Inheritance and breeding of japonica rice with good eating quality in Jiangsu province
Cailin Wang, Yadong Zhang, Chunfang Zhao, Xiaodong Wei, Shu Yao, Lihui Zhou, Zhen Zhu, Tao Chen, Qingyong Zhao, Ling Zhao, Kai Lu, Wenhua LiangInstitute of Food Crops, Jiangsu Academy of Agricultural Sciences, Jiangsu High Quality Rice Risearch and Development Center, Nanjing Branch of China National Center for Rice Improvement, Nanjing 210014, China责任编辑: 王秀娥
收稿日期:2020-12-31修回日期:2021-03-1网络出版日期:2021-05-20
| 基金资助: |
Received:2020-12-31Revised:2021-03-1Online:2021-05-20
| Fund supported: |
作者简介 About authors
王才林,博士,研究员,水稻遗传育种。E-mail:

摘要
为了培育适合在江苏种植的优良食味粳稻品种,以优良食味为主攻目标,对江苏省优质、抗病、高产粳稻品种的遗传基础开展了系统研究。通过关联分析,研究稻米蒸煮品质、营养品质与食味品质的关系,明确直链淀粉含量是影响食味品质的关键性状。直链淀粉含量为10%~14%的半糯米米饭表面光泽透亮、质地柔软、富有弹性,综合了糯米的柔软性和粳米的弹性,冷饭不硬,食味品质极佳,符合长三角地区人们喜食柔软香型粳米的口感需求。筛选出直链淀粉含量在10%左右、含有低直链淀粉含量基因Wx mp的日本半糯粳稻品种“关东194”作为食味品质改良的核心种质。开发Wx mp基因的分子标记,结合利用与抗病、高产相关基因紧密连锁的分子标记,开展优良食味、抗病、高产粳稻聚合育种。育成适合江苏省不同稻区种植的南粳46、南粳5055、南粳9108和南粳5718等优良食味粳稻系列新品种通过审定,形成了覆盖江苏省不同稻区的优良食味粳稻品种布局,累计推广面积超过533万公顷,有力地促进了江苏省及周边地区优质稻米产业的发展,为水稻产业供给侧结构调整,提质增效,保障粮食安全作出了重要贡献。
关键词:
Abstract
In order to develop a variety of japonica rice with good eating quality suitable for planting in Jiangsu Province, the genetic basis of high quality, disease resistance and high yield japonica rice varieties in Jiangsu Province was systematically studied. The relationship among different rice qualities of cooking, nutrition, and eating was studied by association analysis. It was clear that amylose content was the key factor affecting eating quality. The semi waxy rice with amylose content of 10%~14% has bright surface, soft texture, and elasticity, combining the softness of glutinous rice and the elasticity of japonica rice. The cold rice is not hard, and the taste is excellent. It meets the taste requirements of people in Yangtze River Delta region who like to eat soft fragrant japonica rice. The semi waxy japonica rice variety “Kantou 194” with a low expression of amylose content gene Wx mp and an amylose content of about 10% was selected as the core germplasm for improving eating quality. Pyramiding breeding of japonica rice variety with good eating quality, disease resistance and high yield was carried out by examining the development of Wx mp gene molecular markers and the use of closely linked molecular markers with disease resistance and high yield genes. A series of new japonica rice varieties with good taste such as Nanjing 46, Nanjing 5055, Nanjing 9108, and Nanjing 5718, suitable for different rice areas of Jiangsu Province, have been bred and approved by Jiangsu Provincial Variety Approval Committee. The layout of japonica rice varieties with good taste covering different rice areas in Jiangsu Province has been formed. These varieties have been planted with an accumulated area of more than 5.3 million hectares, which has effectively promoted the development of high quality rice industry in Jiangsu Province and its surrounding areas, and made important contributions to the structural adjustment of the supply side of rice industry, improving quality and efficiency, and ensuring food security.
