 为2.79×104 cells · L-1; 隐藻门年均丰度变化不大, 2015—2017年丰度最大值逐年上升; 金藻门年均丰度分别为18.56×104、13.25×104、22.88×104 cells · L-1; 黄藻门仅在2017年6月出现, 丰度为13.39×104 cells · L-1.
图 4(Fig. 4)
 |
| 图 4 大溪水库浮游植物优势种与环境因子的冗余分析 (RDA)(S1:栅列藻; S2:十字藻; S3:盘星藻; S4:空星藻; S5:四角藻; S6:新月藻; S7:卵囊藻; S8:衣藻; S9:小球藻; S10:胶网藻; S11:纤维藻; S12:席藻; S13:蓝纤维藻; S14:色球藻; S15:裂面藻; S16:束丝藻; S17:微囊藻; S18:鱼腥藻; S19:颤藻; S20:鞘丝藻; S21:隐球藻; S22:针杆藻; S23:曲壳藻(弯杆藻); S24:颗粒直链藻; S25:小环藻; S26:曲舟藻; S27:舟形藻; S28:菱形藻; S29:囊裸藻; S30:蓝隐藻; S31:钟罩藻) Fig. 4Redundancy analysis of dominant species and environmental factors of phytoplankton in Reservoir Daxi (RDA)(S1:Scenedesmus; S2:Crucigenia; S3:Pediastrum; S4:Coelastrum Sphasricum; S5:Tetragon; S6:Closterium; S7:Oocyst; S8:Chlamydomonas; S9:Chlorella; S10:Dictyosphaerium; S11:Ankistrodesmus; S12:Phormidium; S13:Dactylococcopsis; S14:Chroococcus; S15:Merismopedia; S16:Aphanizomenon; S17:Microcystis; S18:Anabaena; S19:Oscillatoria; S20:Lyngbya; S21:Aphanocapsa; S22:Synedra; S23:Achnanthes; S24:Melosira granulata; S25:Cyclotella; S26:Pleurosigma; S27:Navicula capitatoradiata; S28:Nitzschia; S29:Trachelomonas; S30:Chroomonas; S31:Dinobryon) |
从浮游植物生物量变化趋势来看, 浮游植物生物量为2.41 ~ 40.45 mg · L-1, 硅藻门占据十分重要的地位, 其年均生物量分别为4.11、5.32和6.11 mg · L-1, 分别占总年均生物量的46.6%、47.7%和36.2%, 其次是绿藻门, 分别占总年均生物量的29.7%、30.3%和30.8%.
3.3 浮游植物群落结构与环境因子之间的关系3.3.1 浮游植物群落结构与环境因子的冗余分析(RDA)对水库浮游植物群落结构与环境因子进行DCA分析, 第一轴长度均小于3, 故采取RDA分析.根据RDA蒙特卡罗检验前向选择检验结果筛选出相关环境因子, 具体结果如图 4所示.由图可知, 2015年对群落结构起主要影响作用的环境因子为温度和透明度, 物种和环境关系的累计解释量为55.3%;2016年主成分轴1和轴2的特征值分别为0.422和0.130, 对物种和环境关系的累计解释量为59.1%, 最主要的影响因子是TN, 对浮游植物生物量变化解释了29.3%, 与TN浓度变化呈正相关的以蓝藻门为主.2017年物种和环境关系的累计解释量为59.9% (λ1=0.520, λ2=0.079), 温度对浮游植物生物量变化有显著影响, 解释率为29.7%, 除十字藻、纤维藻(Ankistrodesmus)及舟形藻(Navicula capitatoradiata)与温度变化呈负相关外, 其余均呈正相关关系.2015—2017年浮游植物生物量和环境因子的综合分析结果表明, 大部分藻种的生物量变化与温度正相关, 与氮素浓度负相关, 透明度及溶解氧含量对浮游植物生物量变化有显著影响, 物种和环境关系的累计解释量为33.3%(λ1=0.289, λ2=0.030).
无论从各年份还是长时间序列影响因子的变化来看, 温度在2015—2017年期间与浮游植物种类生物量均有较强的相关性.与温度呈正相关关系的主要是蓝藻门和绿藻门的物种, 占优频率最高的优势种栅列藻在调查期间与温度的正相关性始终较高, 但绿藻门的纤维藻在3年中与温度始终呈负相关关系.调查期间透明度对浮游植物群落的影响也不容忽视; 氮素并非始终对浮游植物群落组成产生显著的影响, 2016年TN的影响较为显著, 但在长时间序列上氮素含量仍旧占据一定地位.
