Theory, hypothesis and application advance in chamber-based technology and methods for flux measurement
Jie WEI1, Chang-Hua CHEN1, Jing-Yuan WANG1, Xue-Fa WEN,1,2,*1Key Laboratory of Ecosystem Network Observation and Modeling, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China 2College of Resources and Environment, University of Chinese Academy of Sciences, Beijing 100190, China
National Key R&D Program of China(2017YFC0503904) National Natural Science Foundation of China(41830860) National Natural Science Foundation of China(41671257)
Abstract The exchange flux of greenhouse gases, such as carbon (CO2, CH4), nitrogen (N2O) and water vapour (H2O), is the core of material cycle in the ecosystem and the bond of interaction among geosphere, biosphere and atmosphere. The development of stable isotope infrared spectroscopy and mass spectrometry technology and methods makes it possible to measure carbon stable isotopic composition (δ13C) and oxygen stable isotopic composition (δ18O)(CO2), δ13C (CH4), nitrogen stable isotope composition (δ15N) and δ18O (N2O), hydrogen stable isotopic composition (δD) and δ18O (H2O), which realizes the observation of greenhouse gas and its isotope flux at the soil, plant and ecosystem scales in combined with chamber-based technology and methods for flux measurement. Taking the chamber-based technology and methods for CO2 and its δ13C flux measurement as an example, this review which summarizes the basic principle and classification of the flux measurement system, expounds the theory requirements and assumptions of system design, summarizes the application advance and problems of chamber-based technology and methods for flux measurement in soil, plants (leaf, stem, and root) and ecosystem scales from the field to indoor, and prospects the importance of precision and accuracy of gas analysis and measurement data and the representativeness of measurement data in chamber-based flux measurement. Keywords:greenhouse gas;stable isotope;precision;accuracy;representative
PDF (1443KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 魏杰, 陈昌华, 王晶苑, 温学发. 箱式通量观测技术和方法的理论假设及其应用进展. 植物生态学报, 2020, 44(4): 318-329. DOI: 10.17521/cjpe.2019.0201 WEI Jie, CHEN Chang-Hua, WANG Jing-Yuan, WEN Xue-Fa. Theory, hypothesis and application advance in chamber-based technology and methods for flux measurement. Chinese Journal of Plant Ecology, 2020, 44(4): 318-329. DOI: 10.17521/cjpe.2019.0201
生态系统各种生源元素在土壤、植物、动物、微生物和大气间迁移和转化的物质循环过程研究是生态系统生态学的前沿性科学问题(于贵瑞, 2009; 于振良, 2017)。碳循环、氮循环和水循环是最重要的物质循环, 碳(CO2、CH4)、氮(N2O)和水汽(H2O)等温室气体的交换通量是生态系统物质循环的核心, 是地圈-生物圈-大气圈相互作用的纽带(于贵瑞和孙晓敏, 2017)。同位素技术具有指示、示踪和整合功能, 可以辅助解析生态系统碳氮水的生物地球化学循环过程与机制(温学发等, 2019)。稳定同位素红外光谱技术和方法的进步使碳稳定同位素比值(δ13C)和氧稳定同位素比值(δ18O)(CO2)、δ13C (CH4)、氮稳定同位素比值(δ15N)和δ18O (N2O)、氢稳定同位素比值(δD)和δ18O (H2O)等温室气体同位素组成的连续观测成为可能(Wen et al., 2008, 2013), 与箱式通量观测技术和方法结合可以实现温室气体及其同位素通量观测研究, 有助于揭示温室气体在土壤-植物-大气连续体的产生、传输和转化的相关过程与机制等(Wingate et al., 2010; Bowling et al., 2015; Epron et al., 2016)。
Fig. 1System characteristics and CO2 concentration change inside of chamber at non-steady-state (A, B) and steady-state (C, D). Cc and Ca represent the CO2 concentration inside and outside of the chamber, respectively; Pc and Pa represent the atmosphere pressure inside and outside of the chamber, respectively; Tc and Wc represent the air temperature and moisture inside the chamber, respectively.
Fig. 2Diagram on cooperative observation system of the CO2 carbon stable isotope composition (δ13C) flux of soil and plant (A) and observation system of the CO2δ13C flux of canopy and ecosystem (B). In A, we only take leaf and soil chambers as an example to show the flow diagram of gas system (stem and root chambers same as leaf chamber).
土壤CO2及其δ13C通量的原位长期连续观测, 有助于解析土壤呼吸随植物生理和环境等因素变化过程中的微小波动(Midwood et al., 2008; Bowling et al., 2015)。土壤呼吸包括自养呼吸和异养呼吸, 而δ13C已经被证明可以用于拆分这两种组分(Midwood et al., 2008; Albanito et al., 2012)。Midwood等(2008)利用土壤CO2稳态系统与稳定同位素质谱仪(Isotope Ratio Mass Spectrometry, IRMS)连用测定, 通过两元混合模型计算表明大麦(Hordeum vulgare)幼苗根中50%的碳可能来自于光合作用新合成的碳水化合物。Albanito等(2012)通过设计的参考室和双气室的稳态系统, 利用三元混合模型计算出土壤有机质、凋落物和根对土壤呼吸的相对贡献比例分别为37%、33%和30%。Liu等(2018)利用非稳态系统进行周期性升温和降温(5-30 ℃)培养试验, 结果表明土壤微生物呼吸对升温和降温具有明显的非对称响应, 升温阶段土壤微生物呼吸的增长速度更快。
AlbanitoF, McAllisterJL, CescattiA, SmithP, RobinsonD (2012). Dual-chamber measurements of δ13C of soilrespired CO2 partitioned using a field-based three end-member model Soil Biology & Biochemistry, 47, 106-115. DOI:10.1016/j.soilbio.2011.12.011URL [本文引用: 3]
BahnM, SchmittM, SiegwolfR, RichterA, BrüggemannN (2009). Does photosynthesis affect grassland soil-respired CO2 and its carbon isotope composition on a diurnal timescale? New Phytologist, 182, 451-460. DOI:10.1111/j.1469-8137.2008.02755.xURLPMID:19220762 [本文引用: 2] Soil respiration is the largest flux of carbon (C) from terrestrial ecosystems to the atmosphere. Here, we tested the hypothesis that photosynthesis affects the diurnal pattern of grassland soil-respired CO(2) and its C isotope composition (delta(13)C(SR)). A combined shading and pulse-labelling experiment was carried out in a mountain grassland. delta(13)C(SR) was monitored at a high time resolution with a tunable diode laser absorption spectrometer. In unlabelled plots a diurnal pattern of delta(13)C(SR) was observed, which was not explained by soil temperature, moisture or flux rates and contained a component that was also independent of assimilate supply. In labelled plots delta(13)C(SR) reflected a rapid transfer and respiratory use of freshly plant-assimilated C and a diurnal shift in the predominant respiratory C source from recent (i.e. at least 1 d old) to fresh (i.e. photoassimilates produced on the same day). We conclude that in grasslands the plant-derived substrates used for soil respiratory processes vary during the day, and that photosynthesis provides an important and immediate C source. These findings indicate a tight coupling in the plant-soil system and the importance of plant metabolism for soil CO(2) fluxes.
BarbourMM, EvansJR, SimoninKA, von CaemmererS (2016). Online CO2 and H2O oxygen isotope fractionation allows estimation of mesophyll conductance in C4 plants, and reveals that mesophyll conductance decreases as leaves age in both C4 and C3 plants New Phytologist, 210, 875-889. URLPMID:26778088 [本文引用: 1]
BarbourMM, McDowellNG, TcherkezG, BickfordCP, HansonDT (2007). A new measurement technique reveals rapid post-illumination changes in the carbon isotope composition of leaf-respired CO2 Plant, Cell & Environment, 30, 469-482. DOI:10.1111/j.1365-3040.2007.01634.xURLPMID:17324233 [本文引用: 1] We describe an open leaf gas exchange system coupled to a tunable diode laser (TDL) spectroscopy system enabling measurement of the leaf respiratory CO(2) flux and its associated carbon isotope composition (delta(13)C(Rl)) every 3 min. The precision of delta(13)C(Rl) measurement is comparable to that of traditional mass spectrometry techniques. delta(13)C(Rl) from castor bean (Ricinus communis L.) leaves tended to be positively related to the ratio of CO(2) produced to O(2) consumed [respiratory quotient (RQ)] after 24-48 h of prolonged darkness, in support of existing models. Further, the apparent fractionation between respiratory substrates and respired CO(2) within 1-8 h after the start of the dark period was similar to previous observations. In subsequent experiments, R. communis plants were grown under variable water availability to provide a range in delta(13)C of recently fixed carbohydrate. In leaves exposed to high light levels prior to the start of the dark period, CO(2) respired by leaves was up to 11 per thousand more enriched than phloem sap sugars within the first 10-15 min after plants had been moved from the light into the dark. The (13)C enrichment in respired CO(2) then decreased rapidly to within 3-7 per thousand of phloem sap after 30-60 min in the dark. This strong enrichment was not observed if light levels were low prior to the start of the dark period. Measurements of RQ confirmed that carbohydrates were the likely respiratory substrate for plants (RQ > 0.8) within the first 60 min after illumination. The strong (13)C enrichment that followed a high light-to-dark transition coincided with high respiration rates, suggesting that so-called light-enhanced dark respiration (LEDR) is fed by (13)C-enriched metabolites.