Keywords:
PDF (979KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
王才林, 张亚东, 赵春芳, 魏晓东, 姚姝, 周丽慧, 朱镇, 陈涛, 赵庆勇, 赵凌, 路凯, 梁文化. 江苏省优良食味粳稻的遗传与育种研究. 遗传[J], 2021, 43(5): 442-458 doi:10.16288/j.yczz.20-452
Cailin Wang.
水稻是我国最重要的粮食作物,提高水稻产量、改善稻米品质是保障中国口粮绝对安全、提高人民生活水平的重要举措[1]。近几年,江苏省水稻种植面积稳定在220万公顷左右,其中粳稻面积近200万公顷,总产约1700万吨,分别占全国粳稻面积和总产的22%和24%左右,在保障江苏乃至全国粮食安全中的地位举足轻重[2]。因此,上个世纪,江苏省的水稻育种一直以高产为主要目标。
进入21世纪以后,随着人民生活水平的提高,稻米品质越来越受到消费者的重视,水稻产业实行大力发展优质稻的战略性调整,优质成为江苏省粳稻育种的首要目标[3]。同时条纹叶枯病开始在江苏粳稻区爆发流行,对江苏粳稻育种提出严峻挑战。而当时江苏水稻生产上大面积推广的高产粳稻品种往往高而不优、优而不抗。日本品种“越光”的食味品质堪称一绝,黑龙江五常生产的“稻花香2号”在国内也有较高的知名度,但日本和东北品种均属早中粳类型,且不抗条纹叶枯病,产量较低,不适合在江苏大面积种植。因此,江苏粳稻的优质育种必须以高产和抗病为基础。
稻米品质包括加工品质、外观品质、食味品质、营养品质和安全品质[4]。食味品质指稻米在蒸煮和食用过程中所表现的各种理化及感官特性,是稻米品质的核心。优良食味就是食味好、好吃的意思,优良食味粳稻就是指好吃的粳稻品种。然而,食味品质究竟与哪些因素有关?中国地域广大,水稻品种类型多样,不同地区人群对稻米品质的要求及嗜好性各不相同,作为南方粳稻主栽区的江苏,食味品质改良的路径该如何选择?近20年来,本研究团队以优良食味为主攻目标,对江苏省优质、抗病、高产粳稻品种选育的技术途径开展了系统研究。本文总结本团队在粳稻食味品质性状的遗传和改良方面取得的成绩,以期为育种工作者提供参考。
1 稻米食味品质主要基因的遗传变异及其对食味品质的影响
1.1 粳稻食味品质相关基因的关联分析
为了阐明影响稻米食味品质的遗传基础,对253个水稻品种(包括83个籼稻和170个粳稻品种)进行了全基因组关联分析。通过群体结构分析,发现籼粳两个亚种存在较强的群体分化(Fst>0.75),因而对籼、粳两个亚种分别进行了关联分析。结果表明,在整个群体和籼粳两个亚种中,多数性状间存在显著的表型相关。在籼稻和粳稻亚种分别检测到33个和30个遗传位点。大多数位点与淀粉合成相关基因重叠,并在Wx基因上检测到控制直链淀粉含量(amylose content, AC)、胶稠度(gel consistency, GC)和RVA谱特征值等9个性状的14个遗传位点,表明该位点是一个聚集了多个性状的多效性基因位点。8个遗传位点表现出籼粳亚种的特异性,包括籼稻亚种特有的RM588、Wx-(CT)n、SSI和SBE1遗传位点,粳稻亚种特有的RM550、Wxmp、SSIIa和SBE4遗传位点,表明控制食味品质性状的遗传位点存在籼粳亚种间的分化。在粳稻亚种两年都检测到位于第2染色体上的一个SSR位点RM550,是一个潜在的影响AC和糊化特性的稳定表达的新位点,可后续用于候选基因的精细定位[5]。这些结果表明稻米食味品质等复杂性状存在大量微效遗传位点,这些位点为分子标记辅助选择改良水稻品种的食味品质提供了重要基因资源。1.2 Wx基因的遗传变异及其对食味品质的影响
稻米中70%左右是淀粉,包括直链淀粉和支链淀粉两类,还有不到10%的蛋白质、14%左右的水分和少量油脂和矿物质等[6]。为了明确影响稻米食味品质的主要因素,本研究团队以中国东北、长江中下游和云贵高原粳稻及日本、韩国的优质粳稻与构建的遗传群体为材料,系统研究稻米加工品质(糙米率、整精米率)、外观品质(垩白粒率、垩白度)、食味品质(AC、GC、RVA 谱)、营养品质(籽粒蛋白质含量、游离氨基酸总含量、蛋白亚基所占百分比)、产量性状(单株穗数、主穗实粒数、结实率、收获指数)、植株形态性状(穗颈粗度、分蘖角、倒1叶宽、倒2叶宽、倒3叶宽、倒1叶基角、倒1叶开角等)与食味的关系,发现AC和蛋白质含量(protein content, PC)与食味的关系最密切,决定系数高达60%以上,明确AC是影响食味品质的关键性状[7,8,9]。AC低,米饭黏软、外观油润、有光泽、冷不回生、适口性好;反之,则米饭质地硬、黏性小、蓬松干燥、光泽差、冷饭容易回生、适口性差[10]。根据前人研究结果,稻米AC主要受Wx基因的控制。目前已经报道的Wx基因位点的自然等位变异有Wxa、Wxb、Wxop、Wxhp、Wxmp、Wxmq、Wmw、Wxlv、Wxla、Wxin和wx等[11,12,13,14]。选用Wx位点不同基因型的籼稻(含Wxa基因,AC多在20%以上)、粳稻(含Wxb基因,AC多在14%~20%之间)、半糯粳稻(含Wx位点突变形成的低直链淀粉含量基因如Wxmp,AC在5%~14%之间)和糯稻(含wx基因,AC在2%以下)品种为材料,研究不同基因型AC的变化规律及其对食味品质的影响,发现低AC的半糯粳稻食味品质更佳。低AC的稻米俗称软米,其AC在5%~14%之间,是介于一般黏米和糯米之间的中间类型,故又称为半糯米。其胚乳外观呈云雾状、乳白色,透明度略差(图1)。但半糯米煮成的米饭表面光泽透亮,质地柔软,富有弹性,综合了糯米的柔软性和粳米的弹性,冷饭不硬,食味品质较佳,符合长江中下游地区喜食软糯粳米居民的饮食需求。
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1Wx基因不同等位变异的直链淀粉含量与外观品质