3.3.2 浮游植物丰度及生物量与环境因子的Pearson相关性分析对浮游植物生物量与环境因子的Pearson相关分析结果显示(表 4), 浮游植物丰度与NO3--N浓度、溶解氧含量及透明度呈显著负相关关系(p < 0.01), 与温度呈显著正相关(p < 0.05), 相关系数为0.490;浮游植物生物量与TP含量及温度呈显著正相关关系(p < 0.01), 与溶解氧含量及透明度呈显著负相关系, 且与温度相关系数最高, 为0.651, 其次是TP含量, 其相关系数为0.573.可以看出, 影响大溪水库浮游植物丰度及生物量的环境因子种类较少.
表 4(Table 4)
| 表 4 大溪水库浮游植丰度及生物量与环境因子的相关性矩阵 Table 4 Phytoplankton abundance and correlation matrix of biomass and environmental factors in Reservior Daxi | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
表 4 大溪水库浮游植丰度及生物量与环境因子的相关性矩阵 Table 4 Phytoplankton abundance and correlation matrix of biomass and environmental factors in Reservior Daxi
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
3.4 水体营养状态评价3.4.1 综合营养状态指数评价调查期间大溪水库综合营养状态指数(TLI)为37~54, 其数值波动具有一定的规律, 即夏、秋两季较高, 冬、春两季相对较低.TLI(Chl a)的变化趋势与TLI值变化趋势相同; TLI(CODMn)相较于前两年, 2017年数值波动较大但均值有所下降; TLI(TP)数值在2017年波动相对较为平稳; TLI(TN)数值在2016年较为平稳, 2016—2017年的变化规律为7、8、9月较低, 其余月份数值较为稳定.2017年TLI值波动较大且有所上升, 指示该年份水体状态稳定性有所降低, 且水质有逐渐恶化趋势; 除2017年9、10月外, TLI值均维持在30~50之间, 指示水体处于中营养状态.
图 5(Fig. 5)
 |
| 图 5 大溪水库综合营养状态指数(TLI)变化 Fig. 5Changes in trophic level index in Reservoir Daxi |
3.4.2 生物学评价由图 6可知, 在调查期间大溪水库Shannon-Wiener多样性指数(H′)主要集中在1~4, 指示大溪水库水质处于清洁-寡污型至β-中污型, 且2016年水质总体较好.Pielou均匀度指数(J)均小于1, 变化趋势与H′相同, 表明水体水质为清洁型.从两个指数的生物学评价总体情况来看, 水体处于较为清洁的状态.
图 6(Fig. 6)
 |
| 图 6 大溪水库浮游植物多样性及均匀度指数变化 Fig. 6Changes in phytoplankton Shannon-Wiener index and Pielou index in Reservoir Daxi |
4 讨论(Discussion)浮游植物是水生生态系统生物资源的基础, 作为初级生产者, 其种群变动和群落结构直接影响着水生生态系统的结构和功能(孟顺龙等, 2009).水生生态系统浮游植物群落结构的变化与环境因子密切相关, 是其对环境适应的结果(Xiao et al., 2008).因此, 浮游植物生物量和物种组成的变化可以反映环境条件的变化(Suikkanen et al., 2007).各种群丰度由于受营养盐和温度等环境因子的驱动而处于动态变化之中(马沛明等, 2016; 万蕾等, 2007; Elliott, 2010; Lu et al., 2018).大量研究表明, 浮游植物群落存在明显的季节演替现象(Sommer et al., 2012; 马沛明等, 2016; 郑诚等, 2018).