











































 下载:
下载: 





 点击查看大图
点击查看大图全文HTML
--> --> --> 传统曝气生物滤池由于池内脱氮微生物主要为自养硝化细菌,该细菌需要消耗大量的能量,生长速度缓慢,尤其在高盐环境下难以维持较高细菌浓度,从而影响硝化作用效果,故氨氮在较短时间内得不到有效去除,而且硝化过程中易发生

异养硝化作为一种新型的生物脱氮技术,是利用异养硝化细菌在好氧条件下将氨氮或有机氮氧化成NH2OH-N、











本研究团队此前已从海洋沉积物中分离筛选到1株嗜盐异养硝化-好氧反硝脱氮菌Vibrio diabolicus SF16,该菌株不仅能够在较高盐度(1%~5%)的环境中较好地生存,而且对119.77 mg·L?1


1.1. 模拟废水配制
含

含


微量元素溶液主要包括63.70 g·L?1 Na2EDTA、5.50 g·L?1 CaCl2、3.90 g·L?1 ZnSO4·7H2O、5.06 g·L?1 MnCl2·4H2O、5.00 g·L?1 FeSO4·7H2O、1.00 g·L?1 Na2MoO4·2H2O、1.01 g·L?1 CuSO4、1.61 g·L?1 CoCl2·6H2O以及pH 7.0。
1.2. HNBAF系统对模拟含盐废水的脱氮性能
实验所用的曝气生物滤池为一个高50 cm、直径15 cm的有机玻璃圆筒,圆筒内装牡蛎壳填料,其粒径为3~5 cm,高度约30 cm,孔隙率约为80%。曝气生物滤池放置于恒温水浴箱,保持反应容器内温度为(25 ±1) ℃。利用曝气增氧机从反应器底部连续充入空气,保持溶解氧含量为3~4 mg·L?1[25-26]。该反应器接种本实验室筛选到的嗜盐异养硝化-好氧反硝化海洋菌株Vibrio diabolicus SF16[25],并进行挂膜,构建异养硝化型曝气生物滤池(HNBAF)系统。该HNBAF系统正常稳定运行超过0.5 a之后,对其开展脱氮性能测试。1) HNBAF系统对含









2) HNBAF系统对硝化反应中间产物(









3)HNBAF系统对高盐含









1.3. 利用稳定同位素示踪技术测定气态产物
分别从上述正常稳定运行的曝气生物滤池中刮取15 g生物膜,移至标记为A、B、C的3个500 mL锥形瓶中,设计3组实验。其中,标记A、B的锥形瓶各倒入300 mL含

1.4. 水质测定方法
各项水质检测指标均参考《水和废水检测分析方法》[27]规定的方法进行测定:


