Remediation technology of submerged plants in Baiyangdian Lake based on light compensation depth
YAN Jun1,2,, WANG Ying2, WANG Yibo3, WANG Jinggang1,,, YANG Wen2, CHEN Zehao2, GUO Siya1 1.School of Chemical Engineering, Beijing University of Chemical Technology, Beijing 100029, China 2.School of Environment, Beijing Normal University, Beijing 100875, China 3.Grass Industry College, Gansu Agricultural University, Lanzhou 730070, China
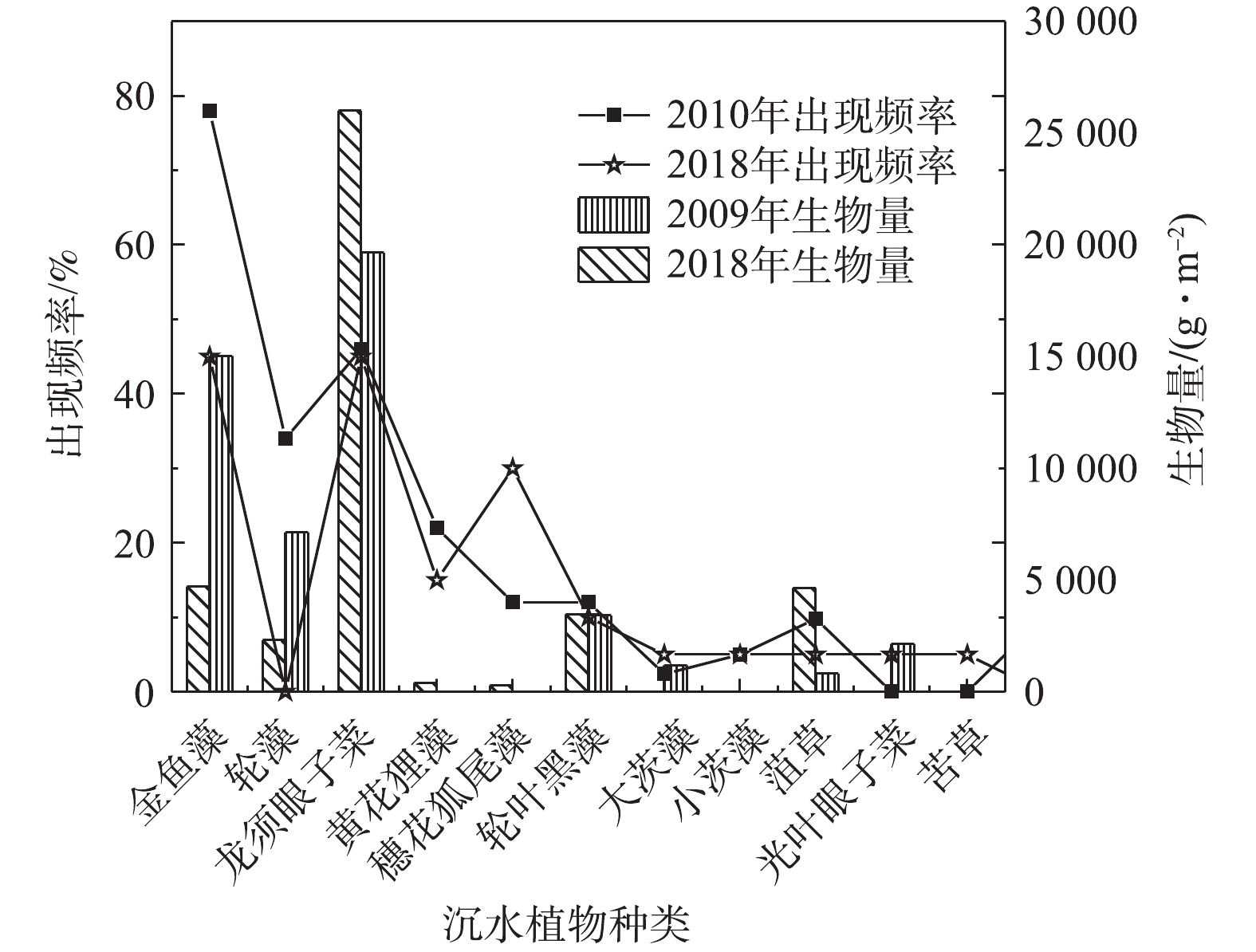
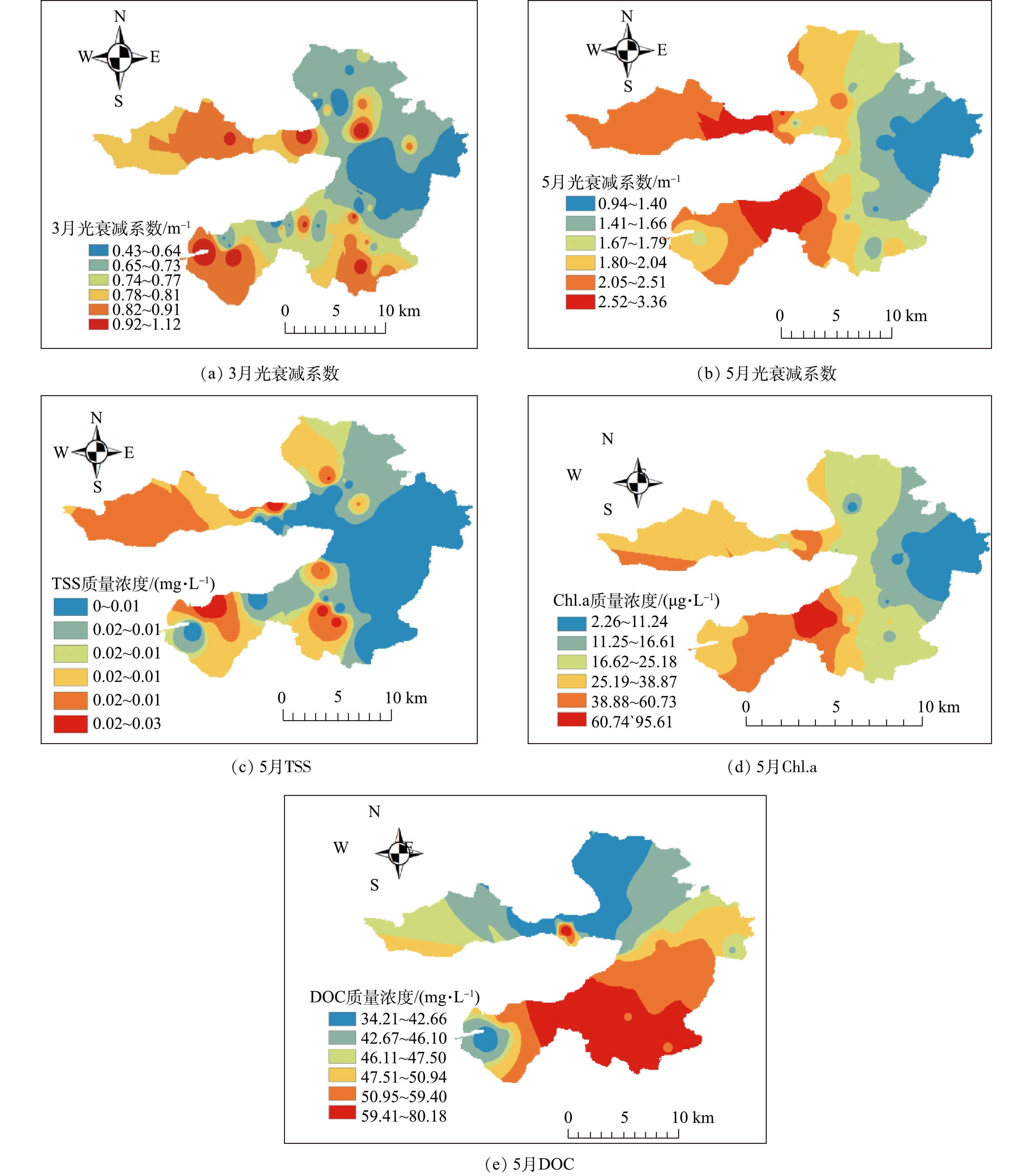
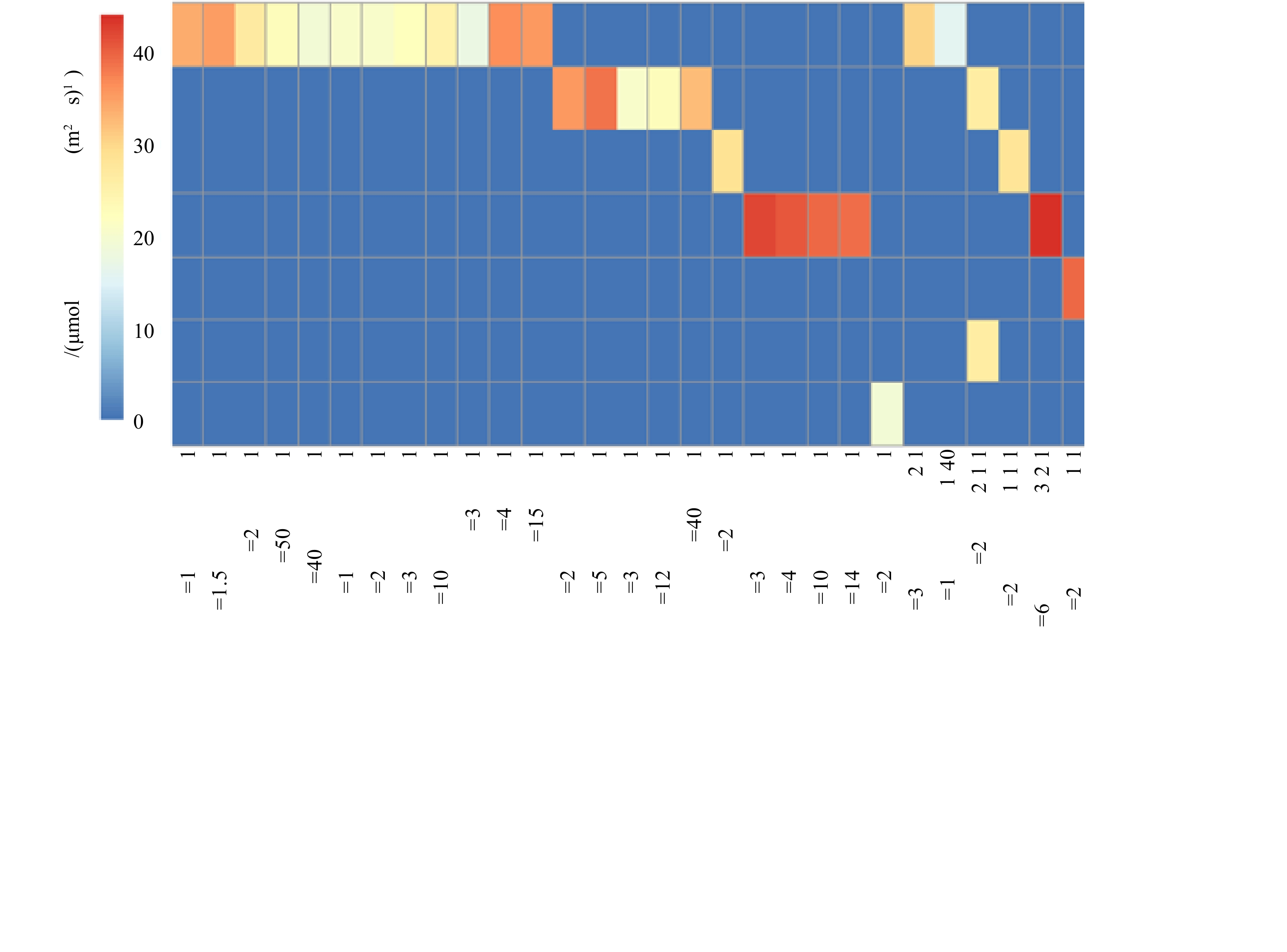
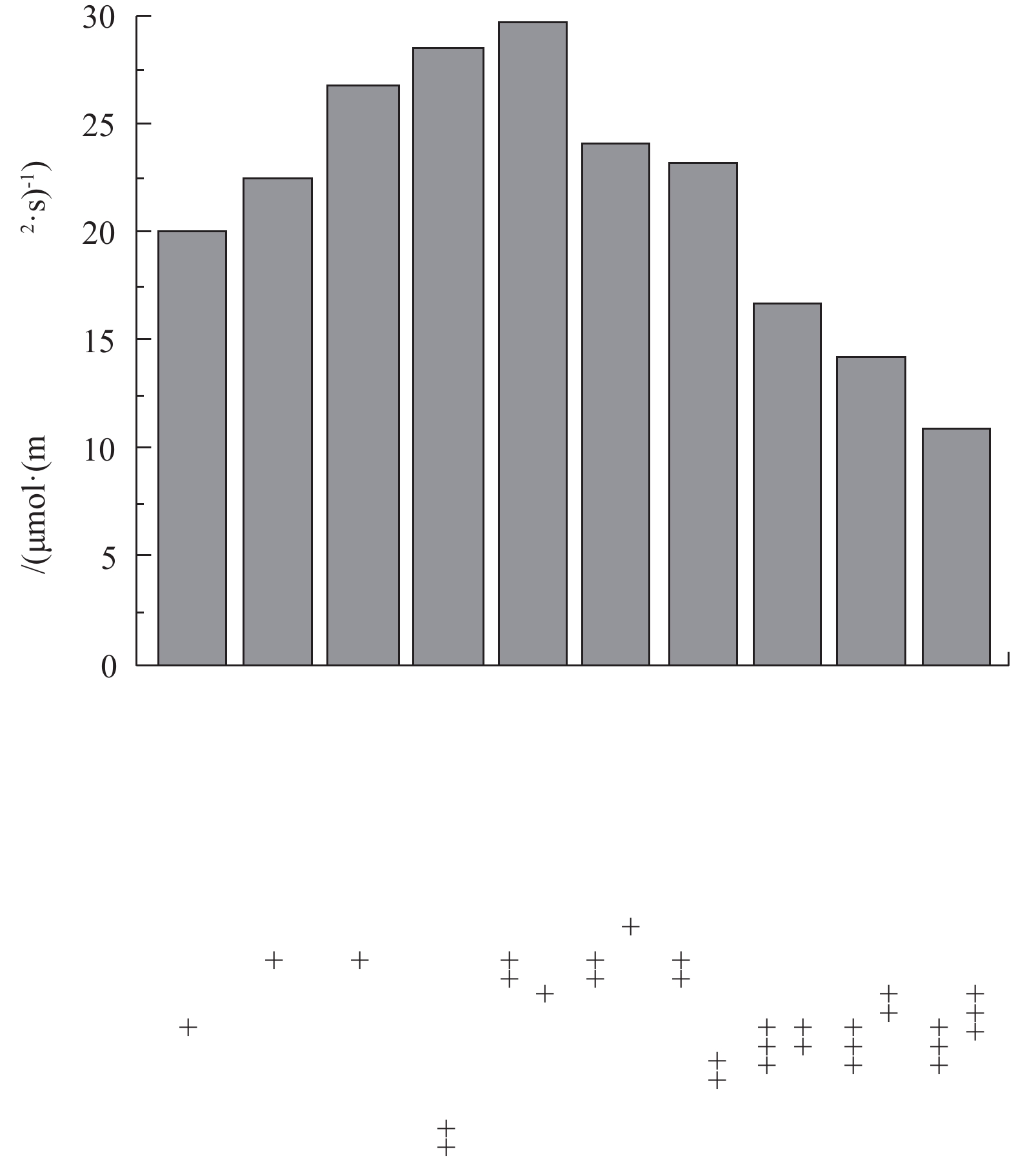
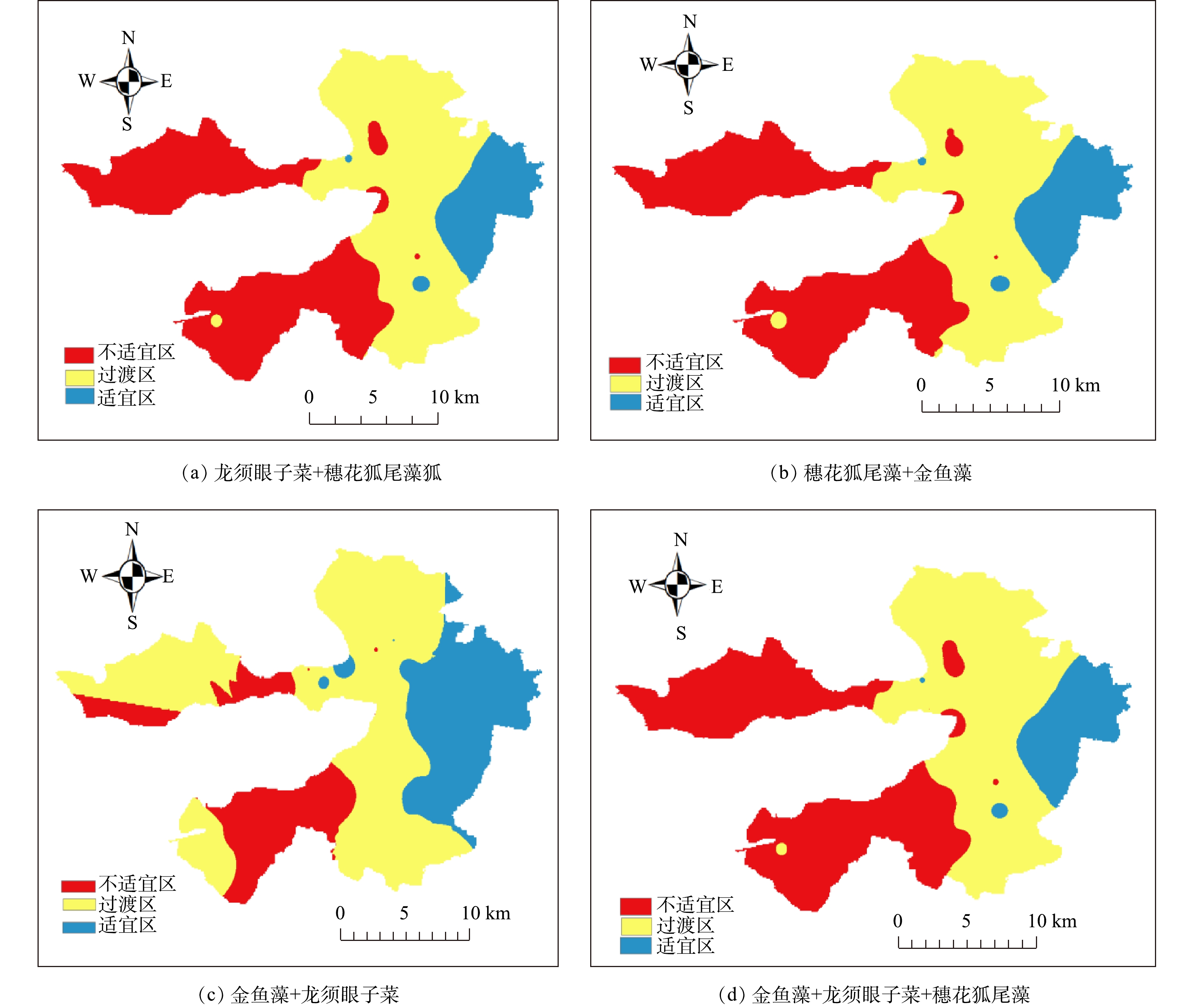
Abstract:Submerged plants are the key to the ecological restoration of shallow lakes. In this study, submerged plants in Baiyangdian Lake area were sampled