从浮游植物丰度来看, 大溪水库浮游植物群落结构变化趋势为硅藻-蓝、绿藻-硅藻, 这与Sommer等(1986)的研究结果基本一致, 该研究认为, 中营养型湖泊中浮游植物的季节演替规律为从冬、春季的隐藻和硅藻占优转变为夏季的绿藻, 紧接夏末秋初为蓝藻占优势, 到秋季为硅藻重新占优.在本研究中, 大溪水库冬季隐藻的优势并不明显, 可能是因为大溪水库营养盐、有机质及悬浮质浓度保持在较低的水平, 使隐藻的竞争优势不显著(刘霞等, 2012).作为水体重要的初级生产者, 温度、光照等光合作用基本条件无疑对浮游植物的生长起着极其重要的作用(杨强等, 2011).根据图 3及图 4可以发现, 浮游植物的生长具有一定的季节性规律且温度对浮游植物种类结构组成的影响较为显著.夏季浮游植物以蓝藻为主且丰度达到最大值, 因为蓝藻的最适温度为25~35 ℃(陈开宁等, 2007), 故在夏、秋季快速增殖并占据绝对优势.微囊藻为夏季的主要优势种, 可以发现其生物量变化在调查期间受温度影响显著, 这与Takamura等(1985)的研究结果相同.温度对微囊藻的生长具有重要的影响, 当温度较低时, 微囊藻的生长受到抑制(吴溶等, 2010), 并形成休眠体, 以这种形式暂存于底泥中, 当温度上升时微囊藻休眠体复苏(Rengefors et al., 1998), 当温度达到15 ℃时, 其生长速率增大, 并且开始少量迁移到水体中(李阔宇等, 2004); 微囊藻是好高温的物种, 在25~35 ℃时增殖速率随温度上升而增大(刘玉生等, 1995).冬季大溪水库浮游植物丰度降低且主要以硅藻门为主, 可能是因为硅藻所适应的温度较低(殷大聪等, 2012).从长时间序列来看, 小环藻与温度呈负相关关系, 可能是由于营养盐含量始终处于较低的水平, 硅藻的生长受到蓝藻的竞争性抑制, 导致其优势不显著.
图 3(Fig. 3)
 |
| 图 3 大溪水库浮游植物种类组成及丰度、生物量变化 Fig. 3Species composition, density and biomass change of phytoplankton in Reservoir Daxi |
通过对2015—2017年浮游植物丰度及生物量分布进行分析, 发现2017年浮游植物丰度及生物量整体均高于前两年, 可能是因为2017年水体营养盐含量的变化导致浮游植物丰度及生物量产生变化.大量研究表明, 营养盐是浮游植物存活的必要条件, 是影响浮游植物生长的关键因子, 且氮或磷含量往往是浮游植物生长的限制条件(Hecky et al., 1988; 孔繁翔等, 2005; Xu et al., 2010; 刘林峰等, 2018).当TN/TP>50时, 藻类生长以磷为主要限制因素; 当TN/TP≤20时, 氮为主要限制因子(Guildford et al., 2000). 2015—2017年大溪水库TN/TP比值分别为50.5±30.9、68.2±65.5及35.8±23.2, 因此, 大溪水库基本上属于氮磷共同限制或磷限制状态.2015及2016年TP含量差异不显著(p=0.80), 2017年TP含量与前两年差异显著, 2017年水体总磷含量较高, 与2016年相比差异不显著, 但氮磷比值显著降低, 水体环境有突破磷限制的趋势, 因此可以推测, 2017年水体磷含量的升高是导致该年份浮游植物总丰度及总生物量上升的主要原因.生物量与水体总磷含量的Person相关性分析结果也在一定程度上解释了这一现象.
温带及寒带地区年降水量少, 温度和光照是引起浮游植物群落结构变化的主要因子(Reynolds, 2006; Lund, 2010; Richardson et al., 2010).在热带与亚热带地区, 温度与光照季节性变化不如温带地区显著, 影响浮游植物群落结构的环境因子更为复杂(Nogueira, 2000; Figueredo et al., 2001).大溪水库对浮游植物群落结构起最主要作用的环境因子是温度, 这与温带地区水库的情况相似, 可能是因为大溪水库位于亚热带地区的北部, 兼具温带水库的特性, 各季节的温度变化显著, 故浮游植物的群落结构变化由温度主导.大溪水库浮游植物群落变化规律具有典型的亚热带水库特点, 即具有明显的季节变化, 春季以绿藻、隐藻、硅藻为主, 夏季以蓝藻和硅藻占优势, 秋季以硅藻隐藻占优势, 冬季以硅藻为主(张婷等, 2009; 盛海燕等, 2010; 施练东等, 2013).