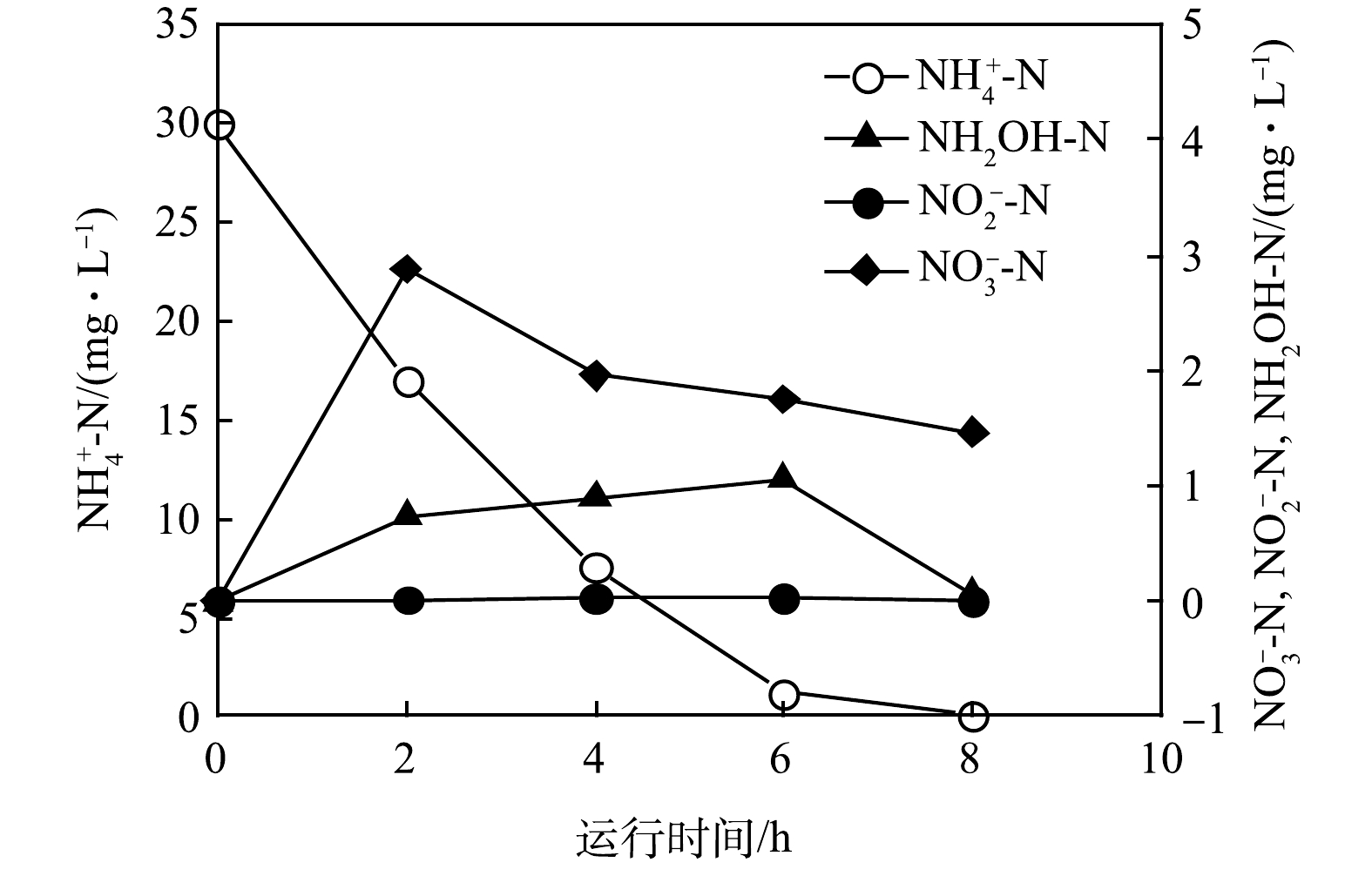
2.1. HNBAF系统对含${\rm{NH}}_4^ + $
-N高盐废水的脱氮性能
以氯化铵为唯一氮源,考察HNBAF系统对模拟高盐废水中