A:wx,糯米,直链淀粉含量(AC)<2%;B:Wxmp,半糯米,AC 5~14%;C:Wxb,黏米,AC 14~20%;D:Wxa,籼米,AC 20~30%。
Fig. 1Amylose content and appearance quality of rice with different allelic variations of Wx gene
于是,本研究团队收集了已报道的14个低AC的半糯突变体[11,12,13,14],研究不同突变体的外观和食味品质发现,半糯粳稻的食味品质虽然总体上均较好,但半糯粳稻的AC也不是越低越好。AC低于8%,米粒不透明,外观品质变差,口感太软,缺乏弹性;而AC达到12%以上,不仅米饭晶莹剔透,富有弹性,冷而不硬,食味品质更佳,符合长江中下游地区人们喜食软香粳米的口感需求,而且外观品质也较好(图2)。本研究团队从已报道的14个低直链淀粉含量基因中,筛选出直链淀粉含量在10%左右、含有低直链淀粉含量Wxmp的日本半糯粳稻品种“关东194”作为食味品质改良的核心种质[15]。
图2
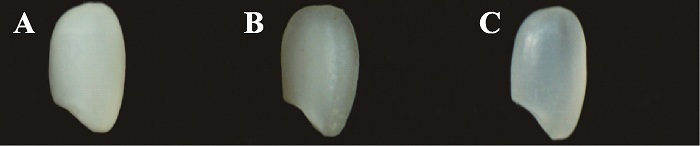 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同直链淀粉含量半糯粳稻米粒的外观品质
A:直链淀粉含量(AC)<8%,米粒不透明;B:AC 10%左右,米粒呈云雾状、半透明;C:AC>12%,米粒透明。
Fig. 2The appearance of semi-waxy rice with different amylose content
1.3 淀粉合成相关基因等位变异及其对食味品质的影响
本研究团队在育种实践中发现,同样半糯基因背景Wxmp的情况下,即使来源于同一杂交组合的不同半糯品系之间,AC的变幅可达5.40%~11.85%[16]。肥料试验和分期播种试验结果表明,穗分化期氮肥用量增加、灌浆初期温度过高或过低均可导致半糯粳稻的AC下降,但下降幅度均在1个百分点以内[17,18,19],表明尚有其他基因影响AC或者基因与环境存在互作。迄今已报道与淀粉合成相关的基因有20多个,其中编码淀粉合成酶的基因除了Wx以外,还有GBSSII、SSI、SSIIa (SSII-3)、SSIIb (SSII-2)、SSIIc (SSII-1)、SSIIIa (SSIII-2)、SSIIIb (SSIII-1)、SSIVa (SSIV-2)和SSIVb (SSIV-1);编码淀粉分支酶的基因有SBE1、SBE3和SBE4,编码去分支酶的基因有ISA1、ISA2、ISA3和PUL[20,21,22]。本研究团队从已报道的20多个淀粉合成相关基因中,筛选出亲本间有差异的淀粉合成酶基因SSIIa、SSIIIa和去分支酶基因PUL,研究这些基因在半糯基因Wxmp背景下的遗传效应。以淀粉合成酶基因SSIIa和SSIIIa表现多态性而其他淀粉合成相关基因无多态性的武粳13与关东194 (Milky Princess)杂交后代衍生的64个半糯品系为材料,分析了Wxmp基因背景下,SSIIa和SSIIIa基因等位变异对AC、GC、糊化温度(gelatinization temperature, GT)及RVA谱特征值的影响。结果表明,SSIIa和SSIIIa等位变异对AC、GC、GT和RVA谱特征值都有显著影响,且两个基因间存在互作效应。SSIIa2和SSIIIa2 (2表示该基因来源于非半糯亲本武粳13)有使AC增高的趋势,分别使AC提高1.87%和1.23%,两年结果基本接近(表1)。单个SSIIa和SSIIIa等位变异对GT无显著影响,而基因型SSIIa1SSIIIa1 (1表示该基因来源于半糯亲本关东194)的GT比SSIIa2SSIIIa2高1.34℃,达到显著水平,表明两个基因的互作对GT有显著影响。GC在不同基因型间均存在极显著差异,SSIIa2和SSIIIa1可分别使GC增加8.74 mm和9.62 mm。从两个基因的互作效应来看,基因型SSIIa2SSIIIa1的GC比基因型SSIIa1SSIIIa2和SSIIa2SSIIIa2分别增加10.64 mm和16.95 mm。SSIIa2使最高黏度、热浆黏度、冷胶黏度、崩解值增加,回复值和消减值下降;而SSIIIa2的效应则相反。两个基因的互作效应,最高黏度、热浆黏度和冷胶黏度均以SSIIa2SSIIIa1最大,崩解值和回复值均以SSIIa2SSIIIa2最大,消减值SSIIa2SSIIIa1最小[23]。
Table 1
表1
表12013年和2014年64个半糯品系SSⅡa和SSⅢa位点不同等位基因对AC的效应
Table 1
| 位点 | 基因型 | 2013年AC | 2014年AC | 平均值 | 基因效应 |
|---|---|---|---|---|---|
| SSⅡa | SSⅡa1 | 9.05 | 8.85 | 8.95 | 1.87 |
| SSⅡa2 | 10.84** | 10.80** | 10.82** | ||
| SSⅢa | SSⅢa1 | 9.37 | 9.17 | 9.27 | 1.23 |
| SSⅢa2 | 10.51** | 10.48** | 10.50** |
新窗口打开|下载CSV
选择在淀粉合成酶基因SSIIa和去分支酶基因PUL存在多态性而其他淀粉合成酶相关基因没有多态性的半糯品系宁0145 (来源于武粳15//武粳13/关东194)和粳稻品种武运粳21进行杂交,获得F2群体与F3株系。利用分子标记,选择含有Wxmp基因的F2单株与F3株系,将这些F2单株与F3株系分成SSIIa1PUL1、SSIIa1PUL2、SSIIa2PUL1和SSIIa2PUL2 4种基因型(1和2分别表示该基因来源于宁0145和武运粳21),分析不同基因型蒸煮食味品质性状的差异,探讨同一Wxmp基因背景下不同SSIIa和PUL等位基因对蒸煮食味品质性状的影响。结果表明,不同基因型间蒸煮食味品质性状均存在显著差异,来源于武运粳21的SSIIa2基因和PUL2基因分别使直链淀粉含量增加0.29%~1.00%和0.62%~1.18%,且PUL的效应大于SSIIa,两者间存在互作效应。SSIIa2基因和PUL2基因对胶稠度和崩解值(breakdown viscosity,BDV)有降低作用,对热浆黏度(hot paste viscosity, HPV)、冷胶黏度(cool paste viscosity, CPV)、消减值(setback viscosity,SBV)和回复值(consistency viscosity,CSV)有升高作用,对GT、最高黏度(peak viscosity, PKV)和峰值时间(peak time, PeT)的作用较小[24]。研究结果明确了Wxmp基因背景下SSIIa和PUL基因对稻米蒸煮食味品质的遗传效应,为SSIIa和PUL基因的分子标记辅助选择改良稻米品质提供了理论依据。上述研究表明,在Wxmp基因背景(AC 10%左右)下,来源于非半糯亲本的SSIIa、SSIIIa和PUL基因可分别使AC提高0.5~1.0个百分点,基因间存在互作效应(表1),阐明了准确筛选AC在10%~14%之间的半糯粳稻基因型的分子机制。为在早期分离世代通过分子标记辅助选择,筛选食味品质优良、外观品质好的理想基因型奠定了理论基础,实现了优良食味粳稻育种理论的新突破。
