许梦洁1, 3,
李霞1,,
1.江苏省农业科学院粮食作物研究所/江苏省优质水稻工程技术研究中心/国家水稻改良中心南京分中心 南京 210014
2.南京林业大学生物与环境学院 南京 210037
3.南京晓庄学院 南京 211171
基金项目: 国家自然科学基金项目31571585
江苏省农业科学院基本科研业务专项ZX[16]2002
江苏省农业科学院粮食作物研究所基金LZS17-9
详细信息
作者简介:何亚飞, 主要研究方向为作物逆境生理。E-mail:1368267588@qq.com
通讯作者:李霞, 主要研究方向为植物光合生理。E-mail:jspplx@jaas.ac.cn
中图分类号:S311;S338计量
文章访问数:1059
HTML全文浏览量:3
PDF下载量:1099
被引次数:0
出版历程
收稿日期:2017-07-17
录用日期:2017-08-22
刊出日期:2018-03-01
Effects of DCMU on anthocyanin synthesis genes and its related signals in C4-pepc gene overexpressed rice under drought conditions
HE Yafei1, 2,,XU Mengjie1, 3,
LI Xia1,,
1. Institute of Food Crops, Jiangsu Academy of Agricultural Sciences/Jiangsu High Quality Rice Engineering Technology Research Center/Nanjing Branch of National Center for Rice Improvement, Nanjing 210014, China
2. College of Biology and Environment, Nanjing Forestry University, Nanjing 210037, China
3. Nanjing Xiaozhuang University, Nanjing 211171, China
Funds: the National Natural Science Foundation of China31571585
the Special Project for Basic Research of Jiangsu Academy of Agricultural SciencesZX[16]2002
the Grant from the Institute of Food Crops of Jiangsu Academy of Agriculture SciencesLZS17-9
More Information
Corresponding author:LI Xia, E-mail:jspplx@jaas.ac.cn
摘要
HTML全文
图
参考文献
相关文章
施引文献
资源附件
访问统计
摘要
摘要:为了揭示高表达转玉米C4-磷酸烯醇式丙酮酸羧化酶(PEPC,EC 4.1.1.31)基因水稻(PC)在耐旱中光合与花青素调节途径的内在联系,本研究以PC和未转基因野生型原种(WT)的水培苗为试验材料,在4~5叶期,通过50 μmol·L-1光合抑制剂DCMU[3-(3',4'-dich-lorophenyl)-1,1-dimethyl-urea]预处理1 h,观察其在12% PEG-6000模拟干旱处理下的表现。结果表明,在模拟干旱条件下,DCMU预处理使两种供试材料相对含水量显著下降,且PC相对含量显著高于WT;干旱处理下,两种材料的花青素含量显著升高,DCMU和干旱处理使两种材料的花青素含量下调,且PC水稻中始终伴随着较高的花青素含量。光合数据表明,与单独12% PEG-6000处理相比,DCMU联合12% PEG-6000处理显著抑制了两种水稻材料的净光合速率、气孔导度、胞间CO2含量及羧化效率,但PC的各指标显著高于WT。同时,DCMU联合12% PEG-6000处理显著下调两种供试材料的内源蔗糖含量,但PC中蔗糖含量显著高于WT。进一步研究发现PC中更高的蔗糖含量与花青素合成有关转录因子bHLH(OsB1,OsB2)、R2R3-MYB(OsC1)、COP1(constitutively photomorphogenic 1)、HY5(elongated hypocotyl 5)更高的转录水平同步,下游花青素合成相关基因OsPAL、OsCHI、OsCHS、OsF3H、OsF3'H、OsDFR、OsANS的表达量增加。PC水稻可能通过诱导NO和Ca2+感受干旱信号,参与转录因子的调节,进而参与花青素合成基因的调控,合成较多的花青素,增强PC水稻对干旱逆境的响应,增强保水能力,最终表现耐旱。
Abstract:Anthocyanins are important antioxidant materials that protects plant from damage by reactive oxygen species (ROS). Especially under adverse conditions, the regulation of sucrose in plants depends on its ability to induce anthocyanin accumulation. To determine the intrinsic relationship between photosynthetic and anthocyanin regulated pathways for C4-phosphoenolpyruvate carboxylate (PEPC, EC 4.1.1.31) gene overexpressed rice (PC) in drought conditions, PC and untransformed wild-type (WT) were treated with 50 μmol·L-1 photosynthetic inhibitor DCMU for 1 h and the performance of the rice seedlings at 4-5 leaf stage observed under 12% PEG-6000 simulated drought. The results showed that DCMU pretreatment significantly reduced relative water contents of WT and PC under simulated 12% PEG-6000 drought condition, and relative water content of PC was significantly higher than that of WT. The anthocyanin content was higher in PC than in WT under 12% PEG-6000 simulated drought or drought plus DCMU pretreatment. 12% PEG-6000 simulated drought decreased anthocyanin contents of PC and WT, while DCMU pretreatment alleviated this effect. Compared with 12% PEG-6000, DCMU plus 12% PEG-6000 significantly inhibited net photosynthetic rate, stomatal conductance, intercellular CO2 and carboxylation efficiency of the two rice lines, but these parameters of PC lines were significantly higher than those of WT lines. Then DCMU plus 12% PEG-6000 down-regulated endogenous sucrose content of the two materials, but sucrose content of PC lines was significantly higher than that of WT lines. Further studies showed that higher sucrose level in PC was associated with higher expression levels of transcriptional factors of bHLH (OsB1, OsB2), R2R3-MYB (OsC1), COP1 (constitutively photomorphogenic 1), HY5 (elongated hypocotyl 5), OsPAL, OsCHI, OsCHS, OsF3H, OsF3'H, OsDFR and OsANS, which resulted in synthesizing more anthocyanin to improve water retention capacity. In addition, PC rice sensed drought signals through NO and Ca2+, which participated in the regulation of transcription factors, regulation of anthocyanin synthesis gene, synthesis of more anthocyanin and thereby enhanced PC rice response to drought stress. This enhanced water retention capacity, stabilized photosynthetic capacity and resisted drought. Therefore, it was beneficial in molecular breeding of "C4 Rice" to study the symphony between high yield and plant resistance.