BowlingDR, EganJE, HallSJ, RiskDA (2015). Environmental forcing does not induce diel or synoptic variation in carbon isotope content of forest soil respiration Biogeosciences, 12, 5143-5160. [本文引用: 4]
BowlingDR, PatakiDE, RandersonJT (2008). Carbon isotopes in terrestrial ecosystem pools and CO2 fluxes New Phytologist, 178, 24-40. URLPMID:18179603 [本文引用: 1]
BuchmannN, EhleringerJR (1998). CO2 concentration profiles, and carbon and oxygen isotopes in C3 and C4 crop canopies Agricultural and Forest Meteorology, 89, 45-58. DOI:10.1016/S0168-1923(97)00059-2URL [本文引用: 1]
ChenCH, PangJP, WeiJ, WenXF, SunXM (2017). Intercomparison of three models for δ13C of respiration with four regression approaches Agricultural and Forest Meteorology, 247, 229-239. [本文引用: 3]
ChenCH, WeiJ, WenXF, SunXM, GuoQJ (2019). Photosynthetic carbon isotope discrimination and effects on daytime NEE partitioning in a subtropical mixed conifer plantation Agricultural and Forest Meteorology, 272-273, 143-155. [本文引用: 1]
DavidsonEA, SavageK, VerchotLV, NavarroR (2002). Minimizing artifacts and biases in chamber-based measurements of soil respiration Agricultural and Forest Meteorology, 113, 21-37. [本文引用: 2]
DoutheC, DreyerE, EpronD, WarrenCR (2011). Mesophyll conductance to CO2, assessed from online TDL-AS records of 13CO2 discrimination, displays small but significant short-term responses to CO2 and irradiance in Eucalyptus seedlings Journal of Experimental Botany, 62, 5335-5346. DOI:10.1093/jxb/err141URLPMID:21841176 [本文引用: 2] Mesophyll conductance (g(m)) is now recognized as an important limiting process for photosynthesis, as it results in a significant decrease of CO(2) diffusion from substomatal cavities where water evaporation occurs, to chloroplast stroma. Over the past decade, an increasing number of studies proposed that g(m) can vary in the short term (e.g. minutes), but these variations are still controversial, especially those potentially induced by changing CO(2) and irradiance. In this study, g(m) data estimated with online (13)C discrimination recorded with a tunable diode laser absorption spectrometer (TDL-AS) during leaf gas exchange measurements, and based on the single point method, are presented. The data were obtained with three Eucalyptus species. A 50% decrease in g(m) was observed when the CO(2) mole fraction was increased from 300 mumol mol(-1) to 900 mumol mol(-1), and a 60% increase when irradiance was increased from 200 mumol mol(-1) to 1100 mumol mol(-1) photosynthetic photon flux density (PPFD). The relative contribution of respiration and photorespiration to overall (13)C discrimination was also estimated. Not taking this contribution into account may lead to a 50% underestimation of g(m) but had little effect on the CO(2)- and irradiance-induced changes. In conclusion, (i) the observed responses of g(m) to CO(2) and irradiance were not artefactual; (ii) the respiratory term is important to assess absolute values of g(m) but has no impact on the responses to CO(2) and PPFD; and (iii) increasing irradiance and reducing the CO(2) mole fraction results in rapid increases in g(m) in Eucalyptus seedlings.
DubbertM, CuntzM, PiaydaA, WernerC (2014). Oxygen isotope signatures of transpired water vapor: the role of isotopic non-steady-state transpiration under natural conditions New Phytologist, 203, 1242-1252. URLPMID:24909361 [本文引用: 1]
EkbladA, H?gbergP (2001). Natural abundance of 13C in CO2 respired from forest soils reveals speed of link between tree photosynthesis and root respiration Oecologia, 127, 305-308. DOI:10.1007/s004420100667URLPMID:28547099 [本文引用: 1] Soil respiration from a boreal mixed coniferous forest showed large seasonal variation in natural abundance of (13)C, ranging from -21.6 per thousand to -26.5 per thousand. We tested if weather conditions could explain this variation in delta(13)C of respired CO2, and found that the air relative humidity 1-4 days before the days of CO2 sampling best explained the variation. This suggested that high delta(13)C values were caused by effects of air humidity on isotope fractionation during photosynthesis and that it took 1-4 days for the C from canopy photosynthesis of 20-25 m trees to become available for root/rhizosphere respiration. We calculated that these new photoassimilates could account for at least 65% of total soil respiration.
EpronD, CabralOMR, LaclauJP, DannouraM, PackerAP, PlainC, Battie-LaclauP, MoreiraMZ, TrivelinPCO, BouilletJP, GérantD, NouvellonY (2016). In situ13CO2 pulse labelling of field-grown eucalypt trees revealed the effects of potassium nutrition and throughfall exclusion on phloem transport of photosynthetic carbon Tree Physiology, 36, 6-21. DOI:10.1093/treephys/tpv090URLPMID:26423335 [本文引用: 6] Potassium (K) is an important limiting factor of tree growth, but little is known of the effects of K supply on the long-distance transport of photosynthetic carbon (C) in the phloem and of the interaction between K fertilization and drought. We pulse-labelled 2-year-old Eucalyptus grandis L. trees grown in a field trial combining K fertilization (+K and -K) and throughfall exclusion (+W and -W), and we estimated the velocity of C transfer by comparing time lags between the uptake of (13)CO2 and its recovery in trunk CO2 efflux recorded at different heights. We also analysed the dynamics of the labelled photosynthates recovered in the foliage and in the phloem sap (inner bark extract). The mean residence time of labelled C in the foliage was short (21-31 h). The time series of (13)C in excess in the foliage was affected by the level of fertilization, whereas the effect of throughfall exclusion was not significant. The velocity of C transfer in the trunk (0.20-0.82 m h(-1)) was twice as high in +K trees than in -K trees, with no significant effect of throughfall exclusion except for one +K -W tree labelled in the middle of the drought season that was exposed to a more pronounced water stress (midday leaf water potential of -2.2 MPa). Our results suggest that besides reductions in photosynthetic C supply and in C demand by sink organs, the lower velocity under K deficiency is due to a lower cross-sectional area of the sieve tubes, whereas an increase in phloem sap viscosity is more likely limiting phloem transport under drought. In all treatments, 10 times less (13)C was recovered in inner bark extracts at the bottom of the trunk when compared with the base of the crown, suggesting that a large part of the labelled assimilates has been exported out of the phloem and replaced by unlabelled C. This supports the 'leakage-retrieval mechanism' that may play a role in maintaining the pressure gradient between source and sink organs required to sustain high velocity of phloem transport in tall trees.
GaoJ, HanGL, HuangBX, ShiSJ, JiaCR, RenYF (2011). Integration and performance test of an automatic multi- channel long-term soil respiration measurement system Scientia Silvae Sinicae, 47(9), 153-157. [本文引用: 2]
GriffisTJ (2013). Tracing the flow of carbon dioxide and water vapor between the biosphere and atmosphere: a review of optical isotope techniques and their application Agricultural and Forest Meteorology, 174-175, 85-109. [本文引用: 3]
GriffisTJ, BakerJM, SargentSD, TannerBD, ZhangJ (2004). Measuring field-scale isotopic CO2 fluxes with tunable diode laser absorption spectroscopy and micrometeorological techniques Agricultural and Forest Meteorology, 124, 15-29. [本文引用: 2]
HafnerS, UnteregelsbacherS, SeeberE, LenaB, XuXL, LiXG, GuggenbergerG, MieheG, KuzyakovY (2012). Effect of grazing on carbon stocks and assimilate partitioning in a Tibetan montane pasture revealed by 13CO2 pulse labeling Global Change Biology, 18, 528-538. [本文引用: 1]
HansonPJ, GundersonCA (2009). Root carbon flux: measurements versus mechanisms New Phytologist, 184, 4-6. URLPMID:19740275 [本文引用: 1]
HeNP, WangRM, GaoY, DaiJZ, WenXF, YuGR (2013). Changes in the temperature sensitivity of SOM decomposition with grassland succession: implications for soil C sequestration Ecology and Evolution, 3, 5045-5054. URLPMID:24455135 [本文引用: 1]
HilmanB, AngertA (2016). Measuring the ratio of CO2 efflux to O2 influx in tree stem respiration Tree Physiology, 36, 1422-1431. URLPMID:27417515 [本文引用: 1]
HopkinsF, Gonzalez-MelerMA, FlowerCE, LynchDJ, CzimczikC, TangJW, SubkeJA (2013). Ecosystem-level controls on root-rhizosphere respiration New Phytologist, 199, 339-351. DOI:10.1111/nph.12271URLPMID:23943914 [本文引用: 1] Recent advances in the partitioning of autotrophic from heterotrophic respiration processes in soils in conjunction with new high temporal resolution soil respiration data sets offer insights into biotic and environmental controls of respiration. Besides temperature, many emerging controlling factors have not yet been incorporated into ecosystem-scale models. We synthesize recent research that has partitioned soil respiration into its process components to evaluate effects of nitrogen, temperature and photosynthesis on autotrophic flux from soils at the ecosystem level. Despite the widely used temperature dependence of root respiration, gross primary productivity (GPP) can explain most patterns of ecosystem root respiration (and to some extent heterotrophic respiration) at within-season time-scales. Specifically, heterotrophi crespiration is influenced by a seasonally variable supply of recent photosynthetic products in the rhizosphere. The contribution of stored root carbon (C) to root respiratory fluxes also varied seasonally, partially decoupling the proportion of photosynthetic C driving root respiration. In order to reflect recent insights, new hierarchical models, which incorporate root respiration as a primary function of GPP and which respond to environmental variables by modifying Callocation belowground, are needed for better prediction of future ecosystem C sequestration.
