孙松,
孙天昊,
林文雄,
福建农林大学作物科学学院/作物遗传育种与综合利用教育部重点实验室/作物生态与分子生理学福建省高校重点实验室 福州 350002
基金项目: 中国博士后科学基金2014T70603
教育部留学归国人员科研启动基金Education Department Liu[2013]1792
国家自然科学基金青年科学基金项目31501232
详细信息
作者简介:贾琪, 研究方向为植物抗逆性与分子遗传学。E-mail:jiaqi@fafu.edu.cn
通讯作者:林文雄, 研究方向为植物生理与分子生态学。E-mail:wenxiong181@163.com
中图分类号:Q945;Q948.1;Q37计量
文章访问数:893
HTML全文浏览量:2
PDF下载量:1634
被引次数:0
出版历程
收稿日期:2017-12-15
录用日期:2018-04-20
刊出日期:2018-08-01
Mechanism of F-box protein family in plant resistance response to environmental stress
JIA Qi,SUN Song,
SUN Tianhao,
LIN Wenxiong,
College of Crop Sciences, Fujian Agriculture and Forestry University/Key Laboratory of Ministry of Education for Genetics, Breeding and Multiple Crop Utilization/Key Laboratory of Crop Ecology and Molecular Physiology(Fujian Province University), Fuzhou 350002, China
Funds: the China Postdoctoral Science Foundation2014T70603
the Scientific Research Starting Foundation for the Returned Overseas Chinese Scholars, Ministry of Education of ChinaEducation Department Liu[2013]1792
the National Natural Sciences Foundation of China31501232
More Information
Corresponding author:LIN Wenxiong, E-mail:wenxiong181@163.com
摘要
HTML全文
图
参考文献
相关文章
施引文献
资源附件
访问统计
摘要
摘要:SCF复合体泛素连接酶E3介导的泛素化蛋白降解是翻译后水平上对生命进程进行调控的一个重要方式。它的关键组分F-box蛋白负责识别被降解的靶底物蛋白。植物F-box基因家族成员众多,极具多样性。F-box蛋白N端常含F-box基序,C端常为蛋白互作保守结构域,该结构具多样性,可识别不同底物,是F-box蛋白分类的依据。研究表明,F-box蛋白参与调控植物的许多生命进程,包括抗逆反应。本文就近年来F-box蛋白在植物抗逆反应中的作用机制进行总结。F-box蛋白大多以SCF复合体泛素连接酶E3介导的泛素化蛋白降解目标蛋白的方式调控抗逆反应,也有不依赖形成SCF复合体的方式行使功能,不少F-box蛋白参与了植物激素信号传导,通过调控转录因子活性而改变下游基因的表达,由此影响了植物的抗逆反应。基因表达谱的生物信息学预测表明,大多数F-box基因参与了植物抗逆反应,目前只有其中一小部分已报道了其抗逆调节功能。在此综述了这些F-box蛋白在植物抗逆胁迫中的研究进展。在干旱和盐碱胁迫反应中,F-box基因常通过影响植物激素脱落酸、乙烯等植物激素信号传导而调控抗逆。由于干旱和盐碱胁迫具协同性,不少F-box基因同时参与抗旱和抗盐碱胁迫,但调节方式有所不同,一些F-box基因对抗干旱和盐碱的反应具协同性,从总体上调控植物的渗透胁迫和离子毒害反应;而另一些F-box基因对干旱和盐胁迫反应的调节作用相反,它们可能在植物抗逆的精细调节中起作用。在低温胁迫反应中,F-box蛋白可调节植物抗低温的CBF信号途径。在生物胁迫反应中,F-box基因常通过影响植物激素茉莉酸和水杨酸途径来调控抗病,病原菌也以攻击植物SCF复合体使植物致病。此外,植物激素信号途径之间相互作用,共同影响抗逆反应。
关键词:F-box蛋白家族/
SCF复合体/
植物/
环境胁迫/
抗逆反应
Abstract:The UPS (ubiquitin proteasome system) mediated by SCF type E3 ubiquitin-ligase is an important mechanism to regulate biological progress at post-translation level. F-box protein, as a key component in SCF complex, could recognize its target protein for degradation. F-box gene family contains numerous members with vast diversity. In general, F-box protein contains F-box motif at N terminus and conserved domain of protein-protein interaction for recognizing target at C terminus. Due to vast diversity of conserved C terminus domains, F-box proteins could recognize wide varieties of targets. Also based on C terminal domains, F-box proteins could be divided into several subfamilies. It showed that plant F-box proteins were involved in many life processes, including response to environmental stress. Here, we reviewed current knowledge of plant F-box proteins in responding to stress. Most of the reported F-box proteins had been shown to function via SCF-dependent protein degradation, with few using SCF-independent mechanisms. Some well-understood F-box proteins were involved in phytohormone signaling pathways. Some reacted to stress through regulating the activity of transcription factors, which influenced expression of downstream genes responding to stress. Bioinformatics analyses of transcriptome showed that many predicted F-box genes were involved in stress-response reactions. Among these, only a few studies had dealt with the functions. The knowledge on the functions under environmental stress was summarized in this study. For drought, salinity and alkality stresses, F-box genes often regulated abscisic acid or ethylene signal pathways. Since drought and salt-alkaline stresses often occurred concomitantly, quite a few F-box genes had been identified to be involved in the response to both stresses in different ways. Some regulated the response to osmotic stress and ionic stress synergistically. However, some functioned inversely, suggesting that they played a role in fine regulations. For cold stress, F-box genes regulated CBF signal pathways. For biotic stress, F-box genes always regulated jasmonate and salicylic acid pathways. Meanwhile, pathogens attacked plant SCF complex for infection. Moreover, phytohormones had crosstalk to coordinate resistance in plants.