HTML全文
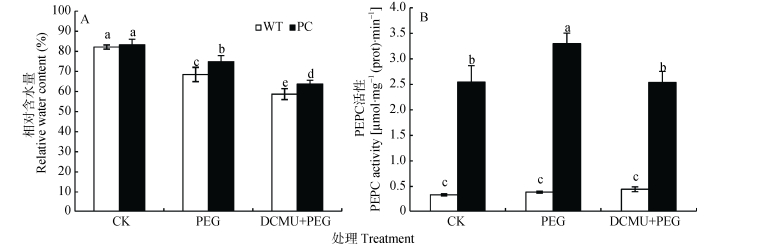
图1DCMU预处理对模拟干旱胁迫下高表达转C4-pepc基因水稻(PC)和野生型水稻(WT)叶片相对含水量(A)和PEPC活性(B)的影响
CK:正常水培培养; PEG: PEG单独施加模拟干旱处理; DCMU+PEG: DCMU预处理之后联合模拟干旱胁迫处理。不同小写字母表示WT和PC的不同处理间差异显著(P<0.05)。
Figure1.Effects of DCMU pretreatment on leaves relative water content (A) and PEPC activity (B) of C4-pepc gene overexpressed rice (PC) and untransformed wild-type rice (WT) under simulated drought stress
CK: normal hydroponic culture; PEG: PEG simulated drought stress; DCMU+PEG: DCMU pretreatment plus simulated drought stress. Different lowercase letters indicate significant differences among different treatments of WT and PC at P < 0.05.
 下载: 全尺寸图片幻灯片
下载: 全尺寸图片幻灯片
图2DCMU预处理对模拟干旱胁迫下高表达转C4-pepc基因水稻(PC)和野生型水稻(WT)叶片光合参数(A:净光合速率; B:气孔导度; C:胞间CO2浓度; D:羧化效率)的影响
CK:正常水培培养; PEG: PEG单独施加模拟干旱处理; DCMU+PEG: DCMU预处理之后联合模拟干旱胁迫处理。不同小写字母表示WT和PC的不同处理间差异显著(P<0.05, LSD test)。
Figure2.Effects of DCMU pretreatment on photosynthetic parameters (A: net photosynthetic rate; B: stomatal conductance; C: intercellular CO2 concentration; D: carboxylation efficiency) of C4-pepc gene overexpressed rice (PC) and untransformed wild-type rice (WT) leaves under simulated drought stress
CK: normal hydroponic culture; PEG: PEG simulated drought stress; DCMU+PEG: DCMU pretreatment plus simulated drought stress. Different lowercase letters indicate significant differences among different treatments of WT and PC at P < 0.05.
下载: 全尺寸图片幻灯片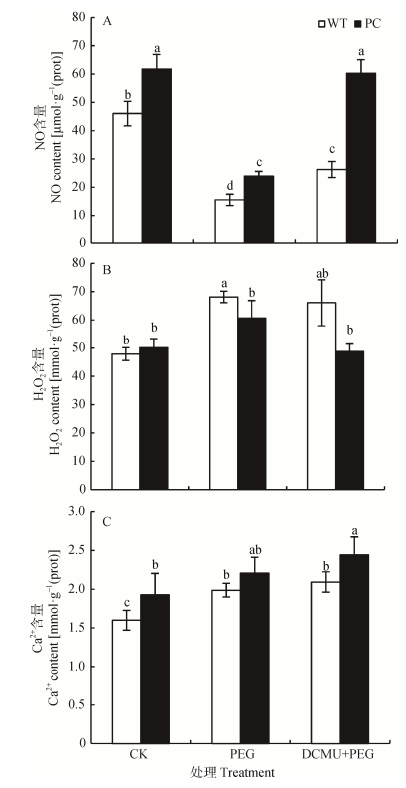
图3DCMU预处理对模拟干旱胁迫下高表达转C4-pepc基因水稻(PC)和野生型水稻(WT)叶片NO (A)、H2O2 (B)和Ca2+ (C)含量的影响
CK:正常水培培养; PEG: PEG单独施加模拟干旱处理; DCMU+PEG: DCMU预处理之后联合模拟干旱胁迫处理。不同小写字母表示WT和PC的不同处理间差异显著(P<0.05, LSD test)。
Figure3.Effects of DCMU pretreatment on contents of NO (A), H2O2 (B) and Ca2+ (C) of C4-pepc gene overexpressed rice (PC) and untransformed wild-type rice (WT) leaves under simulated drought stress
CK: normal hydroponic culture; PEG: PEG simulated drought stress; DCMU+PEG: DCMU pretreatment plus simulated drought stress. Different lowercase letters indicate significant differences among different treatments of WT and PC at P < 0.05.
