Tolerance mechanism of Geobacter sulfurreducens to silver ion and its effects on electricity generation in microbial fuel cell
YUAN Xin, LI Pengsong, GU Yuyi, CHEN Haoqiang, SUN Dezhi, DANG Yan, Engineering Research Center for Water Pollution Source Control and Eco-Remediation, Beijing Key Laboratory for Source Control Technology of Water Pollution, College of Environmental Science and Engineering, Beijing Forestry University, Beijing 100083, China
Abstract:Microbial fuel cell (MFC) is a hot spot research area in sewage treatment. Geobacter is widely recognized in the field of MFC due to its excellent electricity generation ability. The biologically toxic components such as heavy metals in the natural water environment will affect the growth and survival of Geobacter and the ability to generate electricity, which in turn affects the electricity generation performance of MFC. Geobacter is generally tolerant to various kinds of heavy metals such as Ag+. However, the mechanism of its tolerance to higher concentrations of heavy metals is still unclear. In this study, Geobacter sulfurreducens, a model species of Geobacter, was selected to study the regulation of the copZ gene on G. sulfurreducens’s tolerance to Ag+. Results showed that the transcription of the copZ gene of wild-type G. sulfurreducens increased by 24.8 times under 0.05 mmol·L?1Ag+. When copZ was knocked out from the G. sulfurreducens genome, the tolerance of G. sulfurreducens to Ag+ decreased significantly. The growth rate of the copZ-deficient G. sulfurreducens strain in the presence of 0.01 mmol·L?1 Ag+ was only 33.3% of that of the wild-type G. sulfurreducens in the presence of 1 mmol·L?1Ag+. When 0.05 mmol·L?1 Ag+ was added to the microbial fuel cell (MFC) system