Key words:F-box protein/
SCF complex/
Plant/
Environment stress/
Response to stress
HTML全文
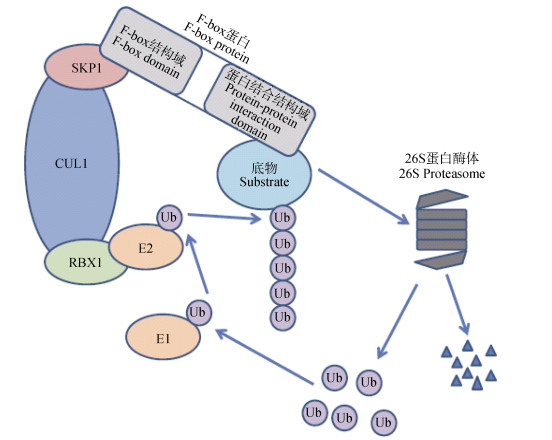
图1复合体介导的蛋白泛素化降解
Figure1.SCF complex mediated protein ubiquitylation and protein degradation
 下载: 全尺寸图片幻灯片
下载: 全尺寸图片幻灯片表1参与植物激素信号传导的F-box蛋白
Table1.F-box proteins involved in phytohormone signaling
| F-box蛋白 F-box protein | 靶蛋白结合序列 Conserved domains for interaction with targets | 物种 Species | 激素途径 Phytohormone pathways | 靶蛋白 Targets | 参考文献 References |
| TIR1, AFB1-5 | LRR | 拟南芥? Arabidopsis thaliana | 生长素受体?Auxin receptor | AUX/IAA(负调控蛋白) (Negative regulator) | [39-40, 42-43] |
| COI1 | LRR | 拟南芥? Arabidopsis thaliana | 茉莉酸受体?JA receptor | JAZ(负调控蛋白) (Negative regulator) | [44-46] |
| SLY1, SNE | GGF, LSL | 拟南芥? Arabidopsis thaliana | 与赤霉素受体GID1互作?Interact with GA receptor GID1 | DELLAs | [47-49] |
| GID2 | GGF, LSL | 水稻?Oryza sativa | DELLAs | [50-52] | |
| EBF1-2 | LRR | 拟南芥? Arabidopsis thaliana | 乙烯?Ethylene | EIN3, EIL1(正调控蛋白) ? (Positive regulator) | [53-55] |
| ETP1-2 | FBA | EIN2 (稳定EIN3, EIL1) ? (Stabilize EIN3, EIL1) | [56-57] | ||
| MAX2 | LRR | 拟南芥? Arabidopsis thaliana | 与独角金内酯受体D14互作?Interact with SL receptor D14 | BES1 | [58-60] |
| DWARF3 (D3) | LRR | 水稻?Oryza sativa | D53 | [61-62] | |
| RMS4 | LRR | 豌豆? Pisum sativum | 独角金内酯途径, 与MAX2同源?SL pathway, homolog of MAX2 | — | [63] |
| MAX2a | LRR | 矮牵牛花? Petunia hybrida | 与拟南芥、水稻独角金内酯受体D14同源的DAD2互作?Interact with DAD2, a homolog of D14 | — | [64-65] |
| RIFP1 | LRR | 拟南芥? Arabidopsis thaliana | ABA途径, 负调控蛋白?ABA pathway, negative regulator | RCAR3(ABA受体) (ABA receptor) | [66] |
| PP2-B11 | PP2 | ABA途径, 负调控蛋白?ABA pathway, negative regulator | SnRK2.3/AtLEA14 | [67-69] | |
| DOR | FBA | ABA合成, 负调控蛋白?ABA synthesis, negative regulator | [70] | ||
| EDL3 | LRR | ABA途径, 正调控蛋白?ABA pathway, positive regulator | — | [71] | |
| TLP9 | Tubby | ABA途径, 正调控蛋白?ABA pathway, positive regulator | — | [72] | |
| FOA1 | FBA | ABA途径, 负调控蛋白?ABA pathway, negative regulator | — | [73] | |
| AFBA1 | FBA | 拟南芥? Arabidopsis thaliana | ABA途径, 正调控蛋白, 与MYB44结合, 保持其稳定性?ABA pathway, positive regulator, interact with MYB44 to stabilize it | ?转录因子TCP4, TCP4, MYB44 Transcriptor factors | [74] |
| CPR1/CPR30 | FBA | 拟南芥? Arabidopsis thaliana | 水杨酸途径, 负调控蛋白?SA pathway, negative regulator | SNC1 RPS2 | [75-78] |
| KIB1 | Kelch | 拟南芥? Arabidopsis thaliana | 芸苔素途径, 正调控蛋白?BR pathway, positive regulator | BIN2 | [79] |
| “?”表示酵母双杂交找到的互作底物蛋白, 未有介导26S蛋白水解酶降解的证据。“?”: the substrates had been identified by yeast two hybrid, but no evidence supported that they would be degraded by 26S proteasome. | |||||
下载: 导出CSV参考文献
| [1] | SMALLE J, VIERSTRA R D. The ubiquitin 26S proteasome proteolytic pathway[J]. Annual Review Plant Biology, 2004, 55:555-590 doi: 10.1146/annurev.arplant.55.031903.141801 |
| [2] | VIERSTRA R D. The ubiquitin-26S proteasome system at the nexus of plant biology[J]. Nature Reviews Molecular Cell Biology, 2009, 10(6):385-397 doi: 10.1038/nrm2688 |
| [3] | TONE S L. The role of ubiquitin and the 26S proteasome in plant abiotic stress signaling[J]. Frontiers in Plant Science, 2014, 5:135 |
| [4] | SADANANDOM A, BAILEY M, EWAN R, et al. The ubiquitin-proteasome system:Central modifier of plant signalling[J]. The New Phytologist, 2012, 196(1):13-28 doi: 10.1111/j.1469-8137.2012.04266.x |
| [5] | LECHNER E, ACHARD P, VANSIRI A, et al. F-box proteins everywhere[J]. Current Opinion in Plant Biology, 2006, 9(6):631-638 doi: 10.1016/j.pbi.2006.09.003 |
| [6] | SOMERS D E, FUJIWARA S. Thinking outside the F-box:Novel ligands for novel receptors[J]. Trends in Plant Science, 2009, 14(4):206-213 doi: 10.1016/j.tplants.2009.01.003 |
| [7] | 秘彩莉, 刘旭, 张学勇. F-box蛋白质在植物生长发育中的功能[J].遗传, 2006, 28(10):1337-1342 doi: 10.3321/j.issn:0253-9772.2006.10.026 BI C L, LIU X, ZHANG X Y. The function of F-box protein in plant growth and development[J]. Hereditas, 2006, 28(10):1337-1342 doi: 10.3321/j.issn:0253-9772.2006.10.026 |