in 2018 and compared with its history. The changes of the underwater light field were also analyzed in March and May 2019. Considering the underwater light field, anti-pollution and light sensitivity of submerged plants, a two-phase restoration plan for submerged plant communities (stain-resistant + medium-tolerant and sensitive species) was determined. The results showed that the submerged plant community surveyed in the spring, summer and autumn of 2018 were taken as research target, the correlation between the measured light compensation point of the community and its composition and the dominant species were determined, and the plant proportion has slight effect on the light compensation point of its community. For example, the combination of P.pectinatus +Ceratophyllum, the light intensities of the community light compensation point with P.pectinatus as the dominant species (2∶1 and 50∶1) were 22.46 μmol·(m2·s)?1 and 26.05 μmol·(m2·s)?1, respectively. The light intensities of the community light compensation point with Ceratophyllum as the dominant species (1∶2 and 1∶5) were 35.01 μmol· (m2·s)?1 and 37.79 μmol ·(m2·s)?1, respectively. The results of the underwater light field showed the light attenuation coefficient had a trend of low values in the east and high values in the west. This spatial variation trend was similar to that of chlorophyll a and total suspended matter. Of which, the effect of chlorophyll was greater. The high-value regions had more human interference, while the low-value regions had less human interference. The depth of light compensation and the