1.4 香味基因的遗传变异及其对食味品质的影响
米饭的香味也是稻米品质的重要指标。品质好的大米不仅外观晶莹剔透,米饭柔软有弹性,煮饭时还散发诱人香味,品尝时清甜爽口,满口留香。随着香味检测技术的研究与发展,已在不同水稻品种中检测到500多种挥发性芳香物质,其中已经得到鉴定的有200多种[25,26,27],主要包括碳氢化合物、酸类、醇类、醛类、酮类、酯类和酚类等[28]。2-乙酰-1-吡咯啉是香稻中主要香味物质,香稻中爆米花香味主要由2-AP形成[26]。研究表明,稻米的香味是由于水稻第8号染色体上的Badh2基因突变,导致甜菜碱脱氢酶活性丧失,使2-AP大量积累产生香味。而非香水稻中Badh2基因编码的Badh2蛋白具有甜菜碱脱氢酶的活性[29,30,31]。多数研究者认为香味遗传受单个隐性基因的控制[32,33],Badh2基因就是控制稻米中香味的隐性基因[34,35,36,37]。Badh2基因全长1509 bp,包含15个外显子和14个内含子,编码503个氨基酸[35,37]。Badh2基因除在水稻的根中不表达外,在水稻其余组织和器官中均有表达。Badh2基因中至少存在17个变异位点。在香稻品种中,常见的是Badh2基因的第7外显子有1个8 bp的缺失和3个单核苷酸多态性位点[38];在不同的香稻品种中,Badh2基因还在第2外显子有1个7 bp的缺失,第4和第5外显子之间存在1个803 bp的缺失[31,39,40],并且在第1、8、10、12、13和14外显子均存在变异位点[31,33,39,41]。Table 2
表2
表2SSIIa和PUL等位基因蒸煮食味品质性状的遗传效应
Table 2
| 基因型 | F2直链 淀粉含量AC(%) | F3直链 淀粉含量AC(%) | 胶稠度 GC (mm) | 糊化 温度GT(℃) | 最高黏 度PKV (cP) | 热浆黏 度HPV (cP) | 崩解值BDV (cP) | 冷胶黏 度CPV (cP) | 消减值SBV (cP) | 回复值CSV (cP) | 峰值时 间PeT (min) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| SSIIa2-SSIIa1 | 1.01 | 0.29 | -2.79 | 0.12 | 2 | 30 | -28 | 66 | 64 | 36 | 0.07 |
| PUL2-PUL1 | 1.18 | 0.62 | -0.31 | -0.06 | 19 | 89 | -70 | 142 | 123 | 53 | 0.10 |
| SSIIa1PUL2-SSIIa1PUL1 | 1.90 | 0.35 | -6.23 | -0.64 | -44 | 21 | -65 | 115 | 158 | 94 | 0.07 |
| SSIIa2PUL1-SSIIa1PUL1 | 1.83 | 0.01 | -8.80 | -0.43 | -60 | -42 | -18 | 33 | 93 | 74 | 0.05 |
| SSIIa2PUL2-SSIIa2PUL1 | 0.16 | 0.79 | 4.92 | 0.39 | 69 | 140 | -72 | 155 | 86 | 14 | 0.11 |
| SSIIa2PUL2-SSIIa1PUL2 | 0.10 | 0.44 | 2.35 | 0.60 | 52 | 77 | -25 | 72 | 20 | -5 | 0.08 |
| SSIIa2PUL2-SSIIa1PUL1 | 2.00 | 0.80 | -3.88 | -0.04 | 9 | 99 | -90 | 187 | 178 | 89 | 0.15 |
新窗口打开|下载CSV
目前已经针对这些位点设计了一系列分子标记,用于香味基因的鉴定和不同香稻品种的筛选以及香稻新品种的培育。功能标记FMU1-2不但可以检测Badh2基因在第7外显子处的8 bp的缺失突变,而且能够检测Badh2基因的5ʹUTR区是否存在1个8 bp的缺失;功能标记FME2-7、FME7、FME12-3、FME13和FME14分别用于鉴定香味基因Badh2在第2、7、12、13和14外显子处的变异类型[33];标记FMbadh2-E4-5用于检测Badh2基因在第4和5外显子之间是否存在803 bp的缺失[40],YY5-YY8用于检测Badh2基因在第7外显子的8 bp缺失和3个单核苷酸多态性变异[42]。这些水稻香味基因功能标记的开发,为香稻分子育种提供了一种准确、快速且有效的手段。
2 食味品质与产量和抗性的分子标记聚合育种
采用分子生物学和信息学方法,分析发现Wxmp基因与Wx位点上的其他复等位基因间在第4外显子区域存在单碱基的替换,获得与目的基因共分离的分子标记[43]。进一步根据Wxmp基因存在的单核苷酸变异,利用四引物扩增受阻突变体系PCR技术进行等位基因变异扩增,实现了Wxmp基因型的快捷、准确鉴定[44]。利用半糯基因Wxmp的四引物标记与条纹叶枯病抗性基因Stv-bi共分离的SCAR标记以及香味基因Badh2、稻瘟病抗性基因Pi-ta、Pi-b与粒型基因GS3的分子标记,进行辅助选择和聚合育种,将优良食味基因、香味基因、条纹叶枯病抗性基因和稻瘟病抗性基因与高产性状聚合到一起,建立了优质、抗病、高产水稻新品种选育的高效育种技术体系[45,46,47],选择效率提高3倍以上,缩短育种年限2~3年。围绕优良食味、抗病、高产育种目标,通过地理远缘杂交,以食味品质优良、抗条纹叶枯病的日本半糯粳稻“关东194”为核心亲本(父本),与江苏省高产粳稻“武香粳14”和“武粳13”杂交,构建综合性状优良、遗传基础丰富的育种群体,通过南繁北育加速世代进程、低世代外观品种筛选和高世代食味品质筛选、结合分子标记辅助选择,将日本优质粳稻关东194的半糯基因Wxmp和条纹叶枯病抗性基因Stv-bi同时导入江苏省高产粳稻品种武香粳14和武粳13中,育成了分别适合江苏省太湖稻区、沿江沿海和苏南稻区、苏中稻区种植的优良食味粳稻新品种南粳46[48]、南粳5055[49]和南粳9108[50],此后,又育成适合淮北稻区及黄淮海地区种植的优良食味粳稻新品种南粳南粳505[51]、南粳2728[52]、南粳5718[53]、南粳58、南粳9308、南粳66、南粳518以及适合苏南、苏中地区种植的南粳3908[54]、南粳晶谷[55]、南粳56、南粳9036等,分别通过江苏省、上海市、山东省、安徽省审定与认定,形成了覆盖江苏省不同稻区的优良食味粳稻品种布局,有效解决了江苏省缺乏食味品质优良的高产粳稻品种等突出问题,实现了优质与抗病、高产的统一,是江苏省优良食味粳稻新品种选育的重大突破。