HutchinsonGL, LivingstonGP (2001). Vents and seals in non-steady-state chambers used for measuring gas exchange between soil and the atmosphere European Journal of Soil Science, 52, 675-682. [本文引用: 2]
ImerD, MerboldL, EugsterW, BuchmannN (2013). Temporal and spatial variations of soil CO2, CH4 and N2O fluxes at three differently managed grasslands Biogeosciences, 10, 5931-5945. [本文引用: 1]
JiaSX, McLaughlinNB, GuJC, LiXP, WangZQ (2013). Relationships between root respiration rate and root morphology, chemistry and anatomy in Larix gmelinii and Fraxinus mandshurica Tree Physiology, 33, 579-589. URLPMID:23824239 [本文引用: 1]
KammerA, TuzsonB, EmmeneggerL, KnohlA, MohnJ, HagedornF (2011). Application of a quantum cascade laser- based spectrometer in a closed chamber system for real- time δ13C and δ18O measurements of soil-respired CO2 Agricultural and Forest Meteorology, 151, 39-48. [本文引用: 3]
KeelingCD (1958). The concentration and isotopic abundances of atmospheric carbon dioxide in rural areas Geochimica et Cosmochimica Acta, 13, 322-334. [本文引用: 1]
KuptzD, FleischmannF, MatyssekR, GramsTEE (2011a). Seasonal patterns of carbon allocation to respiratory pools in 60-yr-old deciduous (Fagus sylvatica) and evergreen (Picea abies) trees assessed via whole-tree stable carbon isotope labeling New Phytologist, 191, 160-172. URLPMID:21395596 [本文引用: 4]
KuptzD, MatyssekR, GramsTEE (2011b). Seasonal dynamics in the stable carbon isotope composition δ13C from non-leafy branch, trunk and coarse root CO2 efflux of adult deciduous (Fagus sylvatica) and evergreen (Picea abies) trees Plant, Cell & Environment, 34, 363-373. URLPMID:21054435 [本文引用: 4]
LaiZR, LuS, ZhangYQ, WuB, QinSG, FengW, LiuJB, FaKY (2016). Diel patterns of fine root respiration in a dryland shrub, measured in situ over different phenological stages Journal of Forest Research, 21, 31-42. [本文引用: 1]
LaiZR, ZhangYQ, WuB, QinSG, FengW, LiuJB (2015). Impacts of morphological traits and temperature on fine root respiration during dormancy of Caragana korshinskii. Ecological Research, 30, 337-345. [本文引用: 1]
LiJJ, LiuL, ChenDM, XuFW, ChengJH, BaiYF (2019). Effects of collar size and buried depth on the measurement of soil respiration in a typical steppe Chinese Journal of Plant Ecology, 43, 152-164. [本文引用: 1]
LiuY, WenXF, ZhangYH, TianJ, GaoY, OstleNJ, NiuSL, ChenSP, SunXM, HeNP (2018). Widespread asymmetric response of soil heterotrophic respiration to warming and cooling Science of the Total Environment, 635, 423-431. URLPMID:29677668 [本文引用: 1]
LivingstonGP, HutchinsonGL, SpartalianK (2006). Trace gas emission in chambers: a non-steady-state diffusion model Soil Science Society of America Journal, 70, 1459-1469. [本文引用: 2]
Lundeg?rdhH (1927). Carbon dioxide evolution of soil and crop growth Soil Science, 23, 417-453. [本文引用: 1]
MaierM, Schack-KirchnerH (2014). Using the gradient method to determine soil gas flux: a review Agricultural and Forest Meteorology, 192-193, 78-95. [本文引用: 2]
MaierM, Schack-KirchnerH, HildebrandEE, SchindlerD (2011). Soil CO2 efflux vs. soil respiration: implications for flux models Agricultural and Forest Meteorology, 151, 1723-1730. DOI:10.1016/j.agrformet.2011.07.006URL [本文引用: 1] In the long term, all CO2 produced in the soil must be emitted by the surface and soil CO2 efflux (F-CO2) must correspond to soil respiration (R-soil). In the short term, however, the efflux can deviate from the instantaneous soil respiration, if the amount of CO2 stored in the soil pore-space (S-CO2) is changing. We measured F-CO2 continuously for one year using an automated chamber system. Simultaneously, vertical soil profiles of CO2 concentration, moisture, and temperature were measured in order to assess the changes in the amount of CO2 stored in the soil. R-soil was calculated as the sum of the rate of change of the CO2 storage over time and Fah. The experiment was split into a warm and a cold season. The dependency of soil respiration and soil efflux on soil temperature and on soil moisture was analyzed separately. Only the moisture-driven model of the warm season was significantly different for F-CO2 and R-soil. At our site, a moisture-driven soil-respiration model derived from CO2 efflux data would underestimate the importance of soil moisture. This effect can be attributed to a temporary storage of CO2 in the soil pore-space after rainfalls where up to 40% of the respired CO2 were stored. (C) 2011 Elsevier B.V.
MakitaN, HiranoY, DannouraM, KominamiY, MizoguchiT, IshiiH, KanazawaY (2009). Fine root morphological traits determine variation in root respiration of Quercus serrata. Tree Physiology, 29, 579-585. URLPMID:19203981 [本文引用: 3]
MakitaN, KosugiY, DannouraM, TakanashiS, NiiyamaK, KassimAR, NikAR (2012). Patterns of root respiration rates and morphological traits in 13 tree species in a tropical forest Tree Physiology, 32, 303-312. URLPMID:22367761
MakitaN, YakuR, OhashiM, FukudaK, IkenoH, HiranoY (2013). Effects of excising and washing treatments on the root respiration rates of Japanese cedar (Cryptomeria japonica) seedlings Journal of Forest Research, 18, 379-383. DOI:10.1007/s10310-012-0355-0URL [本文引用: 1] Tree root respiration is an important component of the carbon balance in forest ecosystems; however, it is not clear whether root preparation treatments (such as excising and washing) affect root respiration measurements. Here, we aimed to compare the respiration rates of roots subjected to different treatments (i.e., washing with water vs. brushing without water, and excising vs. not excising) for 17-month-old seedlings of Cryptomeria japonica. Immediately after sampling an entire root system, the root respiration rate was measured on a mass basis using a closed static chamber system equipped with an infrared gas analyzer. We found that the respiration rates for roots that were excised 10-20 times were significantly higher than those for roots that were not excised. There was no significant difference in the root respiration rates between washing and brushing treatments. Our results indicate that large numbers of excisions (> 10 times) could lead to bias in the measured changes in specific root respiration rates, and imply that differences between washing and brushing treatments do not affect the specific root respiration rate. We conclude that potential variation in recorded root respiration rates could be minimized by standardizing the root preparation technique, which should involve rapidly removing all loose soil and limiting the extent of root excision.
MarronN, PlainC, LongdozB, EpronD (2009). Seasonal and daily time course of the 13C composition in soil CO2 efflux recorded with a tunable diode laser spectrophotometer (TDLS) Plant and Soil, 318, 137-151. DOI:10.1007/s11104-008-9824-9URL [本文引用: 4] Temporal variations of carbon isotope composition of soil CO2 efflux (FS and δ13CFS) at different time scales should reflect both temporal variations of the climate conditions that affect canopy functioning and temporal changes in the relative contribution of autotrophic respiration to total FS. A tunable diode laser spectrophotometer (TDLS) was installed in the Hesse forest (northeast of France) early during the 2007 growing season to determine the seasonal and daily variability in δ13CFS. This method, based on the measurement of the absorption of an infrared laser emission at specific wave lengths of the 13CO2 and 12CO2, allows the continuous monitoring of the two isotopologues. The concentrations of the two isotopologues in FS were continuously monitored from June to November 2007 using chamber method and Keeling plots drawn from nocturnal accumulation of CO2 below the canopy. These TDLS measurements and isotope ratio mass spectrometer based Keeling plots gave very similar values of δ13CFS, showing the reliability of the TDLS system in this context. Results were analysed with regard to seasonal and daily changes in climatic and edaphic variables and compared with the δ13C of CO2 respired by roots, litter and soil incubated under controlled conditions. Pronounced daily as well as seasonal variations in δ13CFS were recorded (up to 1.5‰). The range of variation of δ13CFS was of the same order of magnitude at both diurnal and seasonal scales. δ13CFS observed in the field fluctuated between values of litter and of root respiration recorded during incubation, suggesting that temporal (and probably spatial) variations were associated with changes in the relative contribution of the two compartments during the day and during the season.
MarsdenC, NouvellonY, M?BouAT, Saint-AndreL, JourdanC, KinanaA, EpronD (2008). Two independent estimations of stand-level root respiration on clonal Eucalyptus stands in Congo: up scaling of direct measurements on roots versus the trenched-plot technique New Phytologist, 177, 676-687. DOI:10.1111/j.1469-8137.2007.02300.xURLPMID:18069963 [本文引用: 3] Root respiration at the level of a forest stand, an important component of ecosystem carbon balance, has been estimated in the past using various methods, most of them being indirect and relying on soil respiration measurements. On a 3-yr-old Eucalyptus stand in Congo-Brazzaville, a method involving the upscaling of direct measurements made on roots in situ, was compared with an independent approach using soil respiration measurements conducted on control and trenched plots (i.e. without living roots). The first estimation was based on the knowledge of root-diameter distribution and on a relationship between root diameter and specific respiration rates. The direct technique involving the upscaling of direct measurements on roots resulted in an estimation of 1.53 micromol m(-2) s(-1), c. 50% higher than the mean estimation obtained with the indirect technique (1.05 micromol m(-2) s(-1)). Monte-Carlo simulations showed that the results carried high uncertainty, but this uncertainty was no higher for the direct method than for the trenched-plot method. The reduction of the uncertainties on upscaled results requires more extensive knowledge of temperature sensitivity and more confidence and precision on the respiration rates and biomasses of fine roots.
MatterJM, KelemenPB (2009). Permanent storage of carbon dioxide in geological reservoirs by mineral carbonation Nature Geoscience, 2, 837-841. [本文引用: 1]
MencucciniM, H?ltt?T (2010). The significance of phloem transport for the speed with which canopy photosynthesis and belowground respiration are linked New Phytologist, 185, 189-203. URLPMID:19825019 [本文引用: 1]
MerboldL, SteinlinC, HagedornF (2013). Winter greenhouse gas emissions (CO2, CH4 and N2O) from a sub-alpine grassland Biogeosciences, 10, 3185-3203. DOI:10.5194/bg-10-3185-2013URL [本文引用: 1]
MidwoodAJ, GebbingT, WendlerR, SommerkornM, HuntJE, MillardP (2006). Collection and storage of CO2 for 13C analysis: an application to separate soil CO2 efflux into root- and soil-derived components Rapid Communications in Mass Spectrometry, 20, 3379-3384. URLPMID:17051607 [本文引用: 1]
MidwoodAJ, MillardP (2011). Challenges in measuring the δ13C of the soil surface CO2 efflux Rapid Communications in Mass Spectrometry, 25, 232-242. URLPMID:21181784 [本文引用: 10]
MidwoodAJ, ThorntonB, MillardP (2008). Measuring the 13C content of soil-respired CO2 using a novel open chamber system Rapid Communications in Mass Spectrometry, 22, 2073-2081. DOI:10.1002/rcm.3588URLPMID:18521825 [本文引用: 11] Carbon dioxide respired by soils comes from both autotrophic and heterotrophic respiration. 13C has proved useful in differentiating between these two sources, but requires the collection and analysis of CO2 efflux from the soil. We have developed a novel, open chamber system which allows for the accurate and precise quantification of the delta13C of soil-respired CO2. The chamber was tested using online analyses, by configuring a GasBench II and continuous flow isotope ratio mass spectrometer, to measure the delta13C of the chamber air every 120 s. CO2 of known delta13C value was passed through a column of sand and, using the chamber, the CO2 concentration stabilized rapidly, but 60 min was required before the delta13C value was stable and identical to the cylinder gas (-33.3 per thousand). Changing the chamber CO2 concentration between 200 and 900 micromol.mol(-1) did not affect the measured delta13C of the efflux. Measuring the delta13C of the CO2 efflux from soil cores in the laboratory gave a spread of +/-2 per thousand, attributed to heterogeneity in the soil organic matter and roots. Lateral air movement through dry sand led to a change in the delta13C of the surface efflux of up to 8 per thousand. The chamber was used to measure small transient changes (+/-2 per thousand) in the delta13C of soil-respired CO2 from a peaty podzol after gradual heating from 12 to 35 degrees C over 12 h. Finally, soil-respired CO2 was partitioned in a labelling study and the contribution of autotrophic and heterotrophic respiration to the total efflux determined. Potential applications for the chamber in the study of soil respiration are discussed.