| [8] | 许媛, 李铃仙, 于秀梅, 等. F-box蛋白在植物抗逆境胁迫中的功能[J].植物生理学报, 2015, 51(7):1003-1008 http://www.cqvip.com/QK/90791X/201507/665670034.html XU Y, LI L X, YU X M, et al. The functions of F-box protein in plant resistance to stress[J]. Plant Physiology Journal, 2015, 51(7):1003-1008 http://www.cqvip.com/QK/90791X/201507/665670034.html |
| [9] | 李莉, 李懿星, 夏凯, 等.植物F-box蛋白及其生物学功能研究[J].安徽农业科学, 2010, 38(35):19879-19881 doi: 10.3969/j.issn.0517-6611.2010.35.001 LI L, LI Y X, XIA K, et al. Plant F-box proteins and their biological functions[J]. Journal of Anhui Agricultural Sciences, 2010, 38(35):19879-19881 doi: 10.3969/j.issn.0517-6611.2010.35.001 |
| [10] | 吴丹, 唐冬英, 李新梅, 等. F-box蛋白在植物生长发育中的功能研究进展[J].生命科学研究, 2015, 19(4):362-367 http://www.cqvip.com/QK/90895X/201504/665927296.html WU D, TANG D Y, LI X M, et al. Progresses on F-box protein function in plant growth and development[J]. Life Science Research, 2015, 19(4):362-367 http://www.cqvip.com/QK/90895X/201504/665927296.html |
| [11] | BAI C, SEN P, HOFMANN K, et al. SKP1 connects cell cycle regulators to the ubiquitin proteolysis machinery through a novel motif, the F-box[J]. Cell, 1996, 86(2):263-274 doi: 10.1016/S0092-8674(00)80098-7 |
| [12] | XU G X, MA H, NEI M, et al. Evolution of F-box genes in plants:Different modes of sequence divergence and their relationships with functional diversification[J]. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(3):835-840 doi: 10.1073/pnas.0812043106 |
| [13] | HUA Z H, ZOU C, SHIU S H, et al. Phylogenetic comparison of F-Box (FBX) gene superfamily within the plant kingdom reveals divergent evolutionary histories indicative of genomic drift[J]. PLoS One, 2011, 6(1):e16219 doi: 10.1371/journal.pone.0016219 |
| [14] | JIA Q, XIAO Z X, WONG F L, et al. Genome-wide analyses of the soybean F-box gene family in response to salt stress[J]. International Journal of Molecular Sciences, 2017, 18(4):818 doi: 10.3390/ijms18040818 |
| [15] | 许克恒, 张云彤, 张莹, 等.植物F-box基因家族的研究进展[J].生物技术通报, 2018, 34(1):26-32 http://www.cqvip.com/QK/90799X/201609/670170137.html XU K H, ZHANG Y T, ZHANG Y, et al. Research advances on the F-box gene family in plants[J]. Biotechnology Bulletin, 2018, 34(1):26-32 http://www.cqvip.com/QK/90799X/201609/670170137.html |
| [16] | JIA F J, WU B J, LI H, et al. Genome-wide identification and characterisation of F-box family in maize[J]. Molecular Genetics and Genomics, 2013, 288(11):559-577 doi: 10.1007/s00438-013-0769-1 |
| [17] | GUPTA S, GARG V, KANT C, et al. Genome-wide survey and expression analysis of F-box genes in chickpea[J]. BMC Genomics, 2015, 16:67 doi: 10.1186/s12864-015-1293-y |
| [18] | CUI H R, ZHANG Z R, LYU W, et al. Genome-wide characterization and analysis of F-box protein-encoding genes in the Malus domestica genome[J]. Molecular Genetics and Genomics, 2015, 290(4):1435-1446 doi: 10.1007/s00438-015-1004-z |
| [19] | JAIN M, NIJHAWAN A, ARORA R, et al. F-box proteins in rice. Genome-wide analysis, classification, temporal and spatial gene expression during panicle and seed development, and regulation by light and abiotic stress[J]. Plant Physiology, 2007, 143(4): 1467-1683 |
| [20] | WANG G M, YIN H, QIAO X, et al. F-box genes:Genome-wide expansion, evolution and their contribution to pollen growth in pear (Pyrus bretschneideri)[J]. Plant Science, 2016, 253:164-175 doi: 10.1016/j.plantsci.2016.09.009 |
| [21] | SONG J B, WANG Y X, LI H B, et al. The F-box family genes as key elements in response to salt, heavy mental, and drought stresses in Medicago truncatula[J]. Functional & Integrative Genomics, 2015, 15(4):495-507 https://www.ncbi.nlm.nih.gov/pubmed/25877816 |
| [22] | YANG X H, KALLURI U C, JAWDY S, et al. The F-box gene family is expanded in herbaceous annual plants relative to woody perennial plants[J]. Plant Physiology, 2008, 148(3):1189-1200 doi: 10.1104/pp.108.121921 |
| [23] | BELLIENY-RABELO D, OLIVEIRA A E A, VENANCIO T M. Impact of whole-genome and tandem duplications in the expansion and functional diversification of the F-box family in legumes (Fabaceae)[J]. PLoS One, 2013, 8(2):e55127 doi: 10.1371/journal.pone.0055127 |