which inoculated with the copZ -deficient G. sulfurreducens strain, the output current of the MFC decreased by 6.99%. This study proved that the copZ has a significant regulatory effect on G. sulfurreducens tolerance to Ag+, and revealed the internal mechanism of high Ag+ concentration influence on the electricity productivity performance in the G. sulfurreducens MFC system. Key words:microbial fuel cell/ Geobacter sulfurreducens/ copZ gene/ heavy metal resistance/ silver ion.
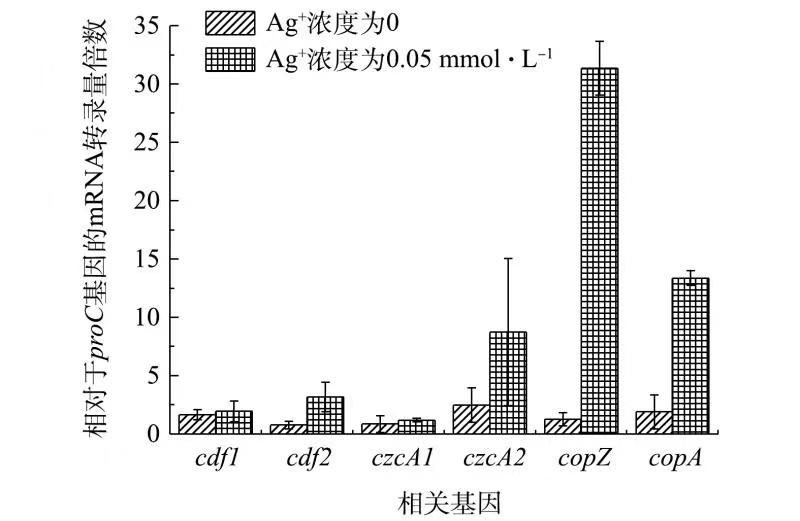
图1不添加和添加Ag+条件下G. sulfurreducens重金属转运相关基因表达变化对比 Figure1.Comparison of the expression changes of heavy metal transport related genes in G. sulfurreducens in the absence or presence of Ag+
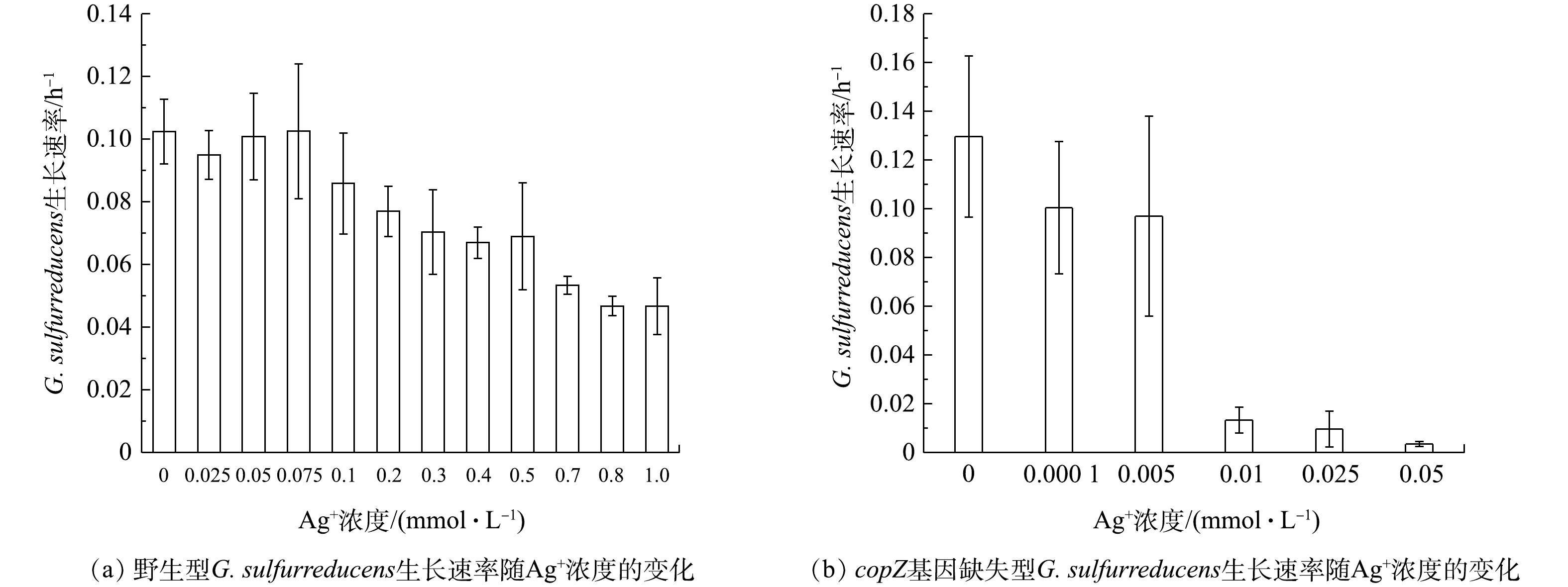
图2野生型与copZ基因缺失型G. sulfurreducens菌株对Ag+的耐受浓度阈值 Figure2.Tolerance threshold of wild-type and copZ-deficient G. sulfurreducens strains to Ag+ concentration
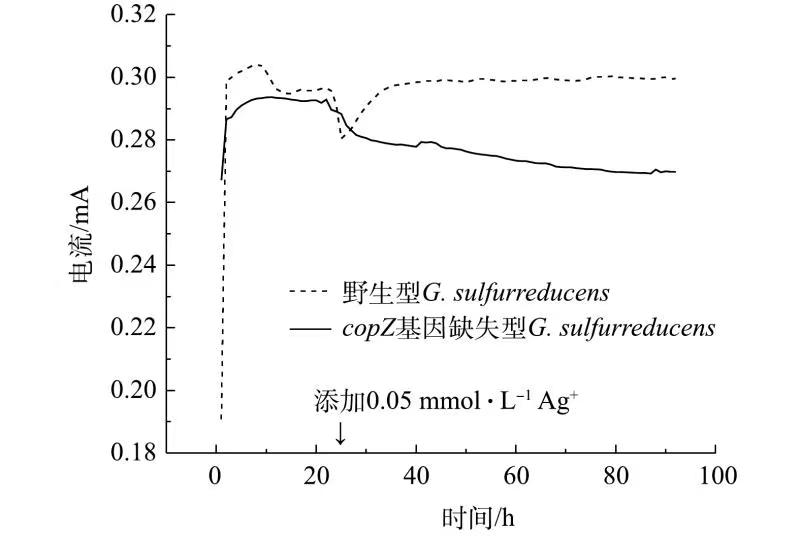
图3Ag+对分别接种了野生型与copZ基因缺失型G. sulfurreducens菌株的MFC系统的产电性能的影响 Figure3.Effect of Ag+ on the electricity production performance of MFC systems inoculated with wild-type and copZ-deficient G. sulfurreducens strains
LOVLEY D R, UEKI T, TIAN Z, et al. Geobacter: The microbe electric's physiology, ecology, and practical applications[J]. Advances in Microbial Physiology, 2011, 59: 1-100.
[2]
BURKHARDT E M, BISCHOFF S, AKOB D M, et al. Heavy mental tolerance of Fe(Ⅲ)-reducing microbial communities in contaminated creek bank soils[J]. Applied & Environmental Microbiology, 2011, 77(9): 3132.
WANG Z, CHEN J G, LI X H, et al. Aquatic toxicity of nanosilver colloids to different trophic organisms: Contributions of particles and free silver ion[J]. Environmental Toxicology & Chemistry, 2012, 31(10): 2408-2413.