transparency of submerged plant populations (communities) showed significant linear positive correlation (most R2 values were higher than 0.74). The ratio of light compensation depth of submerged plant population (community) to actual water depth was used to evaluate the first stage of Baiyangdian restoration area, and through the correlation analysis of early light attenuation coefficient with chlorophyll a, suspended matter and DOC, it was determined that chlorophyll a and suspended matter content in the water body could be reduced, so as to increase the depth of light compensation of submerged plants, and then achieve the restoration of submerged plant communities. The above research results can provide a reference for the ecological management of Baiyangdian and northern lakes. Key words:optical compensation depth/ staged ecological restoration/ Baiyangdian Lake/ submerged plants.
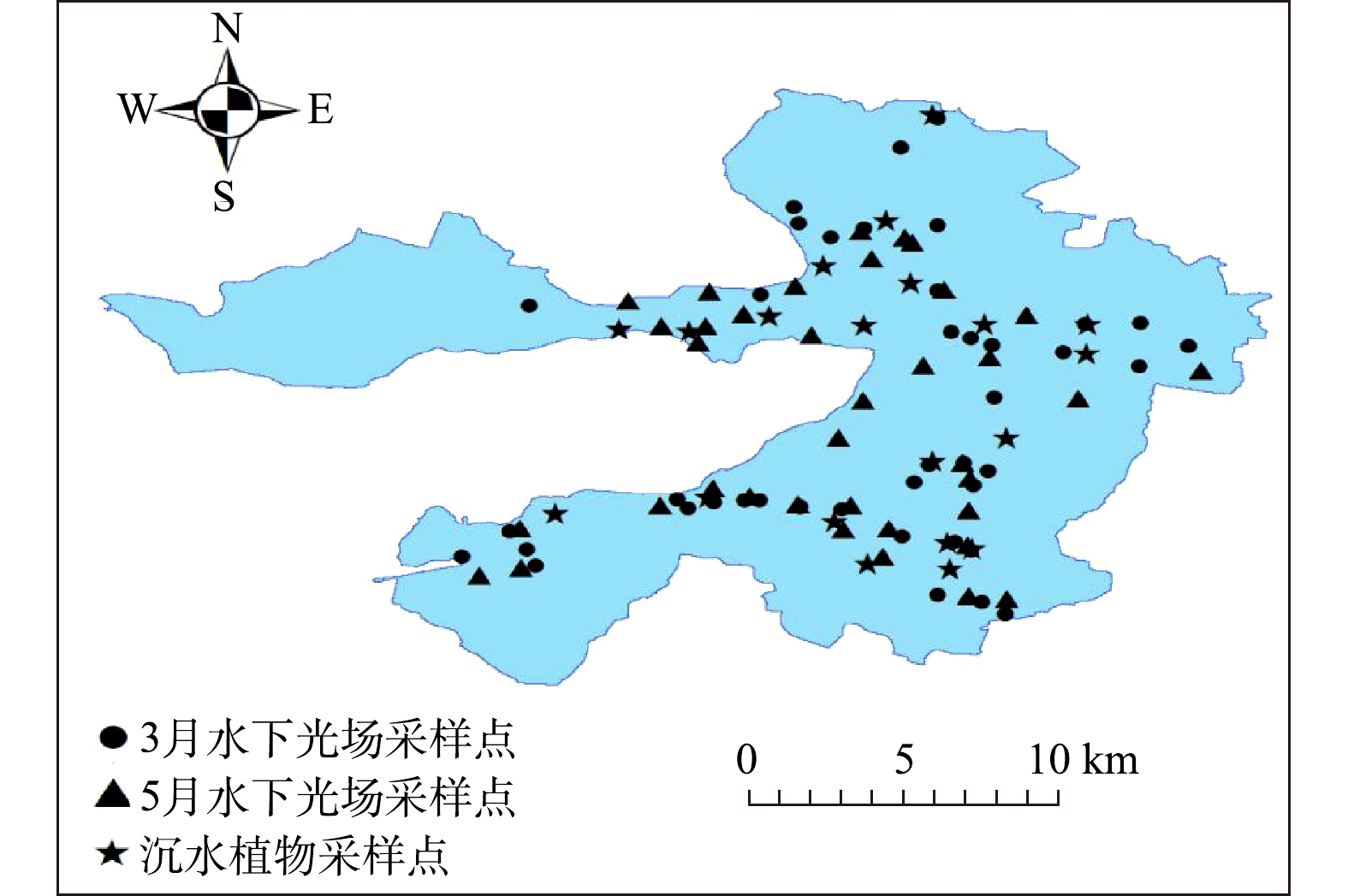
图1白洋淀采样点的分布 Figure1.Distribution of sampling points in Baiyangdian
DONK E V, BUND W J V D. Impact of submerged macrophytes including charophytes on phyto- and zooplankton communities: Allelopathy versus other mechanisms[J]. Aquatic Botany, 2002, 72(1/2/3/4): 274.
[18]
ELLER F, ALNOEE A B, BODERSKOV T, et al. Invasive submerged freshwater macrophytes are more plastic in their response to light intensity than to the availability of free CO2, in air: Equilibrated water[J]. Freshwater Biology, 2015, 60(5): 929-943. doi: 10.1111/fwb.12547
[19]