下载: 全尺寸图片幻灯片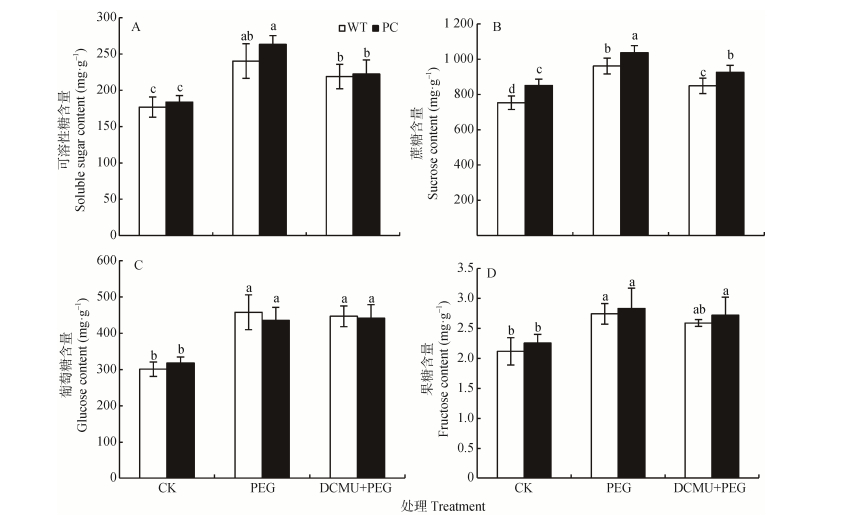
图4DCMU预处理对模拟干旱胁迫下高表达转C4-pepc基因水稻(PC)和野生型水稻(WT)叶片可溶性糖(A)、蔗糖(B)、葡萄糖(C)和果糖(D)含量的影响
CK:正常水培培养; PEG: PEG单独施加模拟干旱处理; DCMU+PEG: DCMU预处理之后联合模拟干旱胁迫处理。不同小写字母表示WT和PC的不同处理间差异显著(P<0.05)。
Figure4.Effects of DCMU pretreatment on contents of soluble sugar (A), sucrose (B), glucose (C) and fructose (D) of C4-pepc gene overexpressed rice (PC) and untransformed wild-type rice (WT) leaves under simulated drought stresses
CK: normal hydroponic culture; PEG: PEG simulated drought stress; DCMU+PEG: DCMU pretreatment plus simulated drought stress. Different lowercase letters indicate significant differences among different treatments of WT and PC at P < 0.05.
下载: 全尺寸图片幻灯片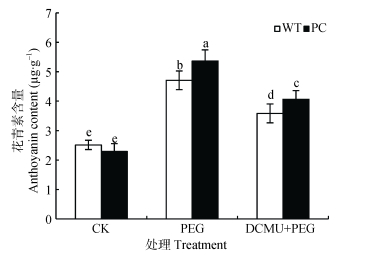
图5DCMU预处理对模拟干旱胁迫下高表达转C4-pepc基因水稻(PC)和野生型水稻(WT)叶片花青素含量影响
CK:正常水培培养; PEG: PEG单独施加模拟干旱处理; DCMU+PEG: DCMU预处理之后联合模拟干旱胁迫处理。不同小写字母表示WT和PC的不同处理间差异显著(P<0.05)。
Figure5.Effect of DCMU pretreatment on anthocyanin content of C4-pepc gene overexpressed rice (PC) and untransformed wild-type rice (WT) leaves under simulated drought stress
CK: normal hydroponic culture; PEG: PEG simulated drought stress; DCMU+PEG: DCMU pretreatment plus simulated drought stress. Different lowercase letters indicate significant differences among different treatments of WT and PC at P < 0.05.