3 代表性品种
3.1 南粳46
南粳46是以日本优质粳稻关东194为父本,与江苏优质高产粳稻武香粳14杂交,经数代外观与食味品质筛选和条纹叶枯病抗性分子标记辅助选择育成,2008年通过江苏省审定(苏审稻200814),2009年通过上海市审定(沪农品审水稻(2009)第003号)。全生育期165 d,属中熟晚粳稻类型,适合在江苏省太湖地区东南部和上海中上等肥力条件下种植[48]。该品种植株生长清秀,叶色淡绿,灌浆速度快,熟相较好,株型紧凑,分蘖力中等偏强,穗型较大,直立穗型。2006-2007年参加江苏省区试,平均亩产608.3 kg,比对照增产5.6%,每亩有效穗19.8万穗,每穗实粒数129粒,结实率90%。根据农业农业农村部食品质量检测中心2007年检测,整精米率66.8%,垩白粒率20.0%,垩白度2.4%,达到国标二级优质稻谷标准。南粳46一直被江苏省农业农村厅列为主导品种,其优异的食味品质和良好的丰产性深受老百姓的喜爱,已成为长三角地区高档优质米打造的首选品种,累计推广种植66.7万公顷。众多稻米生产企业以南粳46为原粮进行优质稻米订单生产,每公斤至少加价0.2元收购。不仅是江苏“苏米”省域公用品牌的核心品种,也是南京市“金陵味道”地产优质稻米、江南味道“苏州大米”、“无锡大米”等区域公用品牌订单种植首选的原粮品种,打造了“苏垦”、“道好”、“晶润”等一批南粳46大米品牌在南京、苏州、上海等多地超市销售。为江苏省优质稻米产业发展和农业供给侧结构性改革作出了重要贡献。
3.2 南粳5055
南粳5055是以关东194为父本,与江苏优质高产粳稻武粳13杂交,于2005年育成的优良食味早熟晚粳稻品种,2011年通过江苏省审定(苏审稻201114),2017年通过上海市引种备案(沪引种2016第3号),2018年通过安徽省引种备案(皖引稻2018107),适宜江苏省沿江及苏南地区、上海市和安徽省沿江江南单季晚粳稻地区种植[49]。该品种株型紧凑,长势较旺,分蘖力较强,叶色较深,群体整齐度较好,穗型中等,偏直立,着粒较密,抗倒性强。每亩有效穗20万左右,每穗总粒130~140粒,结实率90%以上,千粒重24~25 g,株高95~100 cm,全生育期160 d左右,比对照武运粳7号早熟2~3 d。2009-2010年参加江苏省区试,两年平均亩产614.5 kg,较对照宁粳1号增产4.8%,两年增产均达极显著水平;2010年生产试验平均亩产637.7 kg,较对照增产6.0%。接种鉴定:感穗颈瘟,中感白叶枯病、纹枯病、条纹叶枯病。根据农业农村部食品质量检测中心2009年检测:整精米率71.4%,垩白粒率10.0%,垩白度0.8%。南粳5055不仅稻米食味品质优,而且产量潜力高,2012年和2013年连续两年通过了农业农村部组织的超级稻测产验收,平均亩产都超过800 kg,2014年被农业农村部认定为超级稻。南粳5055审定以后一直是江苏省主推品种,年种植面积一直保持在10万公顷以上,近几年的年种植面积超过16.7万公顷,累计种植面积超过133.3万公顷。为我省稻米产业提质增效提供了有力支撑。
3.3 南粳9108
南粳9108是以关东194为父本,与江苏优质高产粳稻武香粳14杂交,应用系谱法和分子标记辅助选择,于2009年育成的优良食味迟熟中粳稻品种,2013年通过江苏省审定(苏审稻201306),2017年通过上海市引种认定(沪引种2016第4号),适宜在江苏省苏中及宁镇扬丘陵地区和上海市种植[50]。该品种株型紧凑,长势较旺,分蘖力较强,叶色淡绿,叶姿较挺,抗倒性较强,后期熟相好。每亩有效穗21.2万,穗实粒数125.5粒,结实率94.2%,千粒重26.4 g,株高96.4 cm,全生育期153 d,比淮稻9号早熟1 d;接种鉴定:感穗颈瘟,中感白叶枯病、高感纹枯病,抗条纹叶枯病。2011-2012年参加江苏省区试,两年区试平均亩产644.2 kg,2011年比淮稻9号增产5.2%,增产达极显著水平,2012年比淮稻9号增产3.2%,比镇稻14增产0.1%;2012年生产试验平均亩产652.1 kg,比对照淮稻9号增产7.3%。根据农业农村部食品质量检测中心2012年检测:整精米率71.4%,垩白粒率10.0%,垩白度3.1%。南粳9108也是一个既优质又高产的品种,2015年被农业农村部确认为超级稻。审定以来一直被列入江苏省主推品种,2016年被农业农村部列为长江中下游主导品种,种植面积不断扩大,2016年超过33.3万公顷,2020年超过46.7万公顷,成为我省单一品种年种植面积最大的水稻品种,累计种植面积超过266.7万公顷。因其优异的食味品质和丰产性,南粳9108深受广大种植户和米业企业的喜爱,兴化大米、射阳大米、淮安大米、建湖大米、海安大米、姜堰大米、阜宁大米等地产大米品牌均以南粳9108作为原粮品种,为助推乡村振兴,带动农业增效、农民增收,提升“苏米”产业升级奠定了良好的基础。
3.4 南粳2728
南粳2728是以武粳15为母本,南粳5055为父本杂交,经稻瘟病和条纹叶枯病抗性基因及半糯基因分子标记辅助选择,结合田间多代选育、抗性鉴定和食味品质鉴定,于2013年育成的食味品质优、稻瘟病抗性好、丰产性好的中熟中粳稻新品种,2018年通过江苏省审定(审定编号:苏审稻20180005),2019年通过安徽省引种备案(备案号:皖引稻2019066),适宜江苏省和安徽省淮北地区种植[52]。该品种株型紧凑,长势较旺,分蘖力强,成穗率高,群体整齐度好,抗倒性较强,穗型中等,半直立穗,叶色绿,叶姿较挺,后期熟相好。每亩有效穗23.8万,每穗实粒数111.8粒,结实率93.0%,千粒重27.2 g,株高101.4 cm,全生育期150 d,与对照徐稻3号相当。该品种对白叶枯病、稻瘟病、条纹叶枯病和纹枯病均具有较好的抗性。根据农业农村部食品质量监督检测测试中心2016年检测:糙米率84.2%,精米率74.2%,整精米率69.7%,垩白率20.0%,垩白度4.9%。2017年在“连天下”第二届连云港优质稻米品鉴会中荣获金奖;2018年在第二届江苏优质稻米博览会“江苏好大米”评选中荣获金奖。2019年在寻找江苏最好吃的大米暨第五届江苏百姓品米节优质大米评比中,南粳2728获得特等奖。2020年1月,在江苏省优质稻品种食味品质评鉴会上荣获金奖。南粳2728丰产稳产性好,适应性强,适宜多种轻简栽培方式。一般亩产650 kg左右,高产田块可达700 kg以上。2016-2017年在泗洪、泗阳、宿迁、睢宁、丰县、邳州、铜山、东海、连云港、灌南、大丰、建湖等地示范种植,表现出食味品质优良,丰产稳产性好,熟期早,广适性好等优点,平均亩产达681.2 kg,其中2017年在东海县平明镇的百亩示范方经江苏省农委组织的专家组验收,平均亩产达到709.7 kg,表现出较高的产量潜力。南粳2728的成功选育填补了江苏淮北地区缺乏优良食味粳稻品种的空白,具有广阔的推广应用前景。