[7]
MIAO A J, SCHWEHR K A, XU C, et al. The algal toxicity of silver engineered nanoparticles and detoxification by exopolymeric substances[J]. Environmental Pollution, 2009, 157(11): 3034-3041. doi: 10.1016/j.envpol.2009.05.047
[8]
BONDARCZUK K, PIOTROWSKA-SEGET Z. Molecular basis of active copper resistance mechanisms in gram-negative bacteria[J]. Cell Biology and Toxicology, 2013, 29(6): 397-405. doi: 10.1007/s10565-013-9262-1
[9]
COPPI M V, LEANG C, SANDLER S J, et al. Development of a genetic system for Geobacter sulfurreducens[J]. Applied & Environmental Microbiology, 2001, 67(7): 3180.
[10]
MOUSER P J, HOLMES D E, PERPETUA L A, et al. Quantifying expression of Geobacter spp. oxidative stress genes in pure culture and during in situ uranium bioremediation[J]. The ISME Journal, 2009, 3(4): 454-465. doi: 10.1038/ismej.2008.126
[11]
AKLUJKAR M, LOVLEY D R. Interference with histidyl-tRNA synthetase by a CRISPR spacer sequence as a factor in the evolution of Pelobacter carbinolicus[J]. BMC Evolutionary Biology, 2010, 10(1): 230. doi: 10.1186/1471-2148-10-230
[12]
ODERMATT A, SUTER H, KRAPF R, et al. Primary structure of two P-type ATPases involved in copper homeostasis in Enterocuccus hirae[J]. Journal of Biological Chemistry, 1993, 268(17): 12775-12779. doi: 10.1016/S0021-9258(18)31455-8
[13]
BANCI L, BERTINI I, CIOFI-BAFFONI S, et al. Understanding copper trafficking in bacteria: interaction between the copper transport protein CopZ and the N-terminal domain of the copper ATPase CopA from Bacillus subtilis[J]. Biochemistry, 2003, 42(7): 1939. doi: 10.1021/bi027096p
[14]
HWANG M G, KATAYAMA H, OHAGKI S, et al. Inactivation of Legionella pneumophila and Pseudomonas aeruginosa: Evaluation of the bactericidal ability of silver cations[J]. Water Research, 2007, 41(18): 4097-4104. doi: 10.1016/j.watres.2007.05.052
[15]
RAIMUNDA D, GONZáLEZ-GUERRERO M, LEEBER B W, et al. The transport mechanism of bacterial Cu+-ATPases: Distinct efflux rates adapted to different function[J]. BioMetals, 2011, 24(3): 467-475. doi: 10.1007/s10534-010-9404-3
Engineering Research Center for Water Pollution Source Control and Eco-Remediation, Beijing Key Laboratory for Source Control Technology of Water Pollution, College of Environmental Science and Engineering, Beijing Forestry University, Beijing 100083, China Received Date: 2020-08-28 Accepted Date: 2021-01-26 Available Online: 2021-06-25 Keywords:microbial fuel cell/ Geobacter sulfurreducens/ copZ gene/ heavy metal resistance/ silver ion Abstract:Microbial fuel cell (MFC) is a hot spot research area in sewage treatment. Geobacter is widely recognized in the field of MFC due to its excellent electricity generation ability. The biologically toxic components such as heavy metals in the natural water environment will affect the growth and survival of Geobacter and the ability to generate electricity, which in turn affects the electricity generation performance of MFC. Geobacter is generally tolerant to various kinds of heavy metals such as Ag+. However, the mechanism of its tolerance to higher concentrations of heavy metals is still unclear. In this study, Geobacter sulfurreducens, a model species of Geobacter, was selected to study the regulation of the copZ gene on G. sulfurreducens’s tolerance to Ag+. Results showed that the transcription of the copZ gene of wild-type G. sulfurreducens increased by 24.8 times under 0.05 mmol·L?1Ag+. When copZ was knocked out from the G. sulfurreducens genome, the tolerance of G. sulfurreducens to Ag+ decreased significantly. The growth rate of the copZ-deficient G. sulfurreducens strain in the presence of 0.01 mmol·L?1 Ag+ was only 33.3% of that of the wild-type G. sulfurreducens in the presence of 1 mmol·L?1Ag+. When 0.05 mmol·L?1 Ag+ was added to the microbial fuel cell (MFC) system which inoculated with the copZ -deficient G. sulfurreducens strain, the output current of the MFC decreased by 6.99%. This study proved that the copZ has a significant regulatory effect on G. sulfurreducens tolerance to Ag+, and revealed the internal mechanism of high Ag+ concentration influence on the electricity productivity performance in the G. sulfurreducens MFC system.