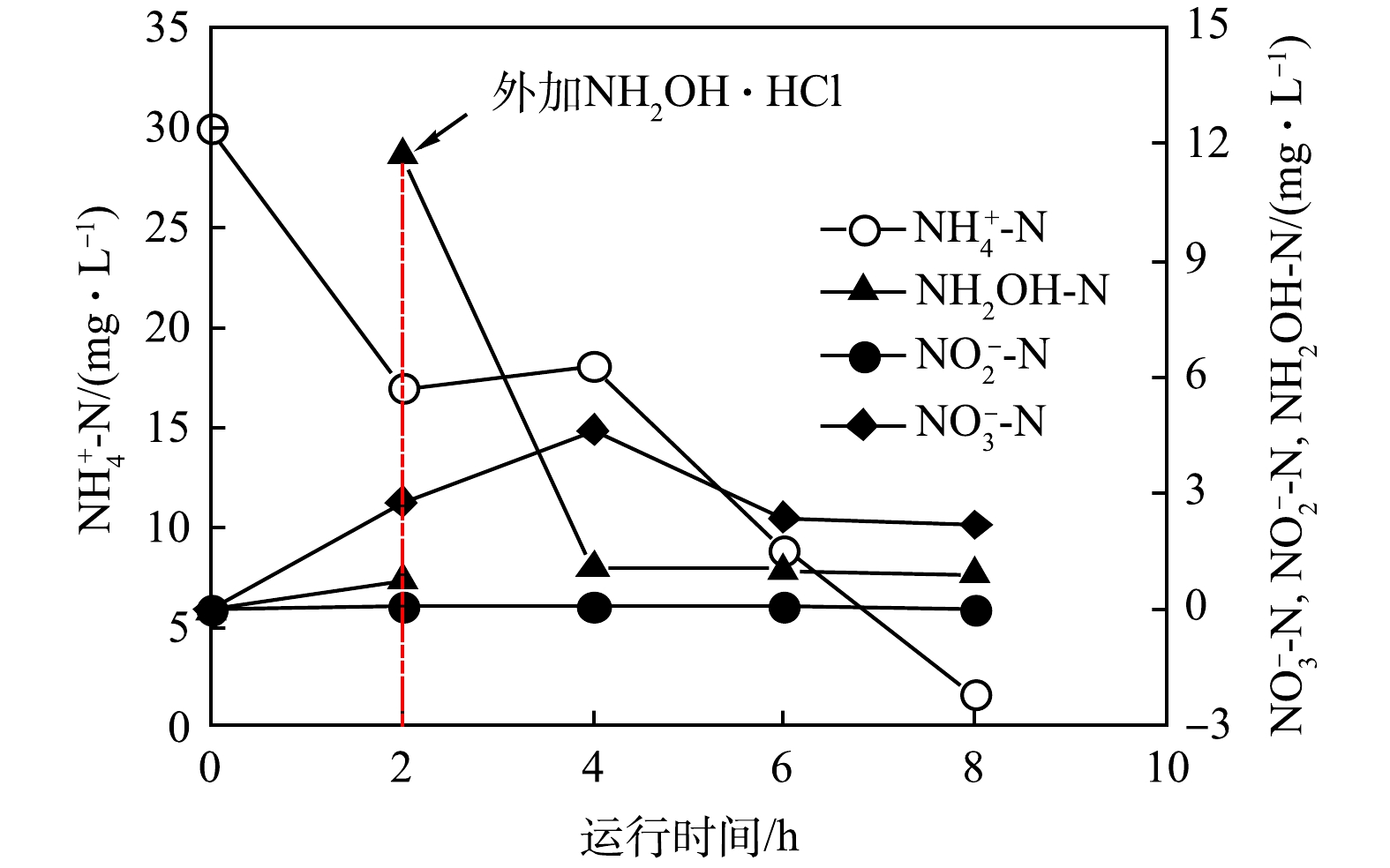
2.2. HNBAF系统对硝化反应中间产物NH2OH-N的利用
为了考察HNBAF系统对硝化反应中间产物NH2OH-N的利用情况,将含