MillardP, MidwoodAJ, HuntJE, WhiteheadD, BouttonTW (2008). Partitioning soil surface CO2 efflux into autotrophic and heterotrophic components, using natural gradients in soil δ13C in an undisturbed savannah soil Soil Biology & Biochemistry, 40, 1575-1582. [本文引用: 2]
MillerJB, TansPP (2003). Calculating isotopic fractionation from atmospheric measurements at various scales Tellus B: Chemical and Physical Meteorology, 55, 207-214. [本文引用: 1]
Moreno-GutiérrezC, DawsonTE, NicolásE, QuerejetaJI (2012). Isotopes reveal contrasting water use strategies among coexisting plant species in a Mediterranean ecosystem New Phytologist, 196, 489-496. URLPMID:22913668 [本文引用: 2]
NickersonN, RiskD (2009). A numerical evaluation of chamber methodologies used in measuring the δ13C of soil respiration Rapid Communications in Mass Spectrometry, 23, 2802-2810. DOI:10.1002/rcm.4189URLPMID:19653202 [本文引用: 1] Measurement of the delta(13)C value of soil-respired CO(2) (delta(r)) has become a commonplace method through which ecosystem function and C dynamics can be better understood. Despite its proven utility there is currently no consensus on the most robust method with which to measure delta(r). Static and dynamic chamber systems are both commonly used for this purpose; however, the literature on these methods provides evidence suggesting that measurements of delta(r) made with these chamber systems are neither repeatable (self-consistent) nor comparable across methodologies. Here we use a three-dimensional (3-D) numerical soil-atmosphere-chamber model to test these chamber systems in a 'surrogate reality'. Our simulations show that each chamber methodology is inherently biased and that no chamber methodology can accurately predict the true delta(r) signature under field conditions. If researchers intend to use delta(r) to study in situ ecosystem processes, the issues with these chamber systems need to be corrected either by using diffusive theory or by designing a new, unbiased delta(r) measurement system.
OhlssonKEA, SinghB, HolmS, NordgrenA, L?vdahlL, H?gbergP (2005). Uncertainties in static closed chamber measurements of the carbon isotopic ratio of soil-respired CO2 Soil Biology & Biochemistry, 37, 2273-2276. [本文引用: 2]
PangJP, WenXF (2018). A review of the calibration methods for measuring the carbon and oxygen isotopes in CO2 based on isotope ratio infrared spectroscopy Chinese Journal of Plant Ecology, 42, 143-152. [本文引用: 1]
PangJP, WenXF, SunXM (2016a). Mixing ratio and carbon isotopic composition investigation of atmospheric CO2 in Beijing, China Science of the Total Environment, 539, 322-330. DOI:10.1016/j.scitotenv.2015.08.130URLPMID:26363727 [本文引用: 1] The stable isotope composition of atmospheric CO2 can be used as a tracer in the study of urban carbon cycles, which are affected by anthropogenic and biogenic CO2 components. Continuous measurements of the mixing ratio and delta(13)C of atmospheric CO2 were conducted in Beijing from Nov. 15, 2012 to Mar. 8, 2014 including two heating seasons and a vegetative season. Both delta(13)C and the isotopic composition of source CO2 (delta(13)CS) were depleted in the heating seasons and enriched in the vegetative season. The diurnal variations in the CO2 mixing ratio and delta(13)C contained two peaks in the heating season, which are due to the effects of morning rush hour traffic. Seasonal and diurnal patterns of the CO2 mixing ratio and delta(13)C were affected by anthropogenic emissions and biogenic activity. Assuming that the primary CO2 sources at night (22:00-04:00) were coal and natural gas combustion during heating seasons I and II, an isotopic mass balance analysis indicated that coal combustion had average contributions of 83.83+/-14.11% and 86.84+/-12.27% and that natural gas had average contributions of 16.17+/-14.11% and 13.16+/-12.27%, respectively. The delta(13)C of background CO2 in air was the main error source in the isotopic mass balance model. Both the mixing ratio and delta(13)C of atmospheric CO2 had significant linear relationships with the air quality index (AQI) and can be used to indicate local air pollution conditions. Energy structure optimization, for example, reducing coal consumption, will improve the local air conditions in Beijing.
PangJP, WenXF, SunXM, HuangK (2016b). Intercomparison of two cavity ring-down spectroscopy analyzers for atmospheric 13CO2/12CO2 measurement Atmospheric Measurement Techniques, 9, 3879-3891. [本文引用: 1]
ParkinTB, KasparTC, SenwoZ, PruegerJH, HatfieldJL (2005). Relationship of soil respiration to crop and landscape in the walnut creek watershed Journal of Hydrometeorology, 6, 812-824. [本文引用: 1]
PlainC, GerantD, MaillardP, DannouraM, DongY, ZellerB, PriaultP, ParentF, EpronD (2009). Tracing of recently assimilated carbon in respiration at high temporal resolution in the field with a tuneable diode laser absorption spectrometer after in situ13CO2 pulse labelling of 20-year-old beech trees Tree Physiology, 29, 1433-1445. DOI:10.1093/treephys/tpp072URLPMID:19797042 [本文引用: 5] The study of the fate of assimilated carbon in respiratory fluxes in the field is needed to resolve the residence and transfer times of carbon in the atmosphere-plant-soil system in forest ecosystems, but it requires high frequency measurements of the isotopic composition of evolved CO2. We developed a closed transparent chamber to label the whole crown of a tree and a labelling system capable of delivering a 3-h pulse of 99% 13CO2 in the field. The isotopic compositions of trunk and soil CO2 effluxes were recorded continuously on two labelled and one control trees by a tuneable diode laser absorption spectrometer during a 2-month chase period following the late summer labelling. The lag times for trunk CO2 effluxes are consistent with a phloem sap velocity of about 1 m h(-1). The isotopic composition (delta13C) of CO2 efflux from the trunk was maximal 2-3 days after labelling and declined thereafter following two exponential decays with a half-life of 2-8 days for the first and a half-life of 15-16 days for the second. The isotopic composition of the soil CO2 efflux was maximal 3-4 days after labelling and the decline was also well fitted with a sum of two exponential functions with a half-life of 3-5 days for the first exponential and a half-life of 16-18 days for the second. The amount of label recovered in CO2 efflux was around 10-15% of the assimilated 13CO2 for soil and 5-13% for trunks. As labelling occurred late in the growing season, substantial allocation to storage is expected.
PowersHH, HuntJE, HansonDT, McDowellNG (2010). A dynamic soil chamber system coupled with a tunable diode laser for online measurements of δ13C, δ18O, and efflux rate of soil-respired CO2 Rapid Communications in Mass Spectrometry, 24, 243-253. URLPMID:20049893 [本文引用: 5]
PumpanenJ, KolariP, IlvesniemiH, MinkkinenK, VesalaT, Niinist?S, LohilaA, LarmolaT, MoreroM, PihlatieM, JanssensI, YusteJC, GrünzweigJM, RethS, SubkeJA, SavageK, KutschW, ?strengG, ZieglerW, AnthoniP, LindrothA, HariP (2004). Comparison of different chamber techniques for measuring soil CO2 efflux Agricultural and Forest Meteorology, 123, 159-176. [本文引用: 1]
ReyA (2015). Mind the gap: non-biological processes contributing to soil CO2 efflux Global Change Biology, 21, 1752-1761. DOI:10.1111/gcb.12821URLPMID:25471988 [本文引用: 2] Widespread recognition of the importance of soil CO2 efflux as a major source of CO2 to the atmosphere has led to active research. A large soil respiration database and recent reviews have compiled data, methods, and current challenges. This study highlights some deficiencies for a proper understanding of soil CO2 efflux focusing on processes of soil CO2 production and transport that have not received enough attention in the current soil respiration literature. It has mostly been assumed that soil CO2 efflux is the result of biological processes (i.e. soil respiration), but recent studies demonstrate that pedochemical and geological processes, such as geothermal and volcanic CO2 degassing, are potentially important in some areas. Besides the microbial decomposition of litter, solar radiation is responsible for photodegradation or photochemical degradation of litter. Diffusion is considered to be the main mechanism of CO2 transport in the soil, but changes in atmospheric pressure and thermal convection may also be important mechanisms driving soil CO2 efflux greater than diffusion under certain conditions. Lateral fluxes of carbon as dissolved organic and inorganic carbon occur and may cause an underestimation of soil CO2 efflux. Traditionally soil CO2 efflux has been measured with accumulation chambers assuming that the main transport mechanism is diffusion. New techniques are available such as improved automated chambers, CO2 concentration profiles and isotopic techniques that may help to elucidate the sources of carbon from soils. We need to develop specific and standardized methods for different CO2 sources to quantify this flux on a global scale. Biogeochemical models should include biological and non-biological CO2 production processes before we can predict the response of soil CO2 efflux to climate change. Improving our understanding of the processes involved in soil CO2 efflux should be a research priority given the importance of this flux in the global carbon budget.
SalomónRL, Valbuena-Caraba?aM, GilL, McGuireMA, TeskeyRO, AubreyDP, González-DoncelI, Rodríguez- CalcerradaJ (2016). Temporal and spatial patterns of internal and external stem CO2 fluxes in a sub-Mediterranean oak Tree Physiology, 36, 1409-1421. DOI:10.1093/treephys/tpw029URLPMID:27126229 [本文引用: 1] To accurately estimate stem respiration (RS), measurements of both carbon dioxide (CO2) efflux to the atmosphere (EA) and internal CO2 flux through xylem (FT) are needed because xylem sap transports respired CO2 upward. However, reports of seasonal dynamics of FT and EA are scarce and no studies exist in Mediterranean species under drought stress conditions. Internal and external CO2 fluxes at three stem heights, together with radial stem growth, temperature, sap flow and shoot water potential, were measured in Quercus pyrenaica Willd. in four measurement campaigns during one growing season. Substantial daytime depressions in temperature-normalized EA were observed throughout the experiment, including prior to budburst, indicating that diel hysteresis between stem temperature and EA cannot be uniquely ascribed to diversion of CO2 in the transpiration stream. Low internal [CO2] (<0.5%) resulted in low contributions of FT to RS throughout the growing season, and RS was mainly explained by EA (>90%). Internal [CO2] was found to vary vertically along the stems. Seasonality in resistance to radial CO2 diffusion was related to shoot water potential. The low internal [CO2] and FT observed in our study may result from the downregulation of xylem respiration in response to a legacy of coppicing as well as high radial diffusion of CO2 through cambium, phloem and bark tissues, which was related to low water content of stems. Long-term studies analyzing temporal and spatial variation in internal and external CO2 fluxes and their interactions are needed to mechanistically understand and model respiration of woody tissues.