3.5 南粳5718
南粳5718是以自主选育的优良食味抗病粳稻中间品系宁7022为母本,与盐都地区农科所选育的优质高产粳稻中间品系盐粳608杂交,经系谱法和分子标记辅助选择,于2015年育成的食味品质优、丰产性好的中熟中粳稻新品种。2019年通过江苏省审定(审定编号:苏审稻20190004),2021年通过安徽省引种备案(备案号:皖引稻2021044),2020年已完成江苏省迟熟中粳组生产试验,适合江苏省和安徽省淮北地区、江苏省苏中地区及宁镇扬丘陵地区种植。该品种苗期叶色深绿,叶姿略披,苗体矮壮,抽穗后叶片挺立、株型紧凑,植株生长清秀,后期转色快,熟相好。全生育期147.9 d,与对照徐稻3号相当。株高102.6 cm,分蘖力中等,成穗率高。每亩有效穗21.2万,穗型较大,每穗总粒数131.5,结实率88.9%,千粒重28.8 g。2017-2018年江苏省农科院植保所接种鉴定,中抗白叶枯病、中感穗颈瘟、中抗条纹叶枯病、抗纹枯病。根据农业农村部食品质量监督检测测试中心2018年检测:糙米率84%,精米率72.2%,整精米率68.8%,垩白率31.0%,垩白度6.8%[53]。2018年在宿迁市和阜宁县优质米评鉴中,南粳5718均获得第一名;2019年和2020年,在寻找江苏最好吃的大米暨第五届和第六届江苏百姓品米节优质大米评比中,南粳5718均获得特等奖;2020年1月,在第四届“华西村杯”江苏好大米评鉴推介会上,南粳5718获得金奖;在江苏省优质稻品种食味品质评鉴会上,南粳5718获得特等奖;在首届和第二届中国好米榜评选活动中,南粳5718均获得“超级好吃”五星大奖。南粳5718除了食味品质好以外,最突出的优点是其茎秆粗壮、抗倒性极强、特别适宜稻虾共作等综合种养模式,而且穗型大、千粒重高、产量潜力大。一般亩产650 kg左右,高产田块可达700 kg以上。2017年和2018年两年平均亩产672.1 kg,比对照增产5.6%;2018年生产试验平均亩产689.1 kg,比对照徐稻3号增产5.1%。2017-2020年在江苏淮北和苏中地区示范种植,品质、抗性和丰产性表现都十分突出。2019年和2020年,农业农村部超级稻验收专家组对南粳5718百亩示范方进行现场测产验收,平均亩产分别达到887.7 kg和792.5 kg,达到了长江中下游地区一季粳稻的“超级稻”产量指标。南粳5718的成功选育丰富了江苏省淮北地区优良食味粳稻种植的品种需求,尤其是为我省稻虾共作等综合种养提供了食味品质优、抗倒性强、产量高的优良食味粳稻品种,具有广阔的应用前景。
3.6 南粳58
南粳58是以自主选育的优良食味抗病粳稻中间品系宁7020为母本,与江苏高产粳稻扬粳9538杂交,经稻瘟病抗性和半糯基因分子标记辅助选择,结合田间多代选育、抗性鉴定和食味品质筛选,于2016年育成的食味品质优、丰产性好的中熟中粳稻新品种。2019年通过江苏省审定(审定编号:苏审稻20190007),适合江苏省淮北地区种植。该品种综合性状好。据2017-2018年区试平均结果,全生育期142.2 d,比对照苏秀867早0.7 d,属于中熟中粳早熟类型。株高100.6 cm,株型较紧凑,分蘖力中等偏上,每穗实粒数115.1粒,分蘖力较强,生长势较旺,亩有效穗22.6万;叶色中绿,叶姿较挺,成熟期转色好,灌浆速度快。结实率89.5%,千粒重26.2 g。据江苏省农科院植物保护研究所抗性鉴定,南粳58中感穗颈瘟、白叶枯病和条纹叶枯病,抗纹枯病。据2018年江苏省区域试验统一取样送农业农村部食品质量检测中心检测,南粳58糙米率84.4%,精米率72.6%,整精米率68.6%。2019年获得连云港市赣榆区首届优质稻米品鉴“海州湾好大米”水稻品种称号,在宿迁市第二届“宿有千香”优质稻米品鉴中被评为“宿迁水稻好品种”,在“2019寻找江苏最好吃的大米”暨第五届江苏百姓品米节优质大米评比中获得“特等奖”,在2020年首届中国“好米榜”评比活动中获得“非常好吃”四星大奖。南粳58丰产性较好,2017年和2018年两年区试平均亩产647.3 kg,较对照苏秀867增产5.3%。2018年参加生产试验,平均亩产656.1 kg,比对照苏秀867增产3.6%。2018年在山东郯城、江苏阜宁、东海、丰县、灌云等10个适宜地区进行试验示范,南粳58表现丰产性稳产性好,平均亩产656.7 kg,其中郯城点产量最高,亩产达683.5 kg。南粳58不仅食味品质佳,有香味,而且具备早熟、丰产稳产性好的特性,由于同时含有稻瘟病抗性基因Pi-ta和Pi-b,稻瘟病抗性较好。
表3列出了南粳系列代表性品种及其亲本的食味品质、抗病、粒型相关性状的基因型,供育种者参考。
Table 3
表3
表3南粳系列代表性品种及其亲本的食味品质、抗病、粒型相关性状的基因型
Table 3
| 品种 | 基因型 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Wxmp | SSIIa | SSIIIa | PUL | Badh2 | Stv-bi | Pi-ta | Pi-b | Pi-km | GW5 | |
| 南粳46 | + | - | + | - | + | + | - | + | - | + |
| 南粳5055 | + | - | + | - | - | + | - | + | - | + |
| 南粳9108 | + | - | + | - | + | + | - | + | - | + |
| 南粳2728 | + | + | + | - | - | + | + | + | + | + |
| 南粳5718 | + | - | + | + | - | + | - | - | + | + |