根据《地表水环境质量标准》(GB3838—2002), 大溪水库水质大部分达到了地表水三类标准, 与刘淼等(2017)研究结果一致, 其主要影响因子为高锰酸盐指数和总氮.结合综合营养状态指数评价结果, 其在调查期间综合营养状态指数范围为37~54, 除2017年9月及10月外, 水体均处于中营养状态.浮游植物Shannon-Wiener多样性指数及Pielou均匀度指数显示, 水体处于清洁型-β寡污型.
调查期间, 浮游植物种类中有金藻门及甲藻门检出, 金藻门耐污性较差, 为贫营养型水体的代表物种(何志辉, 1987).从浮游植物丰度指标来看, 小于30×104 cells · L-1时水体为贫营养型, 在30×104~100×104 cells · L-1时为中营养型, 大于100×104 cells · L-1时水体为富营养型.从浮游植物生物量指标来看, 若浮游植物生物量小于1 mg · L-1, 则水体为贫营养型, 处于1~5 mg · L-1时为中营养型, 5~10 mg · L-1时水体为富营养型, 大于10 mg · L-1时为超富营养型(何志辉, 1987; 王新华等, 2004)调查期间, 大溪水库浮游植物丰度范围为339.87×104~33075.99×104 cells · L-1, 生物量范围为2.4~40.4 mg · L-1, 均值为12.3 mg · L-1.除12月及1月外, 所有监测月份浮游植物生物量均大于中营养型水体的最大阈值5 mg · L-1.根据浮游植物丰度及生物量指示, 大溪水库水体处于富营养-超富营养状态.不仅通过丰度及生物量评价的结果之间存在差异, 且与其他指标评价结果间也存在较大的差异.上游沙河水库水质基本处于Ш类标准(GB3838-2002)(崔扬等, 2014), 利用浮游植物丰度及生物量(浮游植物丰度901×104~31746×104 cells · L-1, 生物量2.30~21.94 mg · L-1)对水质进行评价的结果与大溪水库大致相同.饮用水源水库北京密云水库(武殿伟等, 2001)、湖北金沙河水库(张云等, 2015)等的水质评价结果均为中营养型水体, 但浮游植物丰度及生物量处于较高的水平, 与大溪水库相似.直接利用浮游植物丰度及生物量对水体水质进行评价的结果与其他手段评价结果产生差异的原因如下:一方面是因为藻类生物量变化趋势不仅与数量有关, 还与细胞大小有关(刘林峰等, 2018); 另一方面可能是因为水库具有较为稳定的水生生态环境, 水位稳定, 水体环境适宜浮游植物生长, 如弱碱性的水体环境(刘建康, 1999), 适宜的光照强度及溶解氧含量(由文辉, 1994).因此, 直接利用浮游植物丰度及生物量对水质进行评价的结果会与其他指标评价结果产生差异(何志辉, 1987).
5 结论(Conclusions)1) 2015—2017年, 大溪水库水体水质指标基本达到《地表水环境质量标准》(GB3838—2002)Ⅱ~Ш类标准; 综合营养状态指数表明, 调查期间大溪水库水体处于中营养状态; 根据浮游植物多样性及均匀度指数可知, 水体状态较为清洁.综上, 大溪水库水体处于中营养状态.
2) 浮游植物群落季节波动稳定, 规律为硅藻门(春季)-蓝、绿藻门(夏秋季)-硅藻门(冬季).
3) 调查期间浮游植物群落结构组成的变化与温度的相关性在各年份均达到较高的水平, 表明在北亚热带地区低营养水平的水体中, 温度可能是影响浮游植物群落结构的关键因素.
致谢:在此特向进行浮游植物鉴定工作的刘丽老师, 以及协助进行采样工作的大溪水库管理局有关人员表示诚挚的谢意.