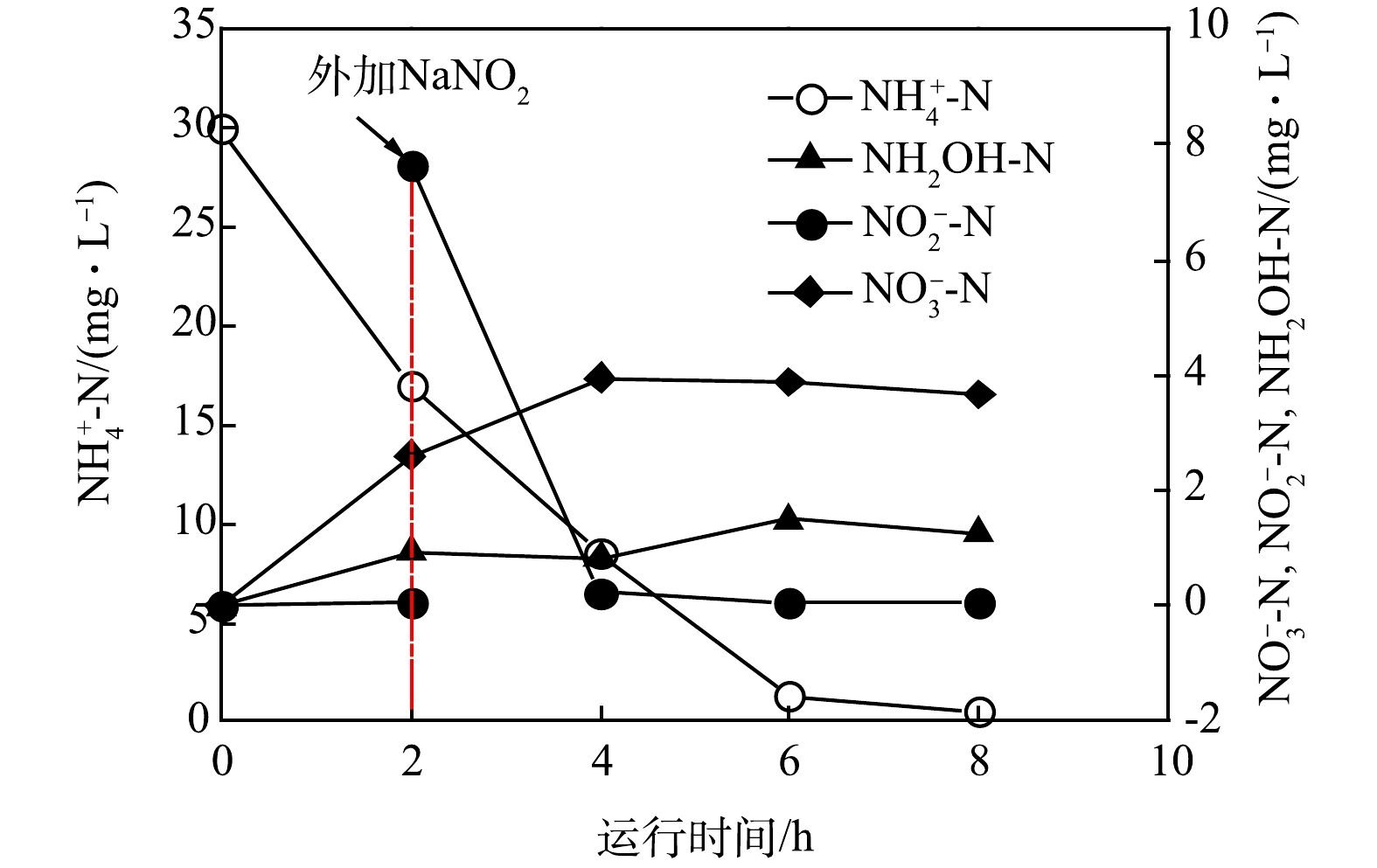
2.3. HNBAF系统对硝化反应中间产物${\rm{NO}}_2^ - $
-N的利用情况
为了考察HNBAF系统对硝化反应中间产物




