SavageK, PhillipsR, DavidsonE (2014). High temporal frequency measurements of greenhouse gas emissions from soils Biogeosciences, 11, 2709-2720. [本文引用: 3]
ShiXL, WangCK, XuF, WangXC (2010). Temporal dynamics and influencing factors of stem respiration for four temperate tree species Acta Ecologica Sinica, 30, 3994-4003. [本文引用: 1]
SnellHSK, RobinsonD, MidwoodAJ (2014). Minimising methodological biases to improve the accuracy of partitioning soil respiration using natural abundance 13C Rapid Communications in Mass Spectrometry, 28, 2341-2351. URLPMID:25279748 [本文引用: 1]
SturmP, TuzsonB, HenneS, EmmeneggerL (2013). Tracking isotopic signatures of CO2 at the high altitude site Jungfraujoch with laser spectroscopy: analytical improvements and representative results Atmospheric Measurement Techniques, 6, 1659-1671. DOI:10.5194/amt-6-1659-2013URL [本文引用: 1] We present the continuous data record of atmospheric CO2 isotopes measured by laser absorption spectroscopy for an almost four year period at the High Altitude Research Station Jungfraujoch (3580 ma.s.l.), Switzerland. The mean annual cycles derived from data of December 2008 to September 2012 exhibit peak-to-peak amplitudes of 11.0 mu mol mol(-1) for CO2, 0.60 parts per thousand for delta C-13 and 0.81 parts per thousand for delta O-18. The high temporal resolution of the measurements also allow us to capture variations on hourly and diurnal timescales. For CO2 the mean diurnal peak-to-peak amplitude is about 1 mu mol mol(-1) in spring, autumn and winter and about 2 mu mol mol(-1) in summer. The mean diurnal variability in the isotope ratios is largest during the summer months too, with an amplitude of about 0.1 parts per thousand both in the delta C-13 and delta O-18, and a smaller or no discernible diurnal cycle during the other seasons. The day-to-day variability, however, is much larger and depends on the origin of the air masses arriving at Jungfraujoch. Backward Lagrangian particle dispersion model simulations revealed a close link between air composition and prevailing transport regimes and could be used to explain part of the observed variability in terms of transport history and influence region. A footprint clustering showed significantly different wintertime CO2, delta C-13 and delta O-18 values depending on the origin and surface residence times of the air masses. Several major updates on the instrument and the calibration procedures were performed in order to further improve the data quality. We describe the new measurement and calibration setup in detail and demonstrate the enhanced performance of the analyzer. A measurement precision of about 0.02 parts per thousand for both isotope ratios has been obtained for an averaging time of 10 min, while the accuracy was estimated to be 0.1 parts per thousand, including the uncertainty of the calibration gases.
SubkeJA, VallackHW, MagnussonT, KeelSG, MetcalfeDB, H?gbergP, InesonP (2009). Short-term dynamics of abiotic and biotic soil 13CO2 effluxes after in situ13CO2 pulse labelling of a boreal pine forest New Phytologist, 183, 349-357. DOI:10.1111/j.1469-8137.2009.02883.xURLPMID:19496953 [本文引用: 2] Physical diffusion of isotopic tracers into and out of soil pores causes considerable uncertainty for the timing and magnitude of plant belowground allocation in pulse-labelling experiments. Here, we partitioned soil CO(2) isotopic fluxes into abiotic tracer flux (physical return), heterotrophic flux, and autotrophic flux contributions following (13)CO(2) labelling of a Swedish Pinus sylvestris forest. Soil CO(2) efflux and its isotopic composition from a combination of deep and surface soil collars was monitored using a field-deployed mass spectrometer. Additionally, (13)CO(2) within the soil profile was monitored. Physical (abiotic) efflux of (13)CO(2) from soil pore spaces was found to be significant for up to 48 h after pulse labelling, and equalled the amount of biotic label flux over 6 d. Measured and modelled changes in (13)CO(2) concentration throughout the soil profile corroborated these results. Tracer return via soil CO(2) efflux correlated significantly with the proximity of collars to trees, while daily amplitudes of total flux (including heterotrophic and autotrophic sources) showed surprising time shifts compared with heterotrophic fluxes. The results show for the first time the significance of the confounding influence of physical isotopic CO(2)-tracer return from the soil matrix, calling for the inclusion of meaningful control treatments in future pulse-chase experiments.
SunLJ, AtakaM, KominamiY, YoshimuraK (2017). Relationship between fine-root exudation and respiration of two Quercus species in a Japanese temperate forest Tree Physiology, 37, 1011-1020. URLPMID:28338964 [本文引用: 2]
SunT, MaoZJ (2011). Functional relationships between morphology and respiration of fine roots in two Chinese temperate tree species Plant and Soil, 346, 375-384. DOI:10.1007/s11104-011-0825-8URL [本文引用: 1] Fine root morphology is closely linked with physiological functions. However, we have limited understanding of relationships between morphological traits and respiration of fine roots. In this study, we measured the fine root respiration rates of Tilia amurensis Rupr. and Pinus koraiensis Sieb. et Zucc with a simplified specially designed root-chamber system in the field. We found that mean diameter and specific root length (SRL) exhibited significant power relationships with fine root respiration in both species. Respiration rates of smaller fine roots (< 0.5 mm for Tilia amurensis and < 0.7 mm for Pinus koraiensis) in mean diameter were much higher and varied widely compared with those larger diameter class (0.6-1.8 mm for Tilia amurensis and 0.8-1.9 mm for Pinus koraiensis). Moreover, significant positive relationships existed between fine root nitrogen (N) concentration and respiration in both species. For Tilia amurensis, SRL explained more of the variability in respiration (84%) than diameter or N concentration, while for Pinus koraiensis, fine root N concentration explained more of the variability in respiration (79%) than diameter or SRL. Our results suggest that fine roots are composed of individual roots differing markedly in respiration and morphology, rather than identical units as traditionally assumed.
SusfalkRB, ChengWX, JohnsonDW, WalkerRF, VerburgP, FuS (2002). Lateral diffusion and atmospheric CO2 mixing compromise estimates of rhizosphere respiration in a forest soil Canadian Journal of Forest Research, 32, 1005-1015. [本文引用: 2]
WangJY, WangXJ, WangJP (2018a). Profile distribution of CO2 in an arid saline-alkali soil with gypsum and wheat straw amendments: a two-year incubation experiment Scientific Reports, 8, 11939. DOI: 10.038/s41598-018-30312-0. URLPMID:30093635 [本文引用: 1]
WangXW, MaoZJ, McGuireMA, TeskeyRO (2019). Stem radial CO2 conductance affects stem respiratory CO2 fluxes in ash and birch trees Journal of Forestry Research, 30, 21-29. [本文引用: 1]
WangYY, LiXX, DongWX, WuDM, HuCS, ZhangYM, LuoYQ (2018b). Depth-dependent greenhouse gas production and consumption in an upland cropping system in northern China Geoderma, 319, 100-112. [本文引用: 1]
WeiJ, LiuWG, WanH, ChengJM, LiWJ (2016). Differential allocation of carbon in fenced and clipped grasslands: a 13C tracer study in the semiarid Chinese Loess Plateau Plant and Soil, 406, 251-263. [本文引用: 1]
WeissI, MizrahiY, RavehE (2009). Chamber response time: a neglected issue in gas exchange measurements Photosynthetica, 47, 121-124. [本文引用: 1]
WenXF, MengY, ZhangXY, SunXM, LeeX (2013). Evaluating calibration strategies for isotope ratio infrared spectroscopy for atmospheric 13CO2/12CO2 measurement Atmospheric Measurement Techniques, 6, 1491-1501. [本文引用: 3]
WenXF, SunXM, LiuYF, LiXB (2007). Effects of linear and exponential fitting on the initial rate of change in CO2 concentration across the soil surface Journal of Plant Ecology (Chinese Version), 31, 380-385. [本文引用: 1]
WenXF, SunXM, ZhangSC, YuGR, SargentSD, LeeXH (2008). Continuous measurement of water vapor D/H and 18O/16O isotope ratios in the atmosphere Journal of Hydrology, 349, 489-500. [本文引用: 1]
WenXF, ZhangXY, WeiJ, LüSD, WangJ, ChenCH, SongXW, WangJY, DaiXQ (2019). Understanding the biogeochemical process and mechanism of ecosystem carbon cycle from the perspective of the earth’s critical zone Advances in Earth Science, 34, 471-479. [本文引用: 1]
WingateL, OgéeJ, BurlettR, BoscA, DevauxM, GraceJ, LoustauD, GesslerA (2010). Photosynthetic carbon isotope discrimination and its relationship to the carbon isotope signals of stem, soil and ecosystem respiration New Phytologist, 188, 576-589. DOI:10.1111/j.1469-8137.2010.03384.xURLPMID:20663061 [本文引用: 2] * Photosynthetic carbon (C) isotope discrimination (Delta(Alpha)) labels photosynthates (delta(A) ) and atmospheric CO(2) (delta(a)) with variable C isotope compositions during fluctuating environmental conditions. In this context, the C isotope composition of respired CO(2) within ecosystems is often hypothesized to vary temporally with Delta(Alpha). * We investigated the relationship between Delta(Alpha) and the C isotope signals from stem (delta(W)), soil (delta(S)) and ecosystem (delta(E)) respired CO(2) to environmental fluctuations, using novel tuneable diode laser absorption spectrometer instrumentation in a mature maritime pine forest. * Broad seasonal changes in Delta(Alpha) were reflected in delta(W,) delta(S) and delta(E). However, respired CO(2) signals had smaller short-term variations than Delta(A) and were offset and delayed by 2-10 d, indicating fractionation and isotopic mixing in a large C pool. Variations in delta(S) did not follow Delta(A) at all times, especially during rainy periods and when there is a strong demand for C allocation above ground. * It is likely that future isotope-enabled vegetation models will need to develop transfer functions that can account for these phenomena in order to interpret and predict the isotopic impact of biosphere gas exchange on the C isotope composition of atmospheric CO(2).