参考文献
| Abrantes N, Antunes S C, Pereira M J, et al. 2006. Seasonal succession of cladocerans and phytoplankton and their interactions in a shallow eutrophic lake (Lake Vela, Portugal)[J]. Acta Oecologica, 29(1): 54-64. |
| 柏祥, 陈开宁, 黄蔚, 等. 2010. 大溪水库水体营养现状分析[J]. 海洋湖沼通报, (3): 115-120. |
| Chapman B R, Ferry B W, Ford T W. 1998. Phytoplankton communities in water bodies at Dungeness, U.K.:analysis of seasonal changes in response to environmental factors[J]. Hydrobiologia, 362(1/3): 161-170. |
| 陈晓江, 杨劼, 杜桂森, 等. 2016. 官厅水库浮游植物功能群季节演替及其驱动因子[J]. 中国环境监测, 32(3): 74-81. |
| 陈开宁, 周万平, 鲍传和, 等. 2007. 浮游植物对湖泊水体生态重建的响应-以太湖五里湖大型围隔示范工程为例[J]. 湖泊科学, 19(4): 359-366. |
| 崔扬, 朱广伟, 李慧赟, 等. 2014. 天目湖沙河水库水质时空分布特征及其与浮游植物群落的关系[J]. 水生态学杂志, 35(3): 10-18. |
| 邓义祥, 张爱军. 1999. 试论藻类在水体污染监测中的运用[J]. 环境与开发, (1): 43-45. |
| Elliott J A. 2010. The seasonal sensitivity of Cyanobacteria and other phytoplankton to changes in flushing rate and water temperature[J]. Global Change Biology, 16(2): 864-876. |
| Figueredo C C, Giani A. 2001. Seasonal variation in the diversity and species richness of phytoplankton in a tropical eutrophic reservoir[J]. Hydrobiologia, 445(1/3): 165-174. |
| Guildford S J, Hecky R E. 2000. Total nitrogen, total phosphorus, and nutrient limitation in lakes and oceans:Is there a common relationship?[J]. Limnology & Oceanography, 45(6): 1213-1223. |
| 郭沛涌, 沈焕庭, 刘阿成, 等. 2009. 应用浮游植物监测与评价长江口水体营养状况[J]. 海洋科学, 33(12): 68-72. |
| Guo L. 2007. Doing battle with the green monster of Taihu Lake[J]. Science, 317(5842): 1166-1166. |
| 国家环境保护总局《水和废水监测分析方法》编委会. 2002. 水和废水监测分析方法(第四版)[M]. 北京: 中国科学出版社. |
| 韩亚慧. 2012. 陕西省4座水库浮游生物的种群结构特征与营养类型研究[J]. 水生态学杂志, 33(6): 14-19. |
| 韩博平. 2010. 中国水库生态学研究的回顾与展望[J]. 湖泊科学, 22(2): 151-160. |
| 何志辉. 1987. 中国湖泊和水库的营养分类[J]. 大连海洋大学学报, 8(1): 1-10. |
| Hecky R E, Kilham P. 1988. Nutrient limitation of phytoplankton in freshwater and marine environments:A review of recent evidence on the effects of enrichment1[J]. Limnology & Oceanography, 33(4): 796-822. |
| 侯伟, 孙韶华, 贾瑞宝. 2016. 中国北方山区水库与引黄水库富营养化特征[J]. 中国环境监测, 32(2): 58-63. |
| 胡鸿钧, 魏印心. 2006. 中国淡水藻类-系统、分类及生态[M]. 北京: 科学出版社. |
| Katsiapi M, Maria M G, Evangelia M, et al. 2011. Phytoplankton and water quality in a Mediterranean drinking-water reservoir (Marathonas Reservoir, Greece)[J]. Environmental Monitoring & Assessment, 181(1/4): 563-575. |
| 孔繁翔, 高光. 2005. 大型浅水富营养化湖泊中蓝藻水华形成机理的思考[J]. 生态学报, 25(3): 589-595. |
| 李秋华, 韩博平. 2007. 南亚热带调水水库春季浮游植物群落的结构与动态[J]. 热带亚热带植物学报, 15(4): 294-300. |
| 李阔宇, 宋立荣, 万能. 2004. 底泥中微囊藻复苏和生长特性的研究[J]. 水生生物学报, 28(2): 113-118. |
| 林秋奇, 胡韧, 段舜山, 等. 2003. 广东省大中型供水水库营养现状及浮游生物的响应[J]. 生态学报, 23(6): 1101-1108. |
| 刘淼, 陈开宁, 孙淑雲, 等. 2017. 大溪水库水质变化趋势及污染成因解析[J]. 环境科学学报, 37(7): 2514-2523. |