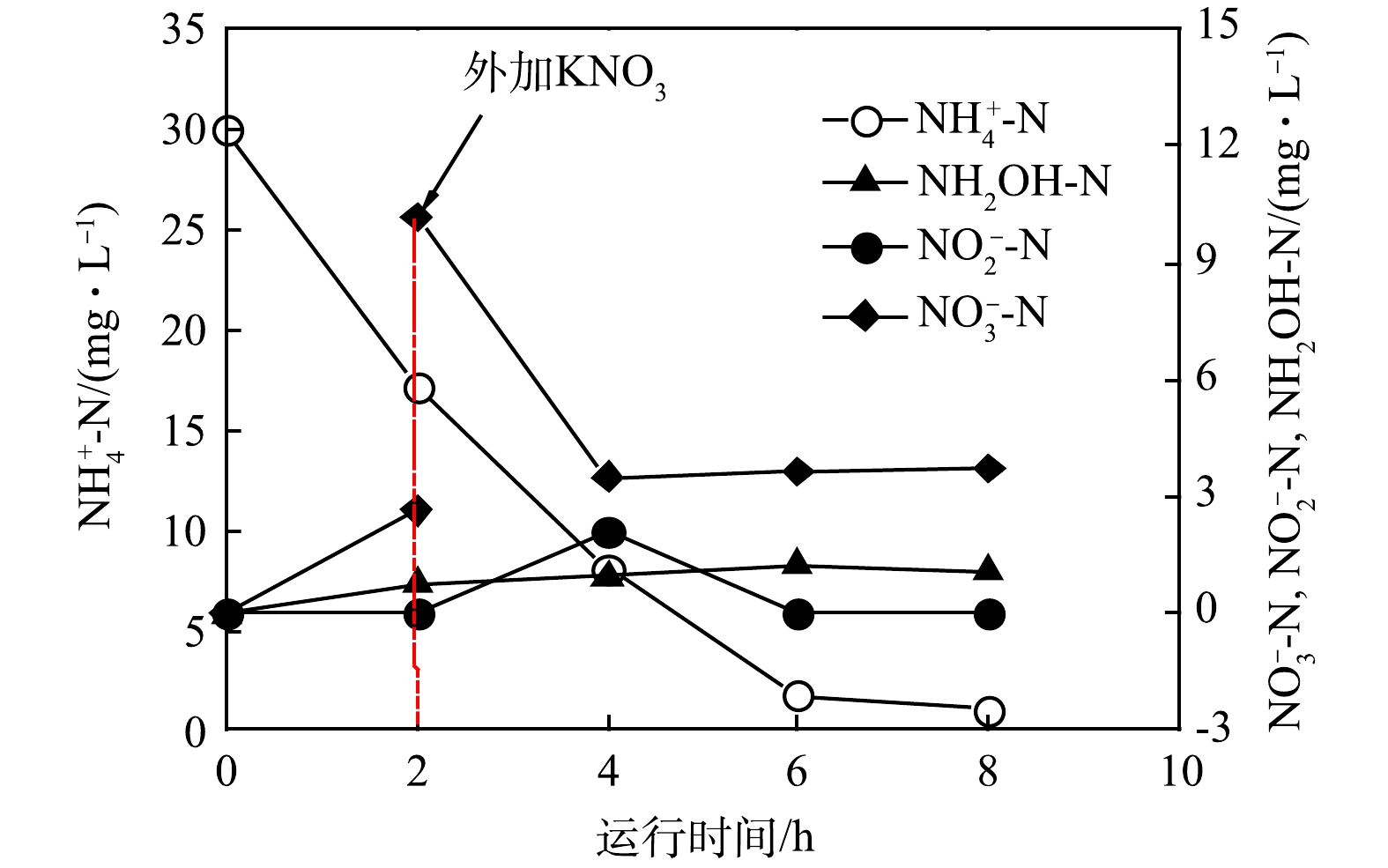
2.4. HNBAF系统对硝化反应中间产物${\rm{NO}}_3^ - $
-N的利用情况
为了考察HNBAF系统对硝化反应中间产物





