下载: 全尺寸图片幻灯片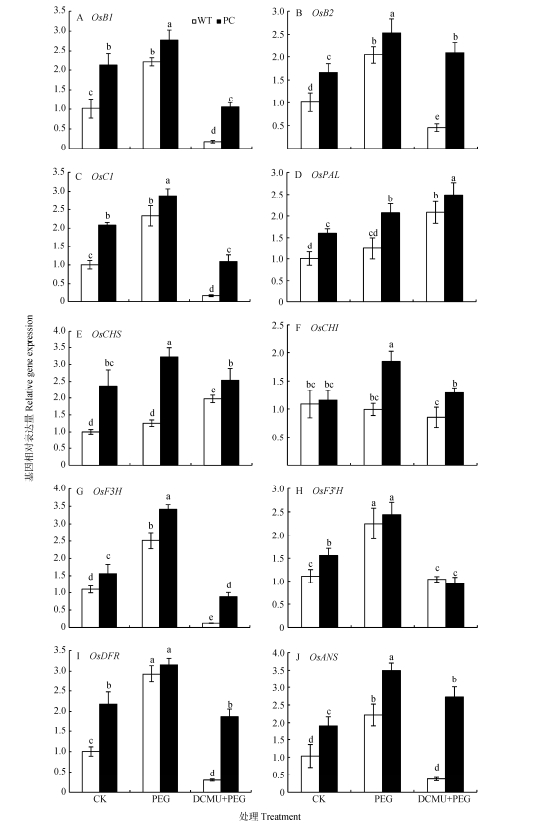
图6DCMU预处理对模拟干旱胁迫下高表达转C4-pepc基因水稻(PC)和野生型水稻(WT)叶片花青素合成酶基因表达的影响
CK:正常水培培养; PEG: PEG单独施加模拟干旱处理; DCMU+PEG: DCMU预处理之后联合模拟干旱胁迫处理。不同小写字母表示WT和PC的不同处理间差异显著(P<0.05, LSD test)。
Figure6.Effects of DCMU pretreatment on expression of anthocyanin synthase genes of C4-pepc gene overexpressed rice (PC) and untransformed wild-type rice (WT) leaves under simulated drought stress
CK: normal hydroponic culture; PEG: PEG simulated drought stress; DCMU+PEG: DCMU pretreatment plus simulated drought stress. Different lowercase letters indicate significant differences among different treatments of WT and PC at P < 0.05.
下载: 全尺寸图片幻灯片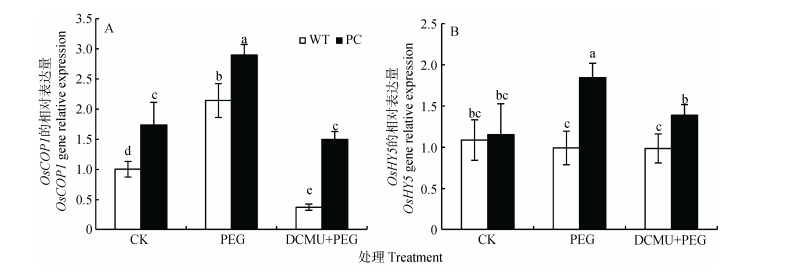
图7DCMU预处理对模拟干旱胁迫下高表达转C4-pepc基因水稻(PC)和野生型水稻(WT)叶片花青素调节因子的影响
CK:正常水培培养; PEG: PEG单独施加模拟干旱处理; DCMU+PEG: DCMU预处理之后联合模拟干旱胁迫处理。不同小写字母表示WT和PC的不同处理间差异显著(P<0.05, LSD test)。
Figure7.Effect of DCMU pretreatment on regulators of anthocyanin of C4-pepc gene overexpressed rice (PC) and untransformed wild-type rice (WT) leaves under simulated drought stress
CK: normal hydroponic culture; PEG: PEG simulated drought stress; DCMU+PEG: DCMU pretreatment plus simulated drought stress. Different lowercase letters indicate significant differences among different treatments of WT and PC at P < 0.05.
下载: 全尺寸图片幻灯片表1QRT-PCR的基因和引物
Table1.Genes and primers for QRT-PCR
| 基因?Gene | 引物序列?Primers sequence | |||
| 基因描述 Gene description | 引物名称 Primers name | 正向引物 Forward primer (F) | 反向引物 Reverse primer (R) | |
| bHLH | ACTIN | CCCTCTTTCATCGGTATGGA | TTGATCTTCATGCTGCTTGG | |
| B1 | GGATGGTCTCCTTGGACTGA | GGGTGGCAGATTCATCACTT | ||
| B2 | GTGGCAATAACGACGACGACTCC | CGTACGGTGTTGACGAGGTA | ||
| MYB | C1 | CGGGTTCTTCTTCCACGAC | CCCGCAACTGCACTTAAAAT | |
| Anthocyanin synthase | PAL | CAAGCTCATGACCTCCACCTA | GTTCATGGTGAGCACCTTCTT | |
| gene | CHS | TCATGTATGGGTGGTTTGGTT | GCCAGGCATCTCTTACACAGG | |
| CHI | CGAGCAGTACTCGGACAAGG | TGAAGGCCTCCTTGAACTTG | ||
| F3H | GAGCAATGGGAGGTTCAAGA | CTTCGATTTTCGACGGAAGA | ||
| F3’H | CCGCTACAGTACCAGCCTTC | TGCCACCATTTCTAGAGTTCC | ||
| Transcription factor | DFR | CGGATGGATGTACTTCGTGTC | CATCCCGTTGCTGATGAAG | |
| ANS | CTCCTCCAGCTCAAGATCAAC | GTTGTGGAGGATGAAGGAGAG | ||
| COP1 | ATAATCCTGGGTCGAGCCAC | TATGGTGATCAGCAGAACCCAC | ||
| HY5 | GGCGGGTGCCGGAGATGG | CGCCGTCGTGTTCTTCTTGAGTATCTGG | ||
下载: 导出CSV参考文献
| [1] | CHAVES M M, FLEXAS J, PINHEIRO C. Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell[J]. Annals of Botany, 2009, 103(4): 551-560 doi: 10.1093/aob/mcn125 |
| [2] | ZHU X G, LONG S P, ORT D R. Improving photosynthetic efficiency for greater yield[J]. Annual Review of Plant Biology, 2010, 61: 235-261 doi: 10.1146/annurev-arplant-042809-112206 |
| [3] | KARKI S, RIZAL G, QUICK W P. Improvement of photosynthesis in rice (Oryza sativa L.) by inserting the C4 pathway[J]. Rice, 2013, 6(1): 28 doi: 10.1186/1939-8433-6-28 |
| [4] | O'LEARY B, PARK J, PLAXTON W C. The remarkable diversity of plant PEPC (phosphoenolpyruvate carboxylase): recent insights into the physiological functions and post-translational controls of non-photosynthetic PEPCs[J]. Biochemical Journal, 2011, 436(1): 15-34 doi: 10.1042/BJ20110078 |
| [5] | KU M S B, AGARIE S, NOMURA M, et al. High-level expression of maize phosphoenolpyruvate carboxylase in transgenic rice plants[J]. Nature Biotechnology, 1999, 17: 76-80 doi: 10.1038/5256 |
| [6] | JEANNEAU M, GERENTES D, FOUEILLASSAR X, et al. Improvement of drought tolerance in maize: towards the functional validation of the Zm-Asr1 gene and increase of water use efficiency by over-expressing C4-PEPC[J]. Biochimie, 2002, 84: 1127-1135 doi: 10.1016/S0300-9084(02)00024-X |