| 刘林峰, 周先华, 高健, 等. 2018. 神农架大九湖湿地浮游植物群落结构特征及营养状态评价[J]. 湖泊科学, 30(2): 417-430. |
| 刘霞, 陆晓华, 陈宇炜. 2012. 太湖北部隐藻生物量时空动态[J]. 湖泊科学, 24(1): 142-148. |
| 刘玉生, 韩梅, 梁占彬. 1995. 光照、温度和营养盐对滇池微囊藻生长的影响[J]. 环境科学研究, 8(6): 7-11. |
| 刘建康. 1999. 高级水生生物学[M]. 北京: 科学出版社. |
| Lu J, Yang Z, Zhang Y. 2018. Algae functional group characteristics in reservoirs and lakes with different trophic levels in northwestern semi-humid and semi-arid regions in China[J]. Journal of Environmental Sciences, 64(2): 166-173. |
| Lund J W G. 2010. The Ecology of the Freshwater Phytoplankton[J]. Biological Reviews, 40(2): 231-290. |
| 马沛明, 施练东, 张俊芳, 等. 2016. 浙江汤浦水库浮游植物季节演替及其影响因子分析[J]. 环境科学, 37(12): 4560-4569. |
| Margalef D R. 1958. Inpersproctivesi marine biology (A Buzzati Traversoed)[M]. California: University California Press, 323-347. |
| 孟顺龙, 陈家长, 范立民, 等. 2009. 2007年太湖五里湖浮游植物生态学特征[J]. 湖泊科学, 21(6): 845-854. |
| Nogueira M G. 2000. Phytoplankton composition, dominance and abundance as indicators of environmental compartmentalization in Jurumirim Reservoir (Paranapanema River), S?o Paulo, Brazil[J]. Hydrobiologia, 445: 165-174. |
| Preston T, Stewart W D P, Reynolds C S. 1980. Bloom-forming cyanobacterium Microcystis aeruginosa overwinters on sediment surface[J]. Nature, 288(5789): 365-367. |
| Reynolds C S. 2006. Ecology of Phytoplankton[M]. New York: Cambridge University Press. |
| Rengefors K, Karlsson I, Hansson L A. 1998. Algal cyst dormancy:a temporal escape from herbivory[J]. Proceedings of the Royal Society B Biological Sciences, 265(1403): 1353-1358. |
| Richardson T L, Gibson C E, Heaney S I. 2010. Temperature, growth and seasonal succession of phytoplankton in Lake Baikal, Siberia[J]. Freshwater Biology, 44(3): 431-440. |
| Shannon C E, Weaver W. 1998. The Mathematical Theory of Communication[M]. Urbana: University of Illinois Press. |
| 盛海燕, 虞左明, 韩轶才, 等. 2010. 亚热带大型河流型水库-富春江水库浮游植物群落及其与环境因子的关系[J]. 湖泊科学, 22(2): 235-243. |
| 施练东, 竺维佳, 张俊芳, 等. 2013. 亚热带水库浮游植物群落结构季节演替及其春季水华成因分析-以浙江汤浦水库为例[J]. 水生态学杂志, 34(2): 32-39. |
| Sommer U, Adrian R, De L, et al. 2012. Beyond the Plankton Ecology Group (PEG) Model:Mechanisms driving plankton succession[J]. Annual Review of Ecology Evolution & Systematics, 43(1): 429-448. |
| Sommer U, Gliwicz Z M, Lampert W, et al. 1986. The PEG-model of seasonal succession of planktonic events in fresh waters[J]. Arch Hydrobiol, 106(4): 433-471. |
| Steinberg C E W, Hartmann H M. 2010. Planktonic bloom-forming Cyanobacteria and the eutrophication of lakes and rivers[J]. Freshwater Biology, 20(2): 279-287. |
| Straskraba M, Tundisi J G. 1999. Guidelines of Lake Management (Volume 9):Reservoir Water Quality Management[M]. Japan: International Lake Environment Committee, 1-60. |