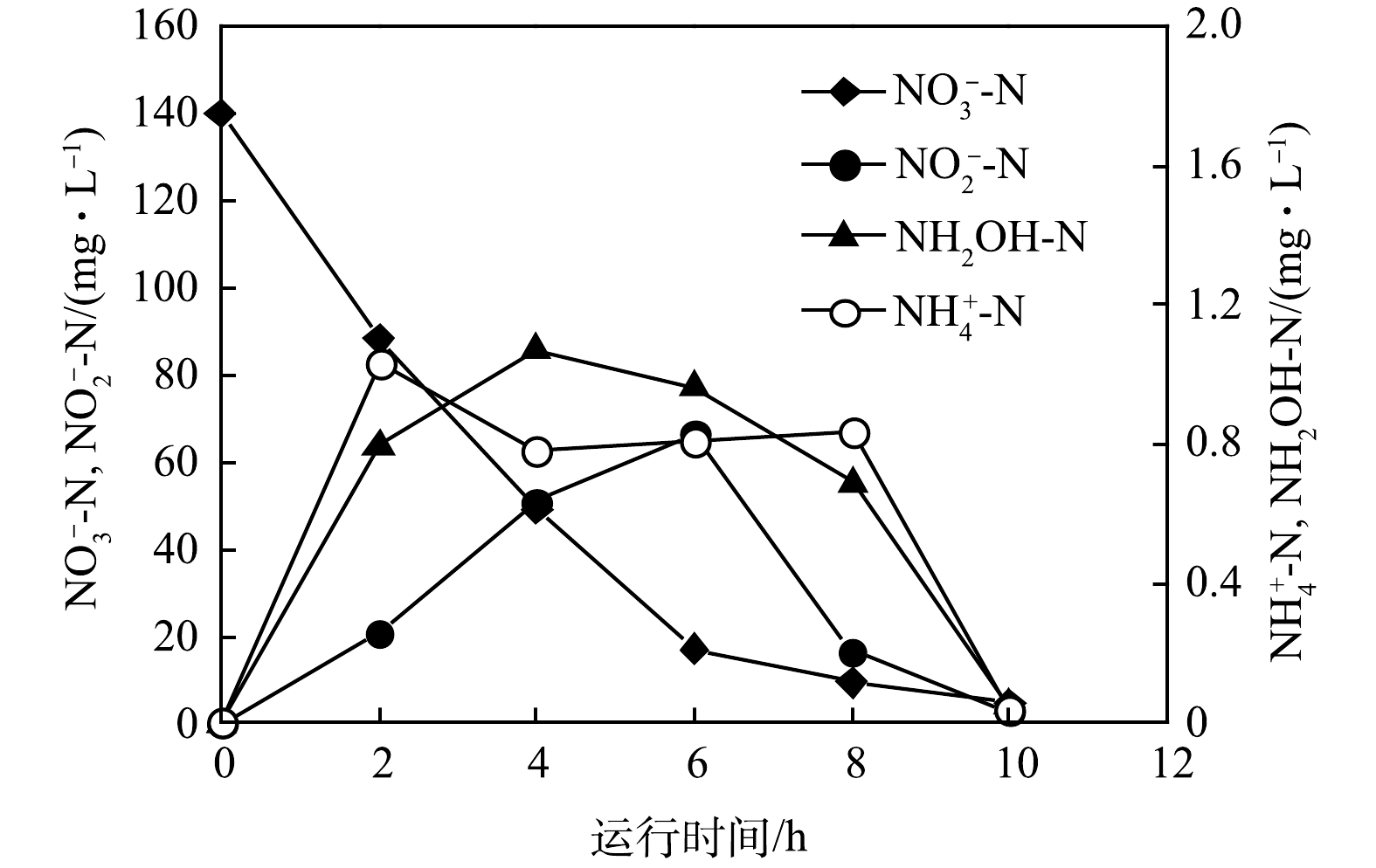
2.5. HNBAF系统对含${\rm{NO}}_3^ - $
-N高盐废水脱氮性能的影响
为了考察HNBAF系统的好氧反硝化性能,以KNO3为唯一氮源,分析该HNBAF系统8 h内对


















2.6. 利用稳定同位素示踪方法测定气态产物
表1为HNBAF系统中的生物膜对3组模拟废水处理所产生的N2O、N2的15N丰度检测结果。由表1可知,当生物膜以Na15NO2和K15NO3作为氮源时,所产生含氮气体样品中N2O和N2的δ15N值都大于0,该结果中有标记15N的N2O和N2产生,这表明HNBAF系统中的生物膜能够通过好氧反硝硝化作用将






2.7. HNBAF系统的脱氮途径
根据本实验中HNBAF系统在脱氮过程中废水中










































综上所述,HNBAF系统对高盐废水中

















因此,可以总结推导出本研究中的HNBAF系统处理高盐废水的主要脱氮途径为:在好氧条件下先通过硝化作用将




















2)当分别以NH4Cl+Na15NO2、NH4Cl+K15NO3、KNO3+Na15NO2为氮源时,均可同时检测到15N2O和15N2,说明HNBAF系统中的生物膜能够通过好氧反硝硝化作用将


3) HNBAF系统对高盐废水中









参考文献