本研究中所用菌株为实验室收集保存的野生型Geobacter sulfurreducens DL1(ATCC 51573),在含有10 mmol·L?1的乙酸作为唯一电子供体和50 mmol·L?1的富马酸作为唯一电子受体的厌氧培养基中恒温(30 °C)培养。 培养基的制备:每升去离子水中加入0.42 g KH2PO4、0.22 g K2HPO4、0.2 g NH4Cl、0.38 g KCl、0.36 g NaCl、0.04 g CaCl2·2H2O、0.10 g MgSO4·7H2O、1.80 g NaHCO3,0.50 g Na2CO3,2.04 g NaC2H3O2·3H2O、6.40 g Na2C4H4O4,再加入0.50 mL质量分数为0.1%刃天青,1 mL浓度为100 mmol·L?1的Na2SeO4溶液,10 mL 维生素溶液以及10 mL 微量矿物质溶液。维生素溶液成分参考文献[9]。微量矿物质溶液的制备:每升去离子水中添加0.10 g MnCl2·4H2O、0.3 g FeSO4·7H2O、0.17 g CoCl2·6H2O、0.20 g ZnSO4·7H2O、0.30 g CuCl2·2H2O、0.005 g AlK(SO4)2·12H2O、0.005 g H3BO3、0.09 g Na2MoO4、0.11 g NiSO4·6H2O及0.2 g Na2WO4·2H2O。 Ag+以硝酸银(AgNO3)形式添加,浓度为0.000 1~1 mmol·L?1。在厌氧条件下,用一次性无菌注射器从接种培养后的培养基中取出1 mL菌液至一次性比色皿中,在可见分光光度计600 nm波长下检测菌液的OD值,再根据培养时间换算出生长速率[10]。
1.2. 定量反转录PCR分析相关基因转录量的方法
用上述培养基培养野生型G. sulurreducens。培养条件分别为菌液中不加Ag+和添加0.05 mmol·L?1 的Ag+。培养至指数增长期后,使用RNEasy Plus minikit(Qiagen)提取RNA,并用不含DNA的DNase(Ambion)处理。RNA样品纯度通过琼脂糖凝胶电泳进行检查,A260/A280的比率为1.8~2.0,并用相关基因引物以提取的RNA样品为模版进行PCR扩增,确保RNA样品不含DNA污染。最后,用Superscript III first-strand synthesis SuperMix(Invitrogen)试剂盒将纯化后的RNA样品反转录为cDNA。 从美国能源部联合基因组研究所网站(www.jgi.doe.gov)获得基因组序列数据用于设计定量反转录PCR(qRT-PCR)引物,主要关注的基因包括助阳离子扩散体(CDF)家族的cdf1(GSU0487)、cdf2(GSU2613),抗瘤细胞分裂蛋白(RND)家族的czcA1(GSU3400)、czcA2(GSU0830)和P型ATP酶(P-type ATPase)家族的copZ(GSU1338)、copA(GSU2452)(见表1)。然后,以上述反转录的cDNA为模版,用7500 qPCR系统(Applied Biosystems)进行qRT-PCR扩增和检测,并用已知浓度的纯化cDNA做系列稀释,构建覆盖6个数量级的标准曲线。
反应体系由2组双极室MFC组成,阴阳两极室的容积均为250 mL,并由质子膜(CMI;CMI7000,Membranes International Inc.,美国)隔开。每升阴阳极溶液含0.42 g KH2PO4、0.22 g K2HPO4、0.20 g NH4Cl2、0.38 g KCl、0.36 g NaCl、0.04 g CaCl2·2H2O、0.10 g MgSO4·7H2O、1.80 g NaHCO3、0.50 g Na2CO3,以及1 mmol·L?1的Na2SeO4溶液10 mL、10 mL 微量矿物质溶液和15 mL 维生素溶液[9]。阳极室培养液中还加入10 mmol·L?1乙酸作为唯一电子供体。配制时将以上组分溶解于800 mL去离子水中,溶解后定容至1 L,各取200 mL溶液并用N2∶CO2=80∶20的混合气曝气30 min。阳极室用丁腈橡胶塞密封以保证厌氧环境,并利用磁力搅拌器进行搅拌。阴阳极均为石墨棒电极(Φ 6 mm×80 mm),参比电极为饱和甘汞电极(相对标准氢电势为+199 mV)。分别在2组MFC的阳极室接种野生型G. sulfurreducens和copZ基因缺失型G. sulurreducens。通过电化学工作站(ChI1030C,上海辰华仪器有限公司)控制阳极电势为+300 mV,同时监测2组MFC的输出电流。
 下载:
下载:  点击查看大图
点击查看大图