| [24] | GAGNE J M, DOWNES B P, SHIU S H, et al. The F-box subunit of the SCF E3 complex is encoded by a diverse superfamily of genes in Arabidopsis[J]. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99(17):11519-11524 doi: 10.1073/pnas.162339999 |
| [25] | KURODA H, TAKAHASHI N, SHIMADA H, et al. Classification and expression analysis of Arabidopsis F-boxcontaining protein genes[J]. Plant and Cell Physiology, 2002, 43(10):1073-1085 doi: 10.1093/pcp/pcf151 |
| [26] | SCHUMANN N, NAVARRO-QUEZADA A, ULLRICH K, et al. Molecular evolution and selection patterns of plant F-box proteins with C-terminal kelch repeats[J]. Plant Physiology, 2011, 155(2):835-850 doi: 10.1104/pp.110.166579 |
| [27] | NAVARRO-QUEZADA A, SCHUMANN N, QUINT M. Plant F-box protein evolution is determined by lineage-specific timing of major gene family expansion waves[J]. PLoS One, 2013, 8(7):e68672 doi: 10.1371/journal.pone.0068672 |
| [28] | HERMAND D. F-box proteins:More than baits for the SCF?[J]. Cell Division, 2006, 1:30 doi: 10.1186/1747-1028-1-30 |
| [29] | HO M S, OU C, CHAN Y R, et al. The utility F-box for protein destruction[J]. Cellular and molecular life sciences:CMLS, 2008, 65(13):1977-2000 doi: 10.1007/s00018-008-7592-6 |
| [30] | NELSON D E, RANDLE S J, LAMAN H. Beyond ubiquitination:The atypical functions of Fbxo7 and other F-box proteins[J]. Open Biology, 2013, 3(10):130131 doi: 10.1098/rsob.130131 |
| [31] | SONG J B, GAO S, SUN D, et al. miR394 and LCR are involved in Arabidopsis salt and drought stress responses in an abscisic acid-dependent manner[J]. BMC Plant Biology, 2013, 13:210 doi: 10.1186/1471-2229-13-210 |
| [32] | CHEN H, LI Z F, XIONG L M. A plant microRNA regulates the adaptation of roots to drought stress[J]. FEBS Letters, 2012, 586(12):1742-1747 doi: 10.1016/j.febslet.2012.05.013 |
| [33] | CHEN Z H, HU L Z, HAN N, et al. Overexpression of a miR393-resistant form of transport inhibitor response protein 1(mTIR1) enhances salt tolerance by increased osmoregulation and Na+ exclusion in Arabidopsis thaliana[J]. Plant and Cell Physiology, 2015, 56(1):73-83 doi: 10.1093/pcp/pcu149 |
| [34] | EGAMBERDIEVA D, WIRTH S J, ALQARAWI A A, et al. Phytohormones and beneficial microbes:Essential components for plants to balance stress and fitness[J]. Frontiers in Microbiology, 2017, 8:2104 doi: 10.3389/fmicb.2017.02104 |
| [35] | VERMA V, RAVINDRAN P, KUMAR P P. Plant hormonemediated regulation of stress responses[J]. BMC Plant Biology, 2016, 16:86 doi: 10.1186/s12870-016-0771-y |
| [36] | DERKSEN H, RAMPITSCH C, DAAYF F. Signaling crosstalk in plant disease resistance[J]. Plant Science, 2013, 207:79-87 doi: 10.1016/j.plantsci.2013.03.004 |
| [37] | BIELACH A, HRTYAN M, TOGNETTI V B. Plants under Stress:Involvement of auxin and cytokinin[J]. International Journal of Molecular Sciences, 2017, 18(7):1427 doi: 10.3390/ijms18071427 |
| [38] | STEFANOWICZ K, LANNOO N, VAN DAMME E J M. Plant F-box proteins-Judges between life and death[J]. Critical Reviews in Plant Sciences, 2015, 34(6):523-552 doi: 10.1080/07352689.2015.1024566 |
| [39] | DHARMASIRI N, DHARMASIRI S, ESTELLE M. The F-box protein TIR1 is an auxin receptor[J]. Nature, 2005, 435(7041):441-445 doi: 10.1038/nature03543 |
| [40] | TAN X, CALDERON-VILLALOBOS L I A, SHARON M, et al. Mechanism of auxin perception by the TIR1 ubiquitin ligase[J]. Nature, 2007, 446(7136):640-645 doi: 10.1038/nature05731 |
| [41] | KEPINSKI S, LEYSER O. The Arabidopsis F-box protein TIR1 is an auxin receptor[J]. Nature, 2005, 435(7041):446-451 doi: 10.1038/nature03542 |
| [42] | NAVARRO L, DUNOYER P, JAY F, et al. A plant miRNA contributes to antibacterial resistance by repressing auxin signaling[J]. Science, 2006, 312(5772):436-439 doi: 10.1126/science.1126088 |
| [43] | DHARMASIRI N, DHARMASIRI S, WEIJERS D, et al. Plant development is regulated by a family of auxin receptor F box proteins[J]. Developmental Cell, 2005, 9(1):109-119 doi: 10.1016/j.devcel.2005.05.014 |