| [7] | JIAO D M, KUANG T Y, LI X, et al. Physiological characteristics of the primitive CO2 concentrating mechanism in PEPC transgenic rice[J]. Science in China Series C: Life Sciences, 2003, 46(4): 438-446 doi: 10.1360/02yc0075 |
| [8] | BANDYOPADHYAY A, DATTA K, ZHANG J, et al. Enhanced photosynthesis rate in genetically engineered indica rice expressing PEPC gene cloned from maize[J]. Plant Science, 2007, 172(6): 1204-1209 doi: 10.1016/j.plantsci.2007.02.016 |
| [9] | LIAN L, WANG X W, ZHU Y S, et al. Physiological and photosynthetic characteristics of indica Hang2 expressing the sugarcane PEPC gene[J]. Molecular Biology Reports, 2014, 41: 2189-2197 doi: 10.1007/s11033-014-3070-4 |
| [10] | REN C G, LI X, LIU X L, et al. Hydrogen peroxide regulated photosynthesis in C4-pepc transgenic rice[J]. Plant Physiology and Biochemistry, 2014, 74: 218-229 doi: 10.1016/j.plaphy.2013.11.011 |
| [11] | DING Z S, SUN X F, HUANG S H, et al. Response of photosynthesis to short-term drought stress in rice seedlings overexpressing C4 phosphoenolpyruvate carboxylase from maize and millet[J]. Photosynthetica, 2015, 53 (4): 481-488 doi: 10.1007/s11099-015-0126-1 |
| [12] | 钱宝云, 刘小龙, 李霞.钙肥对不同内源钙含量水稻品种光合参数的影响[J].江苏农业学报, 2014, 30(3): 467-473 http://www.cnki.com.cn/Article/CJFDTOTAL-JSNB201403003.htm QIAN B Y, LIU X L, LI X. Photosynthesis of rice cultivars with various endogenous calcium contents in response to calcium fertilizer application[J]. Jiangsu Journal of Agricultural Sciences, 2014, 30(3): 467-473 http://www.cnki.com.cn/Article/CJFDTOTAL-JSNB201403003.htm |
| [13] | 霍垲, 陆巍, 李霞.干旱胁迫下调节ATP的含量对提高转玉米C4型pepc水稻光合速率的影响[J].中国生态农业学报, 2015, 23(5): 605-613 http://www.ecoagri.ac.cn/zgstny/ch/reader/view_abstract.aspx?file_no=2015510&flag=1 HUO K, LU W, LI X. Effect of regulating ATP on improving photosynthetic rate of transgenic rice with overexpressing maize C4 pepc under drought stress[J]. Chinese Journal of Eco-Agriculture, 2015, 23(5): 605-613 http://www.ecoagri.ac.cn/zgstny/ch/reader/view_abstract.aspx?file_no=2015510&flag=1 |
| [14] | 唐玉婷, 李霞, 陆巍, 等.高表达转C4型PEPC基因水稻在低氮下诱导碳氮酶稳定光合作用[J].华北农学报, 2015, 30(4): 95-100 doi: 10.7668/hbnxb.2015.04.017 TANG Y T, LI X, LU W, et al. Transgenic rice with high expression of C4-PEPC genes induced higher carbon and nitrogen key enzyme to maintain photosynthesis under low nitrogen conditions[J]. Acta Agriculturae Boreali-Sinica, 2015, 30(4): 95-100 doi: 10.7668/hbnxb.2015.04.017 |
| [15] | JIAO D M, XIA L, HUANG X Q, et al. The characteristics of CO2 assimilation of photosynthesis and chlorophyll fluorescence in transgenic PEPC rice[J]. Science Bulletin, 2001, 46(13): 1080-1084 doi: 10.1007/BF02900682 |
| [16] | DOUBNEROVá V, RY?LAVá H. What can enzymes of C4, photosynthesis do for C3, plants under stress?[J]. Plant Science, 2011, 180(4): 575-583 doi: 10.1016/j.plantsci.2010.12.005 |
| [17] | JIAO D M, LI X, JI B H. Photoprotective effects of high level expression of C4, phosphoenolpyruvate carboxylase in transgenic rice during photoinhibition[J]. Photosynthetica, 2005, 43(4): 501-508 doi: 10.1007/s11099-005-0082-2 |
| [18] | 李霞, 任承钢. ABA、BA及DPI对高表达玉米C4 pepc基因的水稻光合特性及叶绿素荧光特性的影响[J].植物生理学报, 2012, 48(6): 549-556 http://www.cqvip.com/QK/90791A/201206/42213749.html LI X, REN C G. Effects on photosynthetic and fluorescence characteristics under treatments of ABA, BA or DPI in transgenic rice with over-expression C4-pepc gene[J]. Plant Physiology Journal, 2012, 48(6): 549-556 http://www.cqvip.com/QK/90791A/201206/42213749.html |
| [19] | LIU X, LI X, ZHANG C, et al. Phosphoenolpyruvate carboxylase regulation in C4-PEPC-expressing transgenic rice during early responses to drought stress[J]. Physiologia Plantarum, 2017, 159(2): 178-200 doi: 10.1111/ppl.2017.159.issue-2 |