| Suikkanen S, Laamanen M, Huttunen M. 2007. Long-term changes in summer phytoplankton communities of the open northern Baltic Sea[J]. Estuarine, Coastal and Shelf Science, 71(3/4): 580-592. |
| Takamura N, Iwakuma T, Yasuno M. 1985. Photosynthesis and primary production of Microcystis aeruginosa Kütz.in Lake Kasumigaura[J]. Journal of Plankton Research, 7(3): 303-312. |
| Ursula G, Miriam RB, Ina W, et al. 2010. Biotic interactions may overrule direct climate effects on spring phytoplankton dynamics[J]. Global Change Biology, 16(3): 1122-1136. |
| Vera L, Huszar D M, Caraco N F. 1998. The relationship between phytoplankton composition and physical-chemical variables:A comparison of taxonomic and morphological-functional descriptors in six temperate lakes[J]. Freshwater Biology, 40(4): 679-696. |
| 万蕾, 朱伟, 赵联芳. 2007. 氮磷对微囊藻和栅藻生长及竞争的影响[J]. 环境科学, 28(6): 1230-1235. |
| 王新华, 纪炳纯, 李明德, 等. 2004. 引滦工程上游浮游植物及其水质评价[J]. 环境科学研究, 17(4): 18-24. |
| 王明翠, 刘雪芹, 张建辉. 2002. 湖泊富营养化评价方法及分级标准[J]. 中国环境监测, 18(5): 47-49. |
| 吴晓辉, 吴钢, 潘轲旻, 等. 2015. 南水北调来水对密云水库水质和水生生物影响的预测分析[J]. 北京水务, (6): 4-6. |
| 吴天浩, 刘劲松, 邓建明, 等. 2019. 大型过水性湖泊-洪泽湖浮游植物群落结构及其水质生物评价[J]. 湖泊科学, 31(2): 440-448. |
| 邬明伟, 李兆富, 李恒鹏, 等. 2010. 太湖上游大溪水库流域营养盐输移时空特征分析[J]. 农业环境科学学报, 29(12): 2375-2382. |
| 吴溶, 崔莉凤, 卢珊, 等. 2010. 温度光照对铜绿微囊藻生长及藻毒素释放的影响[J]. 环境科学与技术, (S1): 33-36. |
| 武殿伟, 王建厅, 赵盼, 等. 2001. 密云水库的浮游植物群落结构与密度[J]. 植物生态学报, 25(4): 501-504. |
| 夏莹霏, 胡晓东, 徐季雄, 等. 2019. 太湖浮游植物功能群季节演替特征及水质评价[J]. 湖泊科学, 31(1): 136-148. |
| Xiao L J, Wang T. 2008. Grazing of Daphnia galeata and Phyllodiaptomus tunguidus on phytoplankton in Liuxihe Reservoir, South China:in situ bottle experiments[J]. Ecological Science, 27(5): 362-367. |
| 谢飞.2014.珠海供水水库富营养化现状与浮游植物群落特征[D].广州: 暨南大学 http://cdmd.cnki.com.cn/Article/CDMD-10559-1015002642.htm |
| Xu H, Paerl H W, Qin B Q, et al. 2010. Nitrogen and phosphorus inputs control phytoplankton growth in eutrophic Lake Taihu, China[J]. Limnology & Oceanography, 55(1): 420-432. |
| 杨强, 谢平, 徐军, 等. 2011. 河流型硅藻水华研究进展[J]. 长江流域资源与环境, (S1): 159-165. |
| 殷大聪, 黄薇, 吴兴华, 等. 2012. 汉江水华硅藻生物学特性初步研究[J]. 长江科学院院报, 29(2): 6-10. |
| 由文辉. 1994. 我国利用水生生物评价水质的研究进展[J]. 环境与可持续发展, (1): 8-11. |
| 章宗涉, 黄祥飞. 1991. 淡水浮游生物研究方法[M]. 北京: 中国科学出版社. |
| 张元明, 陈亚宁, 张小雷. 2004. 塔里木河下游植物群落分布格局及其环境解释[J]. 地理学报, 59(6): 903-910. |
| 张婷, 李林, 宋立荣. 2009. 熊河水库浮游植物群落结构的周年变化[J]. 生态学报, 29(6): 2971-2979. |
| 张云, 马徐发, 郭飞飞, 等. 2015. 湖北金沙河水库浮游植物群落结构及其与水环境因子的关系[J]. 湖泊科学, 27(5): 902-910. |
| 郑诚, 陆开宏, 徐镇, 等. 2018. 四明湖水库浮游植物功能类群的季节演替及其影响因子[J]. 环境科学, 39(6): 186-195. |
| 中国标准出版社第二室. 2010. 环境监测方法标准汇编[M]. 北京: 中国标准出版社. |
| 中华人名共和国水利部. 2012. 中国水资源公报[M]. 北京: 中国水利水电出版社. |