WuYB, TanHC, DengYC, WuJ, XuXL, WangYF, TangYH, TeruoH, CuiXY (2010). Partitioning pattern of carbon flux in a Kobresia grassland on the Qinghai-Tibetan Plateau revealed by field13C pulse-labeling Global Change Biology, 16, 2322-2333. [本文引用: 1]
XuLK, FurtawMD, MadsenRA, GarciaRL, AndersonDJ, McDermittDK (2006). On maintaining pressure equilibrium between a soil CO2 flux chamber and the ambient air Journal of Geophysical Research, 111, D08S10. DOI: 10.1029/2005JD006435. [本文引用: 1]
YangJY, HeYJ, AubreyDP, ZhuangQL, TeskeyRO (2016). Global patterns and predictors of stem CO2 efflux in forest ecosystems Global Change Biology, 22, 1433-1444. DOI:10.1111/gcb.13188URLPMID:26667780 [本文引用: 2] Stem CO2 efflux (ES) plays an important role in the carbon balance of forest ecosystems. However, its primary controls at the global scale are poorly understood and observation-based global estimates are lacking. We synthesized data from 121 published studies across global forest ecosystems and examined the relationships between annual ES and biotic and abiotic factors at individual, biome, and global scales, and developed a global gridded estimate of annual ES . We tested the following hypotheses: (1) Leaf area index (LAI) will be highly correlated with annual ES at biome and global scales; (2) there will be parallel patterns in stem and root CO2 effluxes (RA) in all forests; (3) annual ES will decline with forest age; and (4) LAI coupled with mean annual temperature (MAT) and mean annual precipitation (MAP) will be sufficient to predict annual ES across forests in different regions. Positive linear relationships were found between ES and LAI, as well as gross primary production (GPP), net primary production (NPP), wood NPP, soil CO2 efflux (RS), and RA . Annual ES was correlated with RA in temperate forests after controlling for GPP and MAT, suggesting other additional factors contributed to the relationship. Annual ES tended to decrease with stand age. Leaf area index, MAT and MAP, predicted 74% of variation in ES at global scales. Our statistical model estimated a global annual ES of 6.7 +/- 1.1 Pg C yr(-1) over the period of 2000-2012 with little interannual variability. Modeled mean annual ES was 71 +/- 43, 270 +/- 103, and 420 +/- 134 g C m(2) yr(-1) for boreal, temperate, and tropical forests, respectively. We recommend that future studies report ES at a standardized constant temperature, incorporate more manipulative treatments, such as fertilization and drought, and whenever possible, simultaneously measure both aboveground and belowground CO2 fluxes.
YangQP, XuM, ChiYG, ZhengYP, ShenRC, LiPX, DaiHT (2012). Temporal and spatial variations of stem CO2 efflux of three species in subtropical China Journal of Plant Ecology, 5, 229-237. [本文引用: 1]
YangQP, ZhangWD, LiRS, ZhengWH, YangJY, XuM, GuanX, HuangK, ChenLC, WangQK, WangSL (2019). Effects of girdling on stem CO2 efflux and its temperature sensitivity in Chinese fir and sweetgum trees Agricultural and Forest Meteorology, 268, 116-123. DOI:10.1016/j.agrformet.2019.01.021URL [本文引用: 2]
YuGR (2009). Scientific Frontier on Human Activities and Ecosystem Changes. Higher Education Press, Beijing. [本文引用: 1]
ZhaoJY, XiaoW, ZhangM, WangJY, WenXF, LeeXH (2020). Applications and prospect of the flux-gradient method in measuring the greenhouse gases and isotope fluxes Chinese Journal of Plant Ecology, 44, 305-317. [本文引用: 1]
ZhaoKJ, DongBQ, JiaZK, MaLY (2018). Effect of climatic factors on the temporal variation of stem respiration in Larix principis-rupprechtii Mayr Agricultural and Forest Meteorology, 248, 441-448. [本文引用: 1]
ZhouJ, YangZY, WuGH, YangYZ, LinGH (2018). The relationship between soil CO2 efflux and its carbon isotopic composition under non-steady-state conditions Agricultural and Forest Meteorology, 256-257, 492-500. [本文引用: 1]
Dual-chamber measurements of δ13C of soilrespired CO2 partitioned using a field-based three end-member model 3 2012
... 稳态系统满足上述假设的方法通常也包括两种.首先, 通过增加CO2浓度调节装置向箱体内注入无CO2的空气, 保持箱体进气口和出气口气体流速相同, 使箱体内总CO2浓度始终与箱体外总CO2浓度保持动态平衡(Midwood et al., 2008; Marron et al., 2009; Powers et al., 2010).其次, 通过参考室与双层气室系统设计达到箱体内总CO2浓度始终与箱体外总CO2浓度保持动态平衡(Albanito et al., 2012), 其中参考气室上表面为CO2自由渗透膜, 双层气室的上层气室相当于上述第一种途径的稳态气室, 中间利用CO2渗透膜隔开, 下层气室为非稳态气室; 参考气室和上层气室利用红外光谱技术测定, 当二者CO2浓度一致时, 利用稳定同位素红外光谱技术测定下层气室CO2 δ13C值. ...
... 土壤CO2及其δ13C通量的原位长期连续观测, 有助于解析土壤呼吸随植物生理和环境等因素变化过程中的微小波动(Midwood et al., 2008; Bowling et al., 2015).土壤呼吸包括自养呼吸和异养呼吸, 而δ13C已经被证明可以用于拆分这两种组分(Midwood et al., 2008; Albanito et al., 2012).Midwood等(2008)利用土壤CO2稳态系统与稳定同位素质谱仪(Isotope Ratio Mass Spectrometry, IRMS)连用测定, 通过两元混合模型计算表明大麦(Hordeum vulgare)幼苗根中50%的碳可能来自于光合作用新合成的碳水化合物.Albanito等(2012)通过设计的参考室和双气室的稳态系统, 利用三元混合模型计算出土壤有机质、凋落物和根对土壤呼吸的相对贡献比例分别为37%、33%和30%.Liu等(2018)利用非稳态系统进行周期性升温和降温(5-30 ℃)培养试验, 结果表明土壤微生物呼吸对升温和降温具有明显的非对称响应, 升温阶段土壤微生物呼吸的增长速度更快. ...
Does photosynthesis affect grassland soil-respired CO2 and its carbon isotope composition on a diurnal timescale? 2 2009
... 稳态箱式通量观测系统是通过使箱体内气体形成开放气路, 保持箱体内外CO2浓度一致或仅略高于大气, 通过测定箱体进气口和出气口CO2浓度差计算通量, 如图1C和1D所示.根据是否增加CO2浓度调节装置, 通常可以分为两类: (1)不增加CO2浓度调节装置, 仅通过将气泵和电磁阀切换实现利用气体分析仪测定进气口(参考气)和出气口(样品气) CO2浓度并计算差值.在测定过程中, 箱体内总CO2浓度略高于大气, 同时12CO2和13CO2浓度均略高于大气(Bahn et al., 2009; Plain et al., 2009; Subke et al., 2009).(2)增加CO2浓度调节装置, 通过向箱体内注入无CO2的空气, 保持箱体进气口和出气口气体流速相同, 使箱体内总CO2浓度始终与箱体外总CO2浓度基本保持一致, 仅略高于大气, 但设计和操作较难(Midwood et al., 2006; Marron et al., 2009).稳态系统通常没有明显的CO2累积(一般低于100 μmol·mol-1)(Midwood & Milllard, 2011), 因此, 测定过程中通常不会对CO2扩散产生明显影响, 这使得稳态系统在测定δ13C时比非稳态系统更有优势(Bahn et al., 2009; Plain et al., 2009; Subke et al., 2009). ...
... C时比非稳态系统更有优势(Bahn et al., 2009; Plain et al., 2009; Subke et al., 2009). ...
Online CO2 and H2O oxygen isotope fractionation allows estimation of mesophyll conductance in C4 plants, and reveals that mesophyll conductance decreases as leaves age in both C4 and C3 plants 1 2016
... 相对于非稳态箱式通量观测系统, 稳态箱式通量观测系统应用更加广泛, 可以采用明箱或暗箱测定.商业化叶片CO2箱式通量观测系统包括LI-6400/6800等, 为稳态系统.通过红外光谱技术对气室内样品气和参考气进行差分测量, 计算CO2通量(Barbour et al., 2007; Douthe et al., 2011).叶片与大气之间的CO2平衡和水汽交换受到气孔导度的控制.随着研究的深入, 发现叶片内部传输过程对碳水交换的影响也非常重要(Moreno-Gutiérrez et al., 2012).叶片CO2 δ13C通量连续观测系统可通过从LI-6400/6800气室进气口和出气口分别抽取部分气体进入稳定同位素红外光谱仪, 即可同步测定叶片CO2及其δ13C通量(Douthe et al., 2011; Barbour et al., 2016), 这为理解CO2在叶片内部传输的动态过程提供了技术支撑, 可揭示叶肉导度对CO2导度和水汽导度之间存在的权衡关系(Moreno-Gutiérrez et al., 2012). ...
A new measurement technique reveals rapid post-illumination changes in the carbon isotope composition of leaf-respired CO2 1 2007
... 相对于非稳态箱式通量观测系统, 稳态箱式通量观测系统应用更加广泛, 可以采用明箱或暗箱测定.商业化叶片CO2箱式通量观测系统包括LI-6400/6800等, 为稳态系统.通过红外光谱技术对气室内样品气和参考气进行差分测量, 计算CO2通量(Barbour et al., 2007; Douthe et al., 2011).叶片与大气之间的CO2平衡和水汽交换受到气孔导度的控制.随着研究的深入, 发现叶片内部传输过程对碳水交换的影响也非常重要(Moreno-Gutiérrez et al., 2012).叶片CO2 δ13C通量连续观测系统可通过从LI-6400/6800气室进气口和出气口分别抽取部分气体进入稳定同位素红外光谱仪, 即可同步测定叶片CO2及其δ13C通量(Douthe et al., 2011; Barbour et al., 2016), 这为理解CO2在叶片内部传输的动态过程提供了技术支撑, 可揭示叶肉导度对CO2导度和水汽导度之间存在的权衡关系(Moreno-Gutiérrez et al., 2012). ...