| [20] | HUO K, LI X, HE Y F, et al. Exogenous ATP enhance signal response of suspension cells of transgenic rice (Oryza sativa L.) expressing maize C4-pepc encoded phosphoenolpyruvate carboxylase under PEG treatment[J]. Plant Growth Regulation, 2016, 82(1): 55–67 doi: 10.1007/s10725-016-0238-z |
| [21] | QIAN B Y, LI X, LIU X L, et al. Enhanced drought tolerance in transgenic rice over-expressing of maize C4 phosphoenolpyruvate carboxylase gene via NO and Ca2+[J]. Journal of Plant Physiology, 2015, 175: 9–20 doi: 10.1016/j.jplph.2014.09.019 |
| [22] | QIAN B Y, LI X, LIU X L, et al. Improved oxidative tolerance in suspension-cultured cells of C4-pepc transgenic rice by H2O2 and Ca2+ under PEG-6000[J]. Journal of Integrative Plant Biology, 2015, 57(6): 534–549 doi: 10.1111/jipb.12283 |
| [23] | NAKASHIMA K, TRAN L S P, DONG V N, et al. Functional analysis of a NAC-type transcription factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice[J]. Plant Journal, 2007, 51: 617-630 doi: 10.1111/j.1365-313X.2007.03168.x |
| [24] | NIJHAWAN A, JAIN M, TYAGI A K, et al. Genomic survey and gene expression analysis of the basic leucine zipper transcription factor family in rice[J]. Plant Physiology, 2008, 146(2): 333-350 http://www.plantphysiol.org/content/plantphysiol/146/2/333.full.pdf |
| [25] | HUANG G T, MA S L, BAI L P, et al. Signal transduction during cold, salt, and drought stresses in plants[J]. Molecular Biology Reports, 2012, 39: 969-987 doi: 10.1007/s11033-011-0823-1 |
| [26] | 朱素琴, 季本华, 焦德茂.亚硫酸氢钠对转PEPC基因水稻叶片光合作用的促进作用[J].科技通报, 2004, 20(6): 523-528 http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=kjtb200406012 ZHU S Q, JI B H, JIAO D M. Promotive effect of NaHSO3 on photosynthesis in PEPC transgenic rice leaves[J]. Bulletin of Science and Technology, 2004, 20(6): 523-528 http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=kjtb200406012 |
| [27] | BISWAL A K, DILNAWAZ F, DAVID K A V, et al. Increase in the intensity of thermoluminescence Q-band during leaf ageing is due to a block in the electron transfer from QA to QB[J]. Luminescence, 2001, 16(5): 309-313 doi: 10.1002/(ISSN)1522-7243 |
| [28] | SUZUKI K, OHMORI Y, RATEL E. High root temperature blocks both linear and cyclic electron transport in the dark during chilling of the leaves of rice seedlings[J]. Plant and Cell Physiology, 2011, 52(9): 1697-1707 doi: 10.1093/pcp/pcr104 |
| [29] | VAN DEN ENDE W, EL-ESAWE S K. Sucrose signaling pathways leading to fructan and anthocyanin accumulation: A dual function in abiotic and biotic stress responses?[J]. Environmental and Experimental Botany, 2014, 108: 4-13 doi: 10.1016/j.envexpbot.2013.09.017 |
| [30] | TENG S, KEURENTJES J, BENTSINK L, et al. Sucrose-specific induction of anthocyanin biosynthesis in Arabidopsis requires the MYB75/PAP1 gene[J]. Plant Physiology, 2005, 139(4): 1840-1852 doi: 10.1104/pp.105.066688 |
| [31] | SANYAL S K, PANDEY A, PANDEY G K. The CBL–CIPK signaling module in plants: a mechanistic perspective[J]. Physiologia Plantarum, 2015, 155: 89-108 doi: 10.1111/ppl.2015.155.issue-2 |
| [32] | SHIN D H, CHOI M G, KANG C S, et al. A wheat R2R3-MYB protein PURPLE PLANT1 (TaPL1) functions as a positive regulator of anthocyanin biosynthesis[J]. Biochemical & Biophysical Research Communications, 2016, 469(3): 686-691 https://www.pubfacts.com/detail/26692488/A-wheat-R2R3-MYB-protein-PURPLE-PLANT1-TaPL1-functions-as-a-positive-regulator-of-anthocyanin-biosyn |
| [33] | YASUDA S, AOYAMA S, HASEGAWA Y, et al. Arabidopsis CBL-interacting protein kinases regulate carbon/nitrogen-nutrient response by phosphorylating ubiquitin ligase ATL31[J]. Molecular Plant, 2017, 4(10): 537-658 https://www.sciencedirect.com/science/article/pii/S1674205217300060 |