| [44] | THINES B, KATSIR L, MELOTTO M, et al. JAZ repressor proteins are targets of the SCFCOI1 complex during jasmonate signalling[J]. Nature, 2007, 448(7154):661-665 doi: 10.1038/nature05960 |
| [45] | CHINI A, FONSECA S, FERNáNDEZ G, et al. The JAZ family of repressors is the missing link in jasmonate signalling[J]. Nature, 2007, 448(7154):666-671 doi: 10.1038/nature06006 |
| [46] | SHEARD L B, TAN X, MAO H B, et al. Jasmonate perception by inositol-phosphate-potentiated COI1-JAZ co-receptor[J]. Nature, 2010, 468(7322):400-405 doi: 10.1038/nature09430 |
| [47] | MCGINNIS K M, THOMAS S G, SOULE J D, et al. The Arabidopsis SLEEPY1 gene encodes a putative F-box subunit of an SCF E3 ubiquitin ligase[J]. The Plant Cell, 2003, 15(5):1120-1130 doi: 10.1105/tpc.010827 |
| [48] | STRADER L C, RITCHIE S, SOULE J D, et al. Recessiveinterfering mutations in the gibberellin signaling gene SLEEPY1 are rescued by overexpression of its homologue, SNEEZY[J]. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(34):12771-12776 doi: 10.1073/pnas.0404287101 |
| [49] | ARⅡZUMI T, LAWRENCE P K, STEBER C M. The role of two F-box proteins, SLEEPY1 and SNEEZY, in Arabidopsis gibberellin signaling[J]. Plant Physiology, 2011, 155(2):765-775 doi: 10.1104/pp.110.166272 |
| [50] | GOMI K, SASAKI A, ITOH H, et al. GID2, an F-box subunit of the SCF E3 complex, specifically interacts with phosphorylated SLR1 protein and regulates the gibberellindependent degradation of SLR1 in rice[J]. The Plant Journal:For Cell And Molecular Biology, 2004, 37(4):626-634 doi: 10.1111/j.1365-313X.2003.01990.x |
| [51] | UEGUCHI-TANAKA M, HIRANO K, HASEGAWA Y, et al. Release of the repressive activity of rice DELLA protein SLR1 by gibberellin does not require SLR1 degradation in the gid2 mutant[J]. The Plant Cell, 2008, 20(9):2437-2446 doi: 10.1105/tpc.108.061648 |
| [52] | SASAKI A, ITOH H, GOMI K, et al. Accumulation of phosphorylated repressor for gibberellin signaling in an F-box mutant[J]. Science, 2003, 299(5614):1896-1898 doi: 10.1126/science.1081077 |
| [53] | POTUSCHAK T, LECHNER E, PARMENTIER Y, et al. EIN3-dependent regulation of plant ethylene hormone signaling by two Arabidopsis F box proteins:EBF1 and EBF2[J]. Cell, 2003, 115(6):679-689 doi: 10.1016/S0092-8674(03)00968-1 |
| [54] | GUO H W, ECKER J R. Plant responses to ethylene gas are mediated by SCF(EBF1/EBF2)-dependent proteolysis of EIN3 transcription factor[J]. Cell, 2003, 115(6):667-677 doi: 10.1016/S0092-8674(03)00969-3 |
| [55] | An F Y, Zhao Q, Ji Y S, et al. Ethylene-induced stabilization of ETHYLENE INSENSITIVE3 and EIN3-LIKE1 is mediated by proteasomal degradation of EIN3 binding F-box 1 and 2 that requires EIN2 in Arabidopsis[J]. The Plant Cell, 2010, 22(7):2384-2401 doi: 10.1105/tpc.110.076588 |
| [56] | QIAO H, CHANG K N, YAZAKI J, et al. Interplay between ethylene, ETP1/ETP2 F-box proteins, and degradation of EIN2 triggers ethylene responses in Arabidopsis[J]. Genes & Development, 2009, 23(4):512-521 https://www.researchgate.net/profile/Junshi_Yazaki/publication/23983018_Interplay_between_ethylene_ETP1ETP2_F-box_proteins_and_degradation_of_EIN2_triggers_ethylene_responses_in_Arabidopsis/links/5808854d08ae63c48fec8377.pdf?origin=publication_detail |
| [57] | QIAO H, SHEN Z X, HUANG S C, et al. Processing and subcellular trafficking of ER-tethered EIN2 control response to ethylene gas[J]. Science, 2012, 338(6105):390-393 doi: 10.1126/science.1225974 |
| [58] | STIRNBERG P, FURNER I J, LEYSER H M O, et al. MAX2 participates in an SCF complex which acts locally at the node to suppress shoot branching[J]. The Plant Journal, 2007, 50(1):80-94 doi: 10.1111/j.1365-313X.2007.03032.x |
| [59] | WANG Y, SUN S Y, ZHU W J, et al. Strigolactone/MAX2-induced degradation of brassinosteroid transcriptional effector BES1 regulates shoot branching[J]. Developmental Cell, 2013, 27(6):681-688 doi: 10.1016/j.devcel.2013.11.010 |
| [60] | CHEVALIER F, NIEMINEN K, SáNCHEZ-FERRERO J C, et al. Strigolactone promotes degradation of dwarf14, an α/β hydrolase essential for strigolactone signaling in Arabidopsis[J]. The Plant Cell, 2014, 26(3):1134-1150 doi: 10.1105/tpc.114.122903 |