SQUIRES M M, LESACK L F W, HUEBERT D. The influence of water transparency on the distribution and abundance of macrophytes among lakes of the Mackenzie Delta, Western Canadian Arctic[J]. Freshwater Biology, 2002, 47(11): 2123-2135. doi: 10.1046/j.1365-2427.2002.00959.x
[20]
SNDERGAARD M, PHILLIPS G, HELLSTEN S, et al. Maximum growing depth of submerged macrophytes in European lakes[J]. Alessandro Oggioni. Hydrobiologia, 2013, 704(1): 165-177. doi: 10.1007/s10750-012-1389-1
[21]
SCNDERGAARD M, JOHANSSON L S, LAURIDSEN T L, et al. Submerged macrophytes as indicators of the ecological quality of lakes[J]. Freshwater Biology, 2010, 55(4): 893-908. doi: 10.1111/j.1365-2427.2009.02331.x
[22]
NURMINEN L, HORPPILA J. Life form dependent impacts of macrophyte vegetation on the ratio of resuspended nutrients[J]. Water Research, 2009, 43(13): 3217-3226. doi: 10.1016/j.watres.2009.04.041
[23]
LI E H, LI W, LIU G H, et al. The effect of different submerged macrophyte species and biomass on sediment resuspension in a shallow freshwater lake[J]. Aquatic Botany, 2008, 88(2): 126.
[24]
IBELINGS B W, PORTIELJE R, LAMMENS E H R R, et al. Resilience of alternative stable states during the recovery of shallow lakes from eutrophication: Lake veluwe as a case study[J]. Ecosystems, 2007, 10(1): 4-16. doi: 10.1007/s10021-006-9009-4
[25]
BACHMANN R W, HOYER M V, CANFIELD D E. Evaluation of recent limnological changes at Lake Apopka[J]. Hydrobiologia, 2001, 448(1/2/3): 19-26.
[26]
SCHREIBER U, KLUGHAMMER C, JRG K. Assessment of wavelength-dependent parameters of photosynthetic electron transport with a new type of multi-color PAM chlorophyll fluorometer[J]. Photosynthesis Research, 2012, 113(1/2/3): 127-144.