| [34] | SANYAL S K, KANWAR P, YADAV A K, et al. Arabidopsis CBL interacting protein kinase 3 interacts with ABR1, an APETALA2 domain transcription factor, to regulate ABA responses[J]. Plant Science, 2017, 254: 48-59 doi: 10.1016/j.plantsci.2016.11.004 |
| [35] | YOSHIDA S, FORNO D A, COCK J H, et al. Laboratory Manual for Physiological Studies of Rice[M]. Philippines: International Rice Research Institute, 1976: 61–64 |
| [36] | LI X, WANG C, REN C G. Effects of 1-butanol, neomycin and calcium on the photosynthetic characteristics of PEPC transgenic rice[J]. African Journal of Biotechnology, 2011, 10: 17466-17476 http://www.freepatentsonline.com/9687001.html |
| [37] | AMBAVARAM M M R, BASU S, KRISHNAN A, et al. Coordinated regulation of photosynthesis in rice increases yield and tolerance to environmental stress[J]. Nature Communications, 2014, 5: 1-14 http://www.nature.com/ncomms/2014/141031/ncomms6302/full/ncomms6302.html |
| [38] | RABINO I, MANCINELLI A L. Light, temperature, and anthocyanin production[J]. Plant Physiology, 1986, 81(3): 922-924 doi: 10.1104/pp.81.3.922 |
| [39] | MURPHY M E, NOACK E. Nitric oxide assay using hemoglobin method[J]. Methods in Enzymology, 1994, 233: 240–250 doi: 10.1016/S0076-6879(94)33027-1 |
| [40] | YANG C Q, LIU W N, ZHAO Z H, et al. Determination of the content of serum calcium with methylthymol blue as chromogenic reagent[J]. Spectroscopy & Spectral Analysis, 1998, 18: 485–487 https://www.researchgate.net/publication/7912269_Determination_of_the_content_of_serum_calcium_with_methylthymol_blue_as_chromogenic_reagent |
| [41] | JUNG H, KIM J K, HA S W. Use of animal viral IRES sequence makes multiple truncated transcripts without mediating polycistronic expression in rice[J]. Journal of the Korean Society for Applied Biological Chemistry, 2011, 54: 678–684 doi: 10.1007/BF03253145 |
| [42] | CHEN P B, LI X, HUO K, et al. Promotion of photosynthesis in transgenic rice over-expressing of maize C4 phosphoenolpyruvate carboxylase gene by nitric oxide donors[J]. Journal of Plant Physiology, 2014, 171: 458–466 doi: 10.1016/j.jplph.2013.11.006 |
| [43] | JIA X H, ZHANG P P, SHI D J, et al. Regulation of PEPC gene expression in Anabaena sp. PCC 7120 and its effects on cyclic electron flow around photosystem I and tolerances to environmental stresses[J]. Journal of Integrative Plant Biology, 2015, 57(5): 468-476 doi: 10.1111/jipb.v57.5 |
| [44] | KEUNEN E, PESHEV D, VANGRONSVELD J, et al. Plant sugars are crucial players in the oxidative challenge during abiotic stress: extending the traditional concept[J]. Plant Cell & Environment, 2013, 36(7): 1242-1255 doi: 10.1111/pce.12061/abstract?globalMessage=0 |
| [45] | LORETI E, POVERO G, NOVI G, et al. Gibberellins, jasmonate and abscisic acid modulate the sucrose-induced expression of anthocyanin biosynthetic genes in Arabidopsis[J]. New Phytologist, 2008, 179(4): 1004-1016 doi: 10.1111/nph.2008.179.issue-4 |
| [46] | ZHANG Y C, GONG S F, LI Q H, et al. Functional and signaling mechanism analysis of rice CRYPTOCHROME 1[J]. Plant Journal, 2006, 46(6): 971-983 doi: 10.1111/tpj.2006.46.issue-6 |
| [47] | SHIN D H, CHOI M, KIM K, et al. HY5 regulates anthocyanin biosynthesis by inducing the transcriptional activation of the MYB75/PAP1 transcription factor in Arabidopsis[J]. FEBS letters, 2013, 587(10): 1543-1547 doi: 10.1016/j.febslet.2013.03.037 |
| [48] | 丁在松, 周宝元, 孙雪芳, 等.干旱胁迫下PEPC过表达增强水稻的耐强光能力[J].作物学报, 2012, 38(2): 285-292 http://www.cqvip.com/QK/90181X/201202/40898053.html DING Z S, ZHOU B Y, SUN X F, et al. High light tolerance is enhanced by overexpressed PEPC in rice under drought stress[J]. Acta Agronomica Sinica, 2012, 38(2): 285-292 http://www.cqvip.com/QK/90181X/201202/40898053.html |