Environmental forcing does not induce diel or synoptic variation in carbon isotope content of forest soil respiration 4 2015
... 生态系统各种生源元素在土壤、植物、动物、微生物和大气间迁移和转化的物质循环过程研究是生态系统生态学的前沿性科学问题(于贵瑞, 2009; 于振良, 2017).碳循环、氮循环和水循环是最重要的物质循环, 碳(CO2、CH4)、氮(N2O)和水汽(H2O)等温室气体的交换通量是生态系统物质循环的核心, 是地圈-生物圈-大气圈相互作用的纽带(于贵瑞和孙晓敏, 2017).同位素技术具有指示、示踪和整合功能, 可以辅助解析生态系统碳氮水的生物地球化学循环过程与机制(温学发等, 2019).稳定同位素红外光谱技术和方法的进步使碳稳定同位素比值(δ13C)和氧稳定同位素比值(δ18O)(CO2)、δ13C (CH4)、氮稳定同位素比值(δ15N)和δ18O (N2O)、氢稳定同位素比值(δD)和δ18O (H2O)等温室气体同位素组成的连续观测成为可能(Wen et al., 2008, 2013), 与箱式通量观测技术和方法结合可以实现温室气体及其同位素通量观测研究, 有助于揭示温室气体在土壤-植物-大气连续体的产生、传输和转化的相关过程与机制等(Wingate et al., 2010; Bowling et al., 2015; Epron et al., 2016). ...
... 目前, 非稳态和稳态箱式通量观测系统在自然条件以及控制条件下土壤和植物根-茎-叶乃至生态系统尺度CO2及其δ13C通量观测中均得到应用(Griffis, 2013; Bowling et al., 2015; Zhou et al., 2018).商业化及研究者自制了土壤和植物CO2及其δ13C通量协同观测系统(图2A)和生态系统或冠层CO2及其δ13C通量观测系统(图2B), 通常将分析仪及箱式装置有效整合为一套全自动在线连续观测系统.值得注意的是, 如图2所示, 如果观测系统形成闭路循环气路则为非稳态系统, 而形成开路循环气路则为稳态系统. ...
... 土壤CO2及其δ13C通量的原位长期连续观测, 有助于解析土壤呼吸随植物生理和环境等因素变化过程中的微小波动(Midwood et al., 2008; Bowling et al., 2015).土壤呼吸包括自养呼吸和异养呼吸, 而δ13C已经被证明可以用于拆分这两种组分(Midwood et al., 2008; Albanito et al., 2012).Midwood等(2008)利用土壤CO2稳态系统与稳定同位素质谱仪(Isotope Ratio Mass Spectrometry, IRMS)连用测定, 通过两元混合模型计算表明大麦(Hordeum vulgare)幼苗根中50%的碳可能来自于光合作用新合成的碳水化合物.Albanito等(2012)通过设计的参考室和双气室的稳态系统, 利用三元混合模型计算出土壤有机质、凋落物和根对土壤呼吸的相对贡献比例分别为37%、33%和30%.Liu等(2018)利用非稳态系统进行周期性升温和降温(5-30 ℃)培养试验, 结果表明土壤微生物呼吸对升温和降温具有明显的非对称响应, 升温阶段土壤微生物呼吸的增长速度更快. ...
... 根据研究需要, 研究者通常将上述土壤、植物叶片、茎干和根CO2及其δ13C通量观测箱体进行组合, 即可实现土壤和植物CO2及其δ13C通量的协同观测(Wingate et al., 2010; Kuptz et al., 2011a, 2011b).叶片(小枝)、茎干和土壤呼吸CO2及其δ13C协同观测系统可用于植物光合作用新合成碳水化合物的迁移转化过程研究(Bowling et al., 2008; Wingate et al., 2010; Kuptz et al., 2011a, 2011b).叶片(小枝)、茎干和土壤呼吸CO2及其δ13C间具有明显的时间滞后效应(Wingate et al., 2010; Bowling et al., 2015).对20-25 m高的大树研究发现, 土壤CO2 δ13C与冠层光合产物δ13C之间存在1-4天的滞后效应, 并且显著受控于空气相对湿度的变化(Ekblad & H?gberg, 2001).Wingate等(2010)研究发现, 与光合判别相比, 叶片呼吸释放CO2 δ13C的短期变异更小, 并且存在2-10天的延迟效应, 这可能是由于后光合分馏效应和新老碳的混合作用共同导致的. ...
Carbon isotopes in terrestrial ecosystem pools and CO2 fluxes 1 2008
... 根据研究需要, 研究者通常将上述土壤、植物叶片、茎干和根CO2及其δ13C通量观测箱体进行组合, 即可实现土壤和植物CO2及其δ13C通量的协同观测(Wingate et al., 2010; Kuptz et al., 2011a, 2011b).叶片(小枝)、茎干和土壤呼吸CO2及其δ13C协同观测系统可用于植物光合作用新合成碳水化合物的迁移转化过程研究(Bowling et al., 2008; Wingate et al., 2010; Kuptz et al., 2011a, 2011b).叶片(小枝)、茎干和土壤呼吸CO2及其δ13C间具有明显的时间滞后效应(Wingate et al., 2010; Bowling et al., 2015).对20-25 m高的大树研究发现, 土壤CO2 δ13C与冠层光合产物δ13C之间存在1-4天的滞后效应, 并且显著受控于空气相对湿度的变化(Ekblad & H?gberg, 2001).Wingate等(2010)研究发现, 与光合判别相比, 叶片呼吸释放CO2 δ13C的短期变异更小, 并且存在2-10天的延迟效应, 这可能是由于后光合分馏效应和新老碳的混合作用共同导致的. ...
CO2 concentration profiles, and carbon and oxygen isotopes in C3 and C4 crop canopies 1 1998
Intercomparison of three models for δ13C of respiration with four regression approaches 3 2017
... 由于气体分析仪只能测定单位体积湿空气的CO2物质的量, 而不能直接测定单位体积干空气的CO2物质的量, 这里湿空气密度等于干空气密度、水汽密度和CO2密度的和.因此, 需要校正水汽密度变化对CO2密度的影响.非稳态系统CO2通量计算是根据箱体内CO2、水汽和空气的3个质量守恒方程联立推导获得.非稳态系统CO2 δ13C通量计算是根据同位素物质守恒原理, 利用Keeling Plot曲线(KP) (Keeling, 1958), Miller-Tans曲线(MT)(Miller & Tans, 2003)等同位素混合模型以及同位素通量比值(FR)模型(Griffis et al., 2004)计算.上述模型的估算偏差和不确定性受到CO2跨度(KP和MT)和d13CO2/dz (FR)降低的影响(Chen et al., 2017).高斯噪声模拟表明不同模型估算结果的不确定性主要来源于δ13C观测误差, 而非CO2观测误差.确定低CO2跨度或FR阈值是保证不同模型计算精度的有效途径(Chen et al., 2017). ...
... 跨度或FR阈值是保证不同模型计算精度的有效途径(Chen et al., 2017). ...
... 所有箱式通量观测系统均需要对气体分析仪进行定期标定.目前常用的CO2及其δ13C气体分析仪主要基于稳定同位素红外光谱技术(Midwood & Millard, 2011; Griffis, 2013; Wen et al., 2013; Pang et al., 2016a), 但该技术自身也存在缺点, 主要表现为δ13C测量值对CO2浓度变化的非线性响应(浓度依赖性)以及对环境条件变化的敏感性导致的漂移(时间漂移).二者均对气体分析仪的精度和准确度产生影响, 需要进行适当校正和数据溯源以保证数据的可靠性(Kammer et al., 2011; Pang et al., 2016b; Chen et al., 2017, 2019).庞家平和温学发(2018)推荐利用3个或3个以上已知CO2浓度和δ13C真值的CO2标准气体涵盖待测气体CO2浓度, 进行气体分析仪的浓度依赖性校正, 同时通过设置适当的校正频率, 校正时间漂移并进行数据溯源. ...
Photosynthetic carbon isotope discrimination and effects on daytime NEE partitioning in a subtropical mixed conifer plantation 1 2019
... 所有箱式通量观测系统均需要对气体分析仪进行定期标定.目前常用的CO2及其δ13C气体分析仪主要基于稳定同位素红外光谱技术(Midwood & Millard, 2011; Griffis, 2013; Wen et al., 2013; Pang et al., 2016a), 但该技术自身也存在缺点, 主要表现为δ13C测量值对CO2浓度变化的非线性响应(浓度依赖性)以及对环境条件变化的敏感性导致的漂移(时间漂移).二者均对气体分析仪的精度和准确度产生影响, 需要进行适当校正和数据溯源以保证数据的可靠性(Kammer et al., 2011; Pang et al., 2016b; Chen et al., 2017, 2019).庞家平和温学发(2018)推荐利用3个或3个以上已知CO2浓度和δ13C真值的CO2标准气体涵盖待测气体CO2浓度, 进行气体分析仪的浓度依赖性校正, 同时通过设置适当的校正频率, 校正时间漂移并进行数据溯源. ...
Minimizing artifacts and biases in chamber-based measurements of soil respiration 2 2002
... 测定过程中, 不能改变待测气体源汇的底物特征及环境条件, 影响待测气体的产生.土壤呼吸依赖于温度和湿度等环境因素, 人为的环境条件改变影响CO2通量(Davidson et al., 2002).测定过程应尽量减少对气室内温度和湿度等条件的影响.土壤环插入土壤的过程会切断土壤中的根系.通常在测定前尽量长的时间将土壤环安装到样地, 使得生态系统能够得到有效恢复; 在不影响系统整体性的前提下, 土壤环插入深度都尽量浅(Midwood & Millard, 2011; 李建军等, 2019), 但存在表层土壤空气流动的问题(Parkin et al., 2005).风不仅能引起气室内压力变化, 也会促进大气CO2与表层土壤CO2的混合(Susfalk et al., 2002; Millard et al., 2008).土壤环插入土壤深度过浅可能会存在大气CO2与表层土壤CO2的混合, 特别是对于干燥、多空隙的土壤来说, 会引入CO2浓度测定误差, 并显著改变测定的δ13C值(Susfalk et al., 2002; Millard et al., 2008).Hutchinson和Livingston (2001)的研究表明, 干燥、多气孔或者粗颗粒的土壤需将土壤环插入土壤超过10 cm; 而细颗粒、湿润或紧实的土壤只需将土壤环插入土壤不超过5 cm.Nickerson和Risk (2009)利用同位素模型模拟了正确的土壤环插入深度对于阻止空气进入气室的重要性, 最差的情况下, 气室内CO2的δ13C富集可达到4‰. ...