| [61] | JIANG L, LIU X, XIONG G S, et al. DWARF 53 acts as a repressor of strigolactone signalling in rice[J]. Nature, 2013, 504(7480):401-405 doi: 10.1038/nature12870 |
| [62] | ZHOU F, LIN Q B, ZHU L H, et al. D14-SCFD3-dependent degradation of D53 regulates strigolactone signalling[J]. Nature, 2013, 504(7480):406-410 doi: 10.1038/nature12878 |
| [63] | BEVERIDGE C A, ROSS J J, MURFET I C. Branching in pea (action of genes Rms3 and Rms4)[J]. Plant Physiology, 1996, 110(3):859-865 doi: 10.1104/pp.110.3.859 |
| [64] | DRUMMOND R S M, SHEEHAN H, SIMONS J L, et al. The expression of petunia strigolactone pathway genes is altered as part of the endogenous developmental program[J]. Frontiers in Plant Science, 2012, 2:115 https://www.researchgate.net/profile/Revel_Drummond/publication/225074666_The_Expression_of_Petunia_Strigolactone_Pathway_Genes_is_Altered_as_Part_of_the_Endogenous_Developmental_Program/links/00b7d5295173aabd76000000.pdf |
| [65] | HAMIAUX C, DRUMMOND R S M, JANSSEN B J, et al. DAD2 is an α/β hydrolase likely to be involved in the perception of the plant branching hormone, strigolactone[J]. Current Biology, 2012, 22(21):2032-2036 doi: 10.1016/j.cub.2012.08.007 |
| [66] | LI Y, ZHANG L, LI D K, et al. The Arabidopsis F-box E3 ligase RIFP1 plays a negative role in abscisic acid signalling by facilitating ABA receptor RCAR3 degradation[J]. Plant, Cell & Environment, 2016, 39(3):571-582 |
| [67] | JIA F J, WANG C Y, HUANG J G, et al. SCF E3 ligase PP2-B11 plays a positive role in response to salt stress in Arabidopsis[J]. Journal of Experimental Botany, 2015, 66(15):4683-4697 doi: 10.1093/jxb/erv245 |
| [68] | CHENG C H, WANG Z J, REN Z Y, et al. SCFAtPP2-B11 modulates ABA signaling by facilitating SnRK2. 3 degradation in Arabidopsis thaliana[J]. PLoS Genetics, 2017, 13(8):e1006947 doi: 10.1371/journal.pgen.1006947 |
| [69] | LI Y Z, JIA F J, YU Y L, et al. The SCF E3 ligase AtPP2-B11 plays a negative role in response to drought stress in Arabidopsis[J]. Plant Molecular Biology Reporter, 2014, 32(5):943-956 doi: 10.1007/s11105-014-0705-5 |
| [70] | ZHANG Y E, XU W Y, LI Z H, et al. F-box protein DOR functions as a novel inhibitory factor for abscisic acid-induced stomatal closure under drought stress in Arabidopsis[J]. Plant Physiology, 2008, 148(4):2121-2133 doi: 10.1104/pp.108.126912 |
| [71] | KOOPS P, PELSER S, IGNATZ M, et al. EDL3 is an F-box protein involved in the regulation of abscisic acid signalling in Arabidopsis thaliana[J]. Journal of Experimental Botany, 2011, 62(15):5547-5560 doi: 10.1093/jxb/err236 |
| [72] | LAI C P, LEE C L, CHEN P H, et al. Molecular analyses of the Arabidopsis TUBBY-like protein gene family[J]. Plant Physiology, 2004, 134(4):1586-1597 doi: 10.1104/pp.103.037820 |
| [73] | PENG J, YU D S, WANG L Q, et al. Arabidopsis F-box gene FOA1 involved in ABA signaling[J]. Science China Life Sciences, 2012, 55(6):497-506 doi: 10.1007/s11427-012-4332-9 |
| [74] | KIM Y Y, CUI M H, NOH M S, et al. The FBA motif-containing protein AFBA1 acts as a novel positive regulator of ABA response in Arabidopsis[J]. Plant &Cell Physiology, 2017, 58(3):574-586 |
| [75] | CHENG Y T, LI Y, HUANG S, et al. Stability of plant immune-receptor resistance proteins is controlled by SKP1-CULLIN1-F-box (SCF)-mediated protein degradation[J]. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(35):14694-14699 doi: 10.1073/pnas.1105685108 |
| [76] | GOU M Y, SHI Z Y, ZHU Y, et al. The F-box protein CPR1/CPR30 negatively regulates R protein SNC1 accumulation[J]. The Plant Journal, 2012, 69(3):411-420 doi: 10.1111/tpj.2012.69.issue-3 |
| [77] | GOU M Y, SU N, ZHENG J, et al. An F-box gene, CPR30, functions as a negative regulator of the defense response in Arabidopsis[J]. The Plant Journal:For Cell And Molecular Biology, 2009, 60(5):757-770 doi: 10.1111/tpj.2009.60.issue-5 |