[27]
ASAEDA T, SULTANA M, MANATUNGE J, et al. The effect of epiphytic algae on the growth and production of Potamogeton perfoliatus L. in two light conditions[J]. Environmental and Experimental Botany, 2004, 52(3): 225-238. doi: 10.1016/j.envexpbot.2004.02.001
[28]
LUND-HANSEN L C. Diffuse attenuation coefficients Kd(PAR) at the estuarine North Sea Baltic Sea transition: Time-series, partitioning, absorption, and scattering[J]. Estuarine Coastal & Shelf Science, 2004, 61(2): 251-259.
[29]
KEMP W M, BATLESON R, BERGSTROM P, et al. Habitat requirements for submerged aquatic vegetation in chesapeake bay: Water quality, light regime, and physical-chemical factors[J]. Estuaries and Coasts, 2004, 27(3): 363-377. doi: 10.1007/BF02803529
[30]
KURTZ J C, YATES D F, MACAULEY J M, et al. Effects of light reduction on growth of the submerged macrophyte Vallisneria americana and the community of root-associated heterotrophic bacteria[J]. Journal of Experimental Marine Biology and Ecology, 2003, 291(2): 199-218. doi: 10.1016/S0022-0981(03)00120-5
[31]
FRENETTE J J, ARTS M T, MORIN J. Spectral gradients of downwelling light in a fluvial lake (Lake Saint-Pierre, St-Lawrence River)[J]. Aquatic Ecology, 2003, 37(1): 77-85. doi: 10.1023/A:1022133530244
1.School of Chemical Engineering, Beijing University of Chemical Technology, Beijing 100029, China 2.School of Environment, Beijing Normal University, Beijing 100875, China 3.Grass Industry College, Gansu Agricultural University, Lanzhou 730070, China Received Date: 2019-11-18 Accepted Date: 2020-05-02 Available Online: 2020-10-14 Keywords:optical compensation depth/ staged ecological restoration/ Baiyangdian Lake/ submerged plants Abstract:Submerged plants are the key to the ecological restoration of shallow lakes. In this study, submerged plants in Baiyangdian Lake area were sampled in 2018 and compared with its history. The changes of the underwater light field were also analyzed in March and May 2019. Considering the underwater light field, anti-pollution and light sensitivity of submerged plants, a two-phase restoration plan for submerged plant communities (stain-resistant + medium-tolerant and sensitive species) was determined. The results showed that the submerged plant community surveyed in the spring, summer and autumn of 2018 were taken as research target, the correlation between the measured light compensation point of the community and its composition and the dominant species were determined, and the plant proportion has slight effect on the light compensation point of its community. For example, the combination of P.pectinatus +Ceratophyllum, the light intensities of the community light compensation point with P.pectinatus as the dominant species (2∶1 and 50∶1) were 22.46 μmol·(m2·s)?1 and 26.05 μmol·(m2·s)?1, respectively. The light intensities of the community light compensation point with Ceratophyllum as the dominant species (1∶2 and 1∶5) were 35.01 μmol· (m2·s)?1 and 37.79 μmol ·(m2·s)?1, respectively. The results of the underwater light field showed the light attenuation coefficient had a trend of low values in the east and high values in the west. This spatial variation trend was similar to that of chlorophyll a and total suspended matter. Of which, the effect of chlorophyll was greater. The high-value regions had more human interference, while the low-value regions had less human interference. The depth of light compensation and the transparency of submerged plant populations (communities) showed significant linear positive correlation (most R2 values were higher than 0.74). The ratio of light compensation depth of submerged plant population (community) to actual water depth was used to evaluate the first stage of Baiyangdian restoration area, and through the correlation analysis of early light attenuation coefficient with chlorophyll a, suspended matter and DOC, it was determined that chlorophyll a and suspended matter content in the water body could be reduced, so as to increase the depth of light compensation of submerged plants, and then achieve the restoration of submerged plant communities. The above research results can provide a reference for the ecological management of Baiyangdian and northern lakes.
 下载:
下载:  点击查看大图
点击查看大图