| 南粳58 | + | - | + | + | + | + | + | + | + | + |
| 武粳13 (亲本) | - | + | + | - | - | - | - | + | - | + |
| 武粳15 (亲本) | - | + | + | + | - | - | + | + | + | + |
| 武香粳14 (亲本) | - | - | + | - | + | - | - | + | - | - |
| 关东194 (亲本) | + | - | + | - | - | + | - | - | + | + |
新窗口打开|下载CSV
4 江苏省优良食味粳稻的食味品质特性
4.1 稻米理化特性
为了明确江苏省优良食味粳稻的理化品质特性,分别以9个已审定的南粳系列半糯粳稻和10个食味品质较好的江苏和北方粳稻为材料进行比较,并以南粳系列品种的亲本关东194、武香粳14和武粳13及日本品种越光作对比。测定结果表明,南粳系列半糯粳稻的AC (10.7%)显著低于非半糯粳稻(18.6%),PC (7.1%)和GC (86.5 mm)则显著高于非半糯粳稻(6.8%和74.0 mm),GT半糯粳稻略高,但两者间差异不显著。在RVA值上,半糯粳稻的峰值黏度(2887.1 cP)和崩解值(1369.3 cP)显著和极显著高于非半糯粳稻的对应值(2660.3 cP和951.4 cP),而最终黏度(2080.4 cP)、消减值(-781.6 cP)和峰值时间(6.0 min)显著或极显著低于非半糯粳稻的对应值(2717.4 cP,57.0 cP和6.4 min),热浆黏度两者间差异不显著[56] (表4)。表明南粳系列半糯粳稻的AC低,GC高,崩解值大,消减值小,且均为负值,因而淀粉的黏性大,米饭柔软,口感较好。表4
Table 4
表4
表4江苏省半糯粳稻与非半糯粳稻的稻米理化特性比较
Table 4
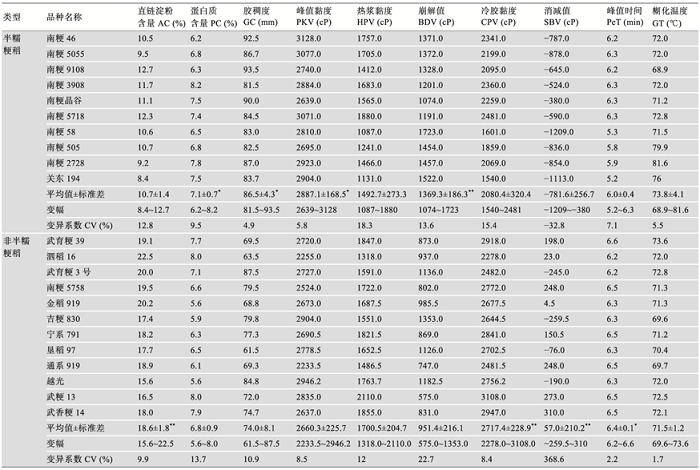 |
新窗口打开|下载CSV
4.2 食味品质特性
选用6个南粳系列半糯粳稻品种与5个北方优质粳稻进行米饭食味及质构特性(反映米饭硬度黏度的特性)分析,结果表明,所有品种的综合食味值均在80分以上,其中南粳46、南粳5718、南粳9108、南粳58、金稻919、吉粳830与日本越光的食味值相当,而南粳505、南粳2728、宁系791、垦稻97、通系919稍低,各品种在米饭外观、口感上与综合食味值表现一致。在米饭质构特性上,南粳系列品种的弹性整体上高于北方粳稻;硬度和黏度值因品种而异,南粳品种和北方品种间无显著差异;在平衡度上,食味值较高的品种往往具有更高的平衡度值。此外,6个南粳品种的食味特性和质构特性明显优于亲本武香粳14和武粳13;除了南粳2728以外,综合食味值也比关东194明显提高(表5)。Table 5
表5
表5南粳系列半糯粳稻品种与其他粳稻品种食味品质及米饭质构特性比较
Table 5
| 品种类型 | 品种 | 食味特性 | 质构特性 | |||||
|---|---|---|---|---|---|---|---|---|
| 外观 | 口感 | 综合食味值 | 硬度 | 黏性 | 弹性 | 平衡度 | ||
| 南粳粳稻 | 南粳46 | 8.40±0.14ab | 8.50±0.14a | 87.5±0.7a | 5.39±0.15e | 1.04±0.03ab | 0.78±0.02a | 0.19±0.01a |
| 南粳5718 | 8.50±0.00ab | 8.55±0.07a | 87.0±0.0a | 5.12±0.08f | 0.69±0.06e | 0.78±0.01a | 0.14±0.00d | |
| 南粳9108 | 7.65±0.07d | 7.95±0.07c | 84.5±0.7bc | 7.62±0.06ab | 1.01±0.04ab | 0.68±0.01b | 0.13±0.00de | |
| 南粳58 | 7.90±0.14c | 8.00±0.14c | 84.5±0.7bc | 4.60±0.09g | 0.69±0.05e | 0.78±0.01a | 0.15±0.01c | |
| 南粳505 | 7.50±0.14d | 7.75±0.07d | 83.5±0.7c | 5.14±0.11f | 0.68±0.08e | 0.79±0.01a | 0.13±0.02de | |
| 南粳2728 | 7.25±0.07e | 7.55±0.07e | 81.0±0.0d | 5.54±0.02e | 0.60±0.11f | 0.76±0.00ab | 0.11±0.02e | |
| 北方粳稻 | 金稻919 | 8.55±0.07a | 8.50±0.14a | 86.0±0.0ab | 5.16±0.04f | 0.77±0.04d | 0.65±0.04bc | 0.15±0.01c |
| 吉粳830 | 8.20±0.00b | 8.25±0.07c | 84.0±0.0bc | 5.49±0.06e | 1.00±0.03ab | 0.66±0.02b | 0.18±0.01ab | |
| 宁系791 | 8.25±0.07b | 8.15±0.07c | 83.5±0.7c | 7.03±0.12c | 0.94±0.01bc | 0.67±0.01b | 0.13±0.00de | |
| 垦稻97 | 8.40±0.00ab | 8.30±0.00bc | 83.0±1.4cd | 6.11±0.15d | 1.01±0.08ab | 0.66±0.01b | 0.16±0.01b | |
| 通系919 | 8.20±0.14b | 8.10±0.14c | 82.5±0.7cd | 7.77±0.03a | 1.12±0.11a | 0.65±0.08bc | 0.14±0.02d | |
| 日本粳稻 亲本 | 越光 | 8.35±0.07ab | 8.35±0.07ab | 84.5±0.7bc | 7.31±0.04bc | 0.89±0.12c | 0.60±0.04c | 0.12±0.02e |
| 武香粳14 | 7.80±.06c | 7.90±0.04c | 78.0±0.5e | 7.60±0.11ab | 0.85±0.04c | 0.62±0.01c | 0.11±0.01e | |
| 武粳13 | 7.81±0.05c | 7.72±0.05d | 75.1±0.3f | 7.32±0.07bc | 0.68±0.03e | 0.61±0.00c | 0.09±0.00f | |