| [78] | HEDTMANN C, GUO W, REIFSCHNEIDER E, et al. The plant immunity regulating F-Box protein CPR1 supports plastid function in absence of pathogens[J]. Frontiers in Plant Science, 2017, 8:1650 doi: 10.3389/fpls.2017.01650 |
| [79] | ZHU J Y, LI Y Y, CAO D M, et al. The F-box protein KIB1 mediates brassinosteroid-induced inactivation and degradation of GSK3-like kinases in Arabidopsis[J]. Molecular Cell, 2017, 66(5):648-657 doi: 10.1016/j.molcel.2017.05.012 |
| [80] | XU J N, XING S S, CUI H R, et al. Genome-wide identification and characterization of the apple (Malus domestica) HECT ubiquitin-protein ligase family and expression analysis of their responsiveness to abiotic stresses[J]. Molecular Genetics and Genomics, 2016, 291(2):635-646 doi: 10.1007/s00438-015-1129-0 |
| [81] | MUNNS R, TESTER M. Mechanisms of salinity tolerance[J]. Annual Review of Plant Biology, 2008, 59:651-681 doi: 10.1146/annurev.arplant.59.032607.092911 |
| [82] | PENG J Y, LI Z H, WEN X, et al. Salt-induced stabilization of EIN3/EIL1 confers salinity tolerance by deterring ROS accumulation in Arabidopsis[J]. PLoS Genetics, 2014, 10(10):e1004664 doi: 10.1371/journal.pgen.1004664 |
| [83] | QUAN R D, WANG J, YANG D X, et al. EIN3 and SOS2 synergistically modulate plant salt tolerance[J]. Scientific Reports, 2017, 7:44637 doi: 10.1038/srep44637 |
| [84] | VAN HA C, LEYVA-GONZáLEZ M A, OSAKABE Y, et al. Positive regulatory role of strigolactone in plant responses to drought and salt stress[J]. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(2):851-856 doi: 10.1073/pnas.1322135111 |
| [85] | AN X H, HAO Y J, LI E M, et al. Functional identification of apple MdJAZ2 in Arabidopsis with reduced JA-sensitivity and increased stress tolerance[J]. Plant Cell Reports, 2017, 36(2):255-265 doi: 10.1007/s00299-016-2077-9 |
| [86] | BU Q Y, LV T X, SHEN H, et al. Regulation of drought tolerance by the F-box protein MAX2 in Arabidopsis[J]. Plant Physiology, 2014, 164(1):424-439 doi: 10.1104/pp.113.226837 |
| [87] | AN J P, LI R, QU F J, et al. Apple F-Box protein MdMAX2 regulates plant photomorphogenesis and stress response[J]. Frontiers in Plant Science, 2016, 7:1685 https://www.ncbi.nlm.nih.gov/pubmed/27909441 |
| [88] | Zhou S M, Sun X D, Yin S H, et al. The role of the F-box gene TaFBA1 from wheat (Triticum aestivum L.) in drought tolerance[J]. Plant Physiology and Biochemistry, 2014, 84:213-223 doi: 10.1016/j.plaphy.2014.09.017 |
| [89] | ZHOU S M, KONG X Z, KANG H H, et al. The involvement of wheat F-box protein gene TaFBA1 in the oxidative stress tolerance of plants[J]. PLoS One, 2015, 10(4):e0122117 doi: 10.1371/journal.pone.0122117 |
| [90] | KONG X Z, ZHOU S M, YIN S H, et al. Stress-inducible expression of an F-box gene TaFBA1 from wheat enhanced the drought tolerance in transgenic tobacco plants without impacting growth and development[J]. Frontiers in Plant Science, 2016, 7:1295 |
| [91] | ZHAO Z X, ZHANG G Q, ZHOU S M, et al. The improvement of salt tolerance in transgenic tobacco by overexpression of wheat F-box gene TaFBA1[J]. Plant Science, 2017, 259:71-85 doi: 10.1016/j.plantsci.2017.03.010 |
| [92] | JIANG B C, SHI Y T, ZHANG X Y, et al. PIF3 is a negative regulator of the CBF pathway and freezing tolerance in Arabidopsis[J]. Proceedings of the National Academy of Sciences of the United States of America, 2017, 114(32):E6695-E6702 doi: 10.1073/pnas.1706226114 |
| [93] | GONZALEZ L E, KELLER K, CHAN K X, et al. Transcriptome analysis uncovers Arabidopsis F-BOX STRESS INDUCED 1 as a regulator of jasmonic acid and abscisic acid stress gene expression[J]. BMC Genomics, 2017, 18(1):533 doi: 10.1186/s12864-017-3864-6 |
| [94] | CALDERóN-VILLALOBOS L I A, NILL C, MARROCCO K, et al. The evolutionarily conserved Arabidopsis thaliana F-box protein AtFBP7 is required for efficient translation during temperature stress[J]. Gene, 2007, 392(1/2):106-116 https://www.deepdyve.com/lp/elsevier/the-evolutionarily-conserved-arabidopsis-thaliana-f-box-protein-atfbp7-lFiLNE7n2B |