| [49] | SANTOS M G, RIBEIRO R V, MACHADO E C, et al. Photosynthetic parameters and leaf water potential of five common bean genotypes under mild water deficit[J]. Biologia Plantarum, 2009, 53(2): 229-236 doi: 10.1007/s10535-009-0044-9 |
| [50] | VAVASSEUR A, RAGHAVENDRA A S. Guard cell metabolism and CO2 sensing[J]. New Phytologist, 2005, 16: 665-682 https://www.researchgate.net/profile/Wagner_Araujo2/publication/51576459_Control_of_stomatal_aperture_A_renaissance_of_the_old_guard/links/00b49528131b52374e000000.pdf?inViewer=true&pdfJsDownload=true&disableCoverPage=true&origin=publication_detail |
| [51] | ZHANG C, LI X, HE Y F, et al. Physiological investigation of C4-phosphoenolpyruvate-carboxylase-introduced rice line shows that sucrose metabolism is involved in the improved drought tolerance[J]. Plant Physiology and Biochemistry, 2017, 115: 328-342 doi: 10.1016/j.plaphy.2017.03.019 |
| [52] | JENKINS C L D. Effects of the phosphoenolpyruvate carboxylase inhibitor 3, 3-dichloro-2-(dihydroxy phosphinoyl methyl) propenoate on photosynthesis. C4 selectivity and studies on C4 photosynthesis[J]. Plant Physiology, 1989, 89(4): 1231-1237 doi: 10.1104/pp.89.4.1231 |
| [53] | AGBARIAH K T, ROTHBEJERANO N. The effect of blue light on energy levels in epidermal strips[J]. Physiologia Plantarum, 1990, 78(1): 100-104 doi: 10.1111/ppl.1990.78.issue-1 |
| [54] | TOMINAGA M, KINOSHITA T, SHIMAZAKI K. Guard-cell chloroplasts provide ATP required for H+ pumping in the plasma membrane and stomatal opening[J]. Plant & Cell Physiology, 2001, 42(8): 795-802 https://www.researchgate.net/publication/11090258_Phosphatidylinositol_3-_and_4-phosphate_are_required_for_normal_stomatal_movements |
| [55] | MENG L S, LI Y Q, LIU M Q, et al. The arabidopsis ANGUSTIFOLIA3-YODA gene cascade induces anthocyanin accumulation by regulating sucrose levels[J]. Frontiers in Plant Science, 2016, 7: 1728 http://www.ncbi.nlm.nih.gov/pmc/articles/PMC5118565/ |
| [56] | SAKAMOTO W, OHMORI T, KAGEYAMA K, et al. The purple leaf (Pl) locus of rice: The Pl(w) allele has a complex organization and includes two genes encoding basic helix-loop-helix proteins involved in anthocyanin biosynthesis[J]. Plant & Cell Physiology, 2001, 42(9): 982-991 https://academic.oup.com/pcp/article/42/9/982/1833701/The-Purple-leaf-Pl-Locus-of-Rice-the-Plw-Allele |
| [57] | DONG W, NIU L L, GU J T, et al. Isolation of a WD40-repeat gene regulating anthocyanin biosynthesis in storage roots of purple-fleshed sweet potato[J]. Acta Physiologiae Plantarum, 2014, 36(5): 1123-1132 doi: 10.1007/s11738-014-1487-y |
| [58] | TOSSI V, AMENTA M, LAMATTINA L, et al. Nitric oxide enhances plant ultraviolet-B protection up-regulating gene expression of the phenylpropanoid biosynthetic pathway[J]. Plant, Cell & Environment, 2011, 34(6): 909-921 http://www.ncbi.nlm.nih.gov/pubmed/21332509 |
| [59] | YIN R H, SKVORTSOVA M Y, LOUBéRY S, et al. COP1 is required for UV-B-induced nuclear accumulation of the UVR8 photoreceptor[J]. Proceedings of the National Academy of Sciences of the United States of America, 2016, 113(30): E4415-E4422 doi: 10.1073/pnas.1607074113 |
| [60] | SRIVASTAVA A K, SENAPATI D, SRIVASTAVA A, et al. Short hypocotyl in white light1 interacts with elongated hypocotyl5 (HY5) and constitutive photomorphogenic1 (COP1) and promotes COP1-mediated degradation of HY5 during arabidopsis seedling development[J]. Plant Physiology, 2015, 169(4): 2922-2934 https://www.researchgate.net/profile/Sreeramaiah_Gangappa |