| 关东194 | 8.00±0.02c | 8.40±0.07ab | 81.4±0.0d | 7.32±0.02bc | 1.10±0.05a | 0.75±0.02ab | 0.15±0.03 | |
新窗口打开|下载CSV
4.3 支链淀粉链长分布
近年来的研究表明,支链淀粉的含量及其链长分布也是影响稻米食味品质的重要因素。支链淀粉的短分支链比例较高,而长分支链比例较低,食味品质较好[57,58]。用荧光糖电泳法对支链淀粉DP6~50链长分布检测结果表明,DP6~12的超短链(A链)比例最高的是南粳3908和南粳晶谷,其次为稻花香、南粳46、越光、南粳9108、南粳58和南粳5055,A链比例均在50%以上,南粳2728、武粳13、武香粳14和关东194的A链比例较低,南粳2728仅为44.38%。而B1 (DP13~24)、B2 (DP25~36)、B3 (DP≥37)链的比例与A链呈相反趋势(表6)。表明食味品质优异的品种,往往具有较高的A链比例和较低的B链比例。A链比例高、B链比例低,说明淀粉的短链多,中长链少,淀粉的糊化特性好,米饭比较柔软,口感较好。Table 6
表6
表6南粳系列品种与其他粳稻品种的支链淀粉分支链比例的比较
Table 6
| 品种 | A链(DP6~12) | B1链(DP13~24) | B2链(DP25~36) | B3链(DP≥37) |
|---|---|---|---|---|
| 南粳46 | 51.50±0.06b | 45.07±0.07f | 2.89±0.02ef | 0.54±0.02c |
| 南粳5055 | 50.08±0.01d | 46.45±0.01d | 2.87±0.02ef | 0.60±0.01bc |
| 南粳9108 | 50.45±0.08cd | 45.83±0.07e | 3.23±0.03c | 0.53±0.00c |
| 南粳2728 | 44.38±0.13f | 51.75±0.14a | 3.06±0.01d | 0.77±0.00a |
| 南粳58 | 50.17±0.80d | 45.65±0.80e | 3.30±0.01bc | 0.82±0.01a |
| 南粳3908 | 52.39±0.29a | 44.29±0.12g | 2.79±0.16f | 0.62±0.13b |
| 南粳晶谷 | 52.27±0.24a | 44.13±0.30g | 2.95±0.02de | 0.62±0.04b |
| 越光 | 50.94±0.06bc | 44.78±0.16f | 3.35±0.02b | 0.80±0.02a |
| 关东194 | 49.36±0.07e | 46.61±0.06d | 3.37±0.02b | 0.64±0.01b |
| 武粳13 | 45.77±0.06f | 49.83±0.07b | 3.64±0.02a | 0.76±0.02a |
| 武香粳14 | 48.81±0.09e | 47.29±0.04c | 3.05±0.01d | 0.63±0.02b |
| 稻花香 | 51.74±0.03b | 44.48±0.10f | 3.02±0.06d | 0.74±0.05a |
新窗口打开|下载CSV
5 结语与展望
江苏省位于北亚热带湿润季风气候区,水稻抽穗以后气温较高,日平均气温一般都在25℃以上,昼夜温差小;江苏水稻土有机质含量低;江苏省又是水稻高产省份,氮肥施用量大,水稻每亩施纯氮一般在20 kg以上,多的甚至高达30 kg。因此,江苏生产的大米往往外观品质较差、蛋白质含量较高。而我国东北地区则相反,水稻抽穗以后气温较低,日平均气温一般都在22℃左右,昼夜温差大;东北黑土地有机质含量高;氮肥施用量至少比江苏少1/3。因此,东北生产的大米外观品质好,蛋白质含量低。所以,江苏要选育象东北那样外观品质好、直链淀粉含量较高的优质米品种难度较大。为了培育适合江苏居民口感的优质米品种,本研究团队通过对稻米食味品质性状的遗传研究,采取降低直链淀粉含量的策略,筛选出直链淀粉含量在10%左右的日本半糯粳稻品种“关东194”作为食味品质改良的核心种质,与江苏高产粳稻品种杂交,通过分子标记辅助选择,结合产量、抗性、食味等性状的表型鉴定,育成南粳46、南粳5055、南粳9108、南粳5718等系列品种,不仅食味品质优,而且产量高、抗性好,种植面积不断扩大。用南粳系列品种开发的优质大米深受长江中下游地区消费市场和广大居民的喜爱,为抽穗灌浆期间气温较高的长江中下游粳稻区培育食味品质较好的粳稻品种闯出了一条新路。近年来,上海、浙江、安徽等周边省、市利用南粳46等的低直链淀粉含量基因Wxmp育成了一批新的半糯粳稻品种在生产上推广应用。今后几年,半糯粳稻品种在长江中下游粳稻区的种植面积将进一步扩大。不仅如此,南方籼稻区同样是采用了降低直链淀粉含量的技术途径,将籼稻的Wxa基因替换成Wxb基因,使籼稻的食味品质得到了大幅度提升。由此可见,通过降低直链淀粉含量来提高稻米的食味品质是籼粳稻普遍适用的技术途径,也证明直链淀粉含量是影响稻米食味品质最主要的因素。利用低直链淀粉含量基因Wxmp培育的半糯粳稻品种虽然食味品质较好,但也带来外观品质较差的弊端。此外,目前在生产上大面积推广种植的南粳46、南粳9108等品种的稻瘟病抗性较差。进一步提高江苏粳稻的稻米品质,培育既好看又好吃的粳稻品种,需要合理利用Wxmp基因以外的淀粉合成相关基因(如SSIIa、SSIIIa、PUL等[24,59])和Wx位点的其他突变基因(如Wxhp[60]、Wxmw [61]、Wxlv[62]、Wxla [63]等);随着生物技术的发展,通过基因编辑创造新的等位变异[64,65,66]也为未来的研究提供了更加广阔的途径。同时,目前江苏粳稻育种利用的稻瘟病抗性基因主要是Pi-b和Pi-ta,稻瘟病抗性的进一步改良,需要拓宽抗性基因的利用。据报道,Pi-gm[67]和Pi-km[68]对江苏稻瘟病生理小种的抗性较好,Pi-gm已经成功应用于江苏粳稻稻瘟病抗性的改良[69],可以扩大利用。此外,理想株型基因[70]、绿色革命伴侣基因[71]、直立穗基因[72]、粒型基因[73]等的利用也为江苏粳稻产量潜力的提高提供了广阔的前景。值得指出的是,蛋白质含量也是影响食味品质的主要因素。虽然降低直链淀粉含量是气温较高的南方稻区提升稻米食味品质的有效技术途径,但随着稻米食味品质遗传调控机制的进一步明晰,稻米食味品质遗传改良的路径必将进一步完善。
(责任编委: 王秀娥)
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}