| [95] | BEVILACQUA C B, BASU S, PEREIRA A, et al. Analysis of stress-responsive gene expression in cultivated and weedy rice differing in cold stress tolerance[J]. PLoS One, 2015, 10(7):e0132100 doi: 10.1371/journal.pone.0132100 |
| [96] | ZHANG C K, LANG P, DANE F, et al. Cold acclimation induced genes of trifoliate orange (Poncirus trifoliata)[J]. Plant Cell Reports, 2005, 23(10/11):764-769 |
| [97] | CHEN R G, GUO W L, YIN Y X, et al. A novel F-box protein CaF-box is involved in responses to plant hormones and abiotic stress in pepper (Capsicum annuum L.)[J]. International journal of molecular sciences, 2014, 15(2):2413-2430 doi: 10.3390/ijms15022413 |
| [98] | HUANG Y, MINAKER S, ROTH C, et al. An E4 ligase facilitates polyubiquitination of plant immune receptor resistance proteins in Arabidopsis[J]. The Plant Cell, 2014, 26(1):485-496 doi: 10.1105/tpc.113.119057 |
| [99] | RALHAN A, SCH?TTLE S, THUROW C, et al. The vascular pathogen Verticillium longisporum requires a jasmonic acid-independent COI1 function in roots to elicit disease symptoms in Arabidopsis shoots[J]. Plant Physiology, 2012, 159(3):1192-1203 doi: 10.1104/pp.112.198598 |
| [100] | AZEVEDO C, SADANANDOM A, KITAGAWA K, et al. The RAR1 interactor SGT1, an essential component of R gene-triggered disease resistance[J]. Science, 2002, 295(5562):2073-2076 doi: 10.1126/science.1067554 |
| [101] | AUSTIN M J, MUSKETT P, KAHN K, et al. Regulatory role of SGT1 in early R gene-mediated plant defenses[J]. Science, 2002, 295(5562):2077-2080 doi: 10.1126/science.1067747 |
| [102] | LLORENTE F, MUSKETT P, SáNCHEZ-VALLET A, et al. Repression of the auxin response pathway increases Arabidopsis susceptibility to necrotrophic fungi[J]. Molecular Plant, 2008, 1(3):496-509 doi: 10.1093/mp/ssn025 |
| [103] | PⅡSIL? M, KECELI M A, BRADER G, et al. The F-box protein MAX2 contributes to resistance to bacterial phytopathogens in Arabidopsis thaliana[J]. BMC Plant Biology, 2015, 15:53 doi: 10.1186/s12870-015-0434-4 |
| [104] | Cao Y F, Yang Y Y, Zhang H J, et al. Overexpression of a rice defense-related F-box protein gene OsDRF1 in tobacco improves disease resistance through potentiation of defense gene expression[J]. Physiologia Plantarum, 2008, 134(3):440-452 doi: 10.1111/ppl.2008.134.issue-3 |
| [105] | VAN DEN BURG H A, TSITSIGIANNIS D I, ROWLAND O, et al. The F-box protein ACRE189/ACIF1 regulates cell death and defense responses activated during pathogen recognition in tobacco and tomato[J]. The Plant Cell, 2008, 20(3):697-719 doi: 10.1105/tpc.107.056978 |
| [106] | KIM H S, DELANEY T P. Arabidopsis SON1 is an F-box protein that regulates a novel induced defense response independent of both salicylic acid and systemic acquired resistance[J]. The Plant Cell, 2002, 14(7):1469-1482 doi: 10.1105/tpc.001867 |
| [107] | THIEL H, HLEIBIEH K, GILMER D, et al. The P25 pathogenicity factor of Beet necrotic yellow vein virus targets the sugar beet 26S proteasome involved in the induction of a hypersensitive resistance response via interaction with an F-box protein[J]. Molecular Plant-Microbe Interactions:MPMI, 2012, 25(8):1058-1072 doi: 10.1094/MPMI-03-12-0057-R |
| [108] | BAUMBERGER N, TSAI C H, LIE M, et al. The polerovirus silencing suppressor P0 targets ARGONAUTE proteins for degradation[J]. Current Biology, 2007, 17(18):1609-1614 doi: 10.1016/j.cub.2007.08.039 |
| [109] | BORTOLAMIOL D, PAZHOUHANDEH M, MARROCCO K, et al. The polerovirus F Box protein P0 targets ARGONAUTE1 to suppress RNA silencing[J]. Current Biology, 2007, 17(18):1615-1621 doi: 10.1016/j.cub.2007.07.061 |
| [110] | FUSARO A F, CORREA R L, NAKASUGI K, et al. The Enamovirus P0 protein is a silencing suppressor which inhibits local and systemic RNA silencing through AGO1 degradation[J]. Virology, 2012, 426(2):178-187 doi: 10.1016/j.virol.2012.01.026 |
| [111] | LI H O, YAO R F, MA S, et al. Efficient ASK-assisted system for expression and purification of plant F-box proteins[J]. The Plant Journal:for Cell and Molecular Biology, 2017, 92(4):736-743 doi: 10.1111/tpj.2017.92.issue-4 |