Mesophyll conductance to CO2, assessed from online TDL-AS records of 13CO2 discrimination, displays small but significant short-term responses to CO2 and irradiance in Eucalyptus seedlings 2 2011
... 相对于非稳态箱式通量观测系统, 稳态箱式通量观测系统应用更加广泛, 可以采用明箱或暗箱测定.商业化叶片CO2箱式通量观测系统包括LI-6400/6800等, 为稳态系统.通过红外光谱技术对气室内样品气和参考气进行差分测量, 计算CO2通量(Barbour et al., 2007; Douthe et al., 2011).叶片与大气之间的CO2平衡和水汽交换受到气孔导度的控制.随着研究的深入, 发现叶片内部传输过程对碳水交换的影响也非常重要(Moreno-Gutiérrez et al., 2012).叶片CO2 δ13C通量连续观测系统可通过从LI-6400/6800气室进气口和出气口分别抽取部分气体进入稳定同位素红外光谱仪, 即可同步测定叶片CO2及其δ13C通量(Douthe et al., 2011; Barbour et al., 2016), 这为理解CO2在叶片内部传输的动态过程提供了技术支撑, 可揭示叶肉导度对CO2导度和水汽导度之间存在的权衡关系(Moreno-Gutiérrez et al., 2012). ...
... C通量(Douthe et al., 2011; Barbour et al., 2016), 这为理解CO2在叶片内部传输的动态过程提供了技术支撑, 可揭示叶肉导度对CO2导度和水汽导度之间存在的权衡关系(Moreno-Gutiérrez et al., 2012). ...
Oxygen isotope signatures of transpired water vapor: the role of isotopic non-steady-state transpiration under natural conditions 1 2014
... 由于原理简单且操作方便, 箱式通量观测技术和方法在自然或控制条件下的土壤、植物乃至生态系统尺度温室气体通量观测研究中得到广泛应用(Kuptz et al., 2011a, 2011b; He et al., 2013; Merbold et al., 2013; Savage et al., 2014).从早期的静态箱-碱液吸收滴定(Lundegardh, 1927)和静态箱-气相色谱测定(Midwood & Millard, 2011; Imer et al., 2013)等人工离线测定模式, 发展到目前通常采用的自动箱-红外光谱测定(Plain et al., 2009; Dubbert et al., 2014; Savage et al., 2014)等自动在线测定模式.但是, 通常缺乏对观测系统设计的理论要求和假设等问题的综合考虑和评价.首先, 如何保证系统测定数据的精度和准确度, 即保证系统气密性及气体混合、箱体内外气体浓度和压力保持一致等(Xu et al., 2006; Midwood et al., 2008); 其次, 如何保证系统测定数据的代表性, 即待测气体的产生和传输不受影响等(Midwood et al., 2008; Weiss et al., 2009; Snell et al., 2014). ...
Natural abundance of 13C in CO2 respired from forest soils reveals speed of link between tree photosynthesis and root respiration 1 2001
... 根据研究需要, 研究者通常将上述土壤、植物叶片、茎干和根CO2及其δ13C通量观测箱体进行组合, 即可实现土壤和植物CO2及其δ13C通量的协同观测(Wingate et al., 2010; Kuptz et al., 2011a, 2011b).叶片(小枝)、茎干和土壤呼吸CO2及其δ13C协同观测系统可用于植物光合作用新合成碳水化合物的迁移转化过程研究(Bowling et al., 2008; Wingate et al., 2010; Kuptz et al., 2011a, 2011b).叶片(小枝)、茎干和土壤呼吸CO2及其δ13C间具有明显的时间滞后效应(Wingate et al., 2010; Bowling et al., 2015).对20-25 m高的大树研究发现, 土壤CO2 δ13C与冠层光合产物δ13C之间存在1-4天的滞后效应, 并且显著受控于空气相对湿度的变化(Ekblad & H?gberg, 2001).Wingate等(2010)研究发现, 与光合判别相比, 叶片呼吸释放CO2 δ13C的短期变异更小, 并且存在2-10天的延迟效应, 这可能是由于后光合分馏效应和新老碳的混合作用共同导致的. ...
In situ13CO2 pulse labelling of field-grown eucalypt trees revealed the effects of potassium nutrition and throughfall exclusion on phloem transport of photosynthetic carbon 6 2016
... 生态系统各种生源元素在土壤、植物、动物、微生物和大气间迁移和转化的物质循环过程研究是生态系统生态学的前沿性科学问题(于贵瑞, 2009; 于振良, 2017).碳循环、氮循环和水循环是最重要的物质循环, 碳(CO2、CH4)、氮(N2O)和水汽(H2O)等温室气体的交换通量是生态系统物质循环的核心, 是地圈-生物圈-大气圈相互作用的纽带(于贵瑞和孙晓敏, 2017).同位素技术具有指示、示踪和整合功能, 可以辅助解析生态系统碳氮水的生物地球化学循环过程与机制(温学发等, 2019).稳定同位素红外光谱技术和方法的进步使碳稳定同位素比值(δ13C)和氧稳定同位素比值(δ18O)(CO2)、δ13C (CH4)、氮稳定同位素比值(δ15N)和δ18O (N2O)、氢稳定同位素比值(δD)和δ18O (H2O)等温室气体同位素组成的连续观测成为可能(Wen et al., 2008, 2013), 与箱式通量观测技术和方法结合可以实现温室气体及其同位素通量观测研究, 有助于揭示温室气体在土壤-植物-大气连续体的产生、传输和转化的相关过程与机制等(Wingate et al., 2010; Bowling et al., 2015; Epron et al., 2016). ...
... 所有箱式通量观测系统均需要对系统测定数据的精度和准确度进行评价.可以通过零通量和模拟通量测试对箱式通量观测系统测定数据的精度和准度进行评价(Midwood et al., 2008; Powers et al., 2010; 高峻等, 2011; 于贵瑞等, 2015).零通量测试与系统漏率测试类似, 但不需要使用标气, 只要将气室箱体密封进行正常测定即可, 然后计算CO2通量评价系统是否满足气密性的基本需求.模拟通量测试需要利用注射泵向模拟装置中匀速地注入标准气体来模拟土壤气体的排放过程, 将实际测定排放速率与模拟通量的设定值对比分析, 即可验证观测系统的准确度和精度. ...
Tracing the flow of carbon dioxide and water vapor between the biosphere and atmosphere: a review of optical isotope techniques and their application 3 2013
... 目前, 非稳态和稳态箱式通量观测系统在自然条件以及控制条件下土壤和植物根-茎-叶乃至生态系统尺度CO2及其δ13C通量观测中均得到应用(Griffis, 2013; Bowling et al., 2015; Zhou et al., 2018).商业化及研究者自制了土壤和植物CO2及其δ13C通量协同观测系统(图2A)和生态系统或冠层CO2及其δ13C通量观测系统(图2B), 通常将分析仪及箱式装置有效整合为一套全自动在线连续观测系统.值得注意的是, 如图2所示, 如果观测系统形成闭路循环气路则为非稳态系统, 而形成开路循环气路则为稳态系统. ...
... 所有箱式通量观测系统均需要对气体分析仪进行定期标定.目前常用的CO2及其δ13C气体分析仪主要基于稳定同位素红外光谱技术(Midwood & Millard, 2011; Griffis, 2013; Wen et al., 2013; Pang et al., 2016a), 但该技术自身也存在缺点, 主要表现为δ13C测量值对CO2浓度变化的非线性响应(浓度依赖性)以及对环境条件变化的敏感性导致的漂移(时间漂移).二者均对气体分析仪的精度和准确度产生影响, 需要进行适当校正和数据溯源以保证数据的可靠性(Kammer et al., 2011; Pang et al., 2016b; Chen et al., 2017, 2019).庞家平和温学发(2018)推荐利用3个或3个以上已知CO2浓度和δ13C真值的CO2标准气体涵盖待测气体CO2浓度, 进行气体分析仪的浓度依赖性校正, 同时通过设置适当的校正频率, 校正时间漂移并进行数据溯源. ...
Measuring field-scale isotopic CO2 fluxes with tunable diode laser absorption spectroscopy and micrometeorological techniques 2 2004
... 由于气体分析仪只能测定单位体积湿空气的CO2物质的量, 而不能直接测定单位体积干空气的CO2物质的量, 这里湿空气密度等于干空气密度、水汽密度和CO2密度的和.因此, 需要校正水汽密度变化对CO2密度的影响.非稳态系统CO2通量计算是根据箱体内CO2、水汽和空气的3个质量守恒方程联立推导获得.非稳态系统CO2 δ13C通量计算是根据同位素物质守恒原理, 利用Keeling Plot曲线(KP) (Keeling, 1958), Miller-Tans曲线(MT)(Miller & Tans, 2003)等同位素混合模型以及同位素通量比值(FR)模型(Griffis et al., 2004)计算.上述模型的估算偏差和不确定性受到CO2跨度(KP和MT)和d13CO2/dz (FR)降低的影响(Chen et al., 2017).高斯噪声模拟表明不同模型估算结果的不确定性主要来源于δ13C观测误差, 而非CO2观测误差.确定低CO2跨度或FR阈值是保证不同模型计算精度的有效途径(Chen et al., 2017). ...
Effect of grazing on carbon stocks and assimilate partitioning in a Tibetan montane pasture revealed by 13CO2 pulse labeling 1 2012
... 生态系统箱式通量观测系统主要分为两种: 冠层通量测定(不包括土壤)和生态系统通量测定(包括土壤), 可以采用明箱或暗箱测定.生态系统通量观测多应用于草本或小灌木生态系统通量观测研究(Wu et al., 2010; Hafner et al., 2012; Wei et al., 2016).高大乔木根据研究目标可以选择进行冠层或生态系统CO2及其δ13C通量观测.但由于难度较高而选择进行13CO2脉冲标记研究(Plain et al., 2009; Epron et al., 2016).通常利用不同高度的钢制脚手架环绕树的冠层, 利用高透光率的聚合物膜或有机玻璃等制作冠层箱(明箱或暗箱), 箱体的底座由两个半圆形不锈钢板制成, 钢板中间开口, 以便容纳茎干, 并固定在脚手架上(Plain et al., 2009; Epron et al., 2016).为了维持箱体内的温湿度条件, 通常配套安装空调, 利用风扇混匀冠层箱内的气体(Epron et al., 2016).Epron等(2016)利用27或38 m3的高透光率膜罩住整个桉树(Eucalyptus robusta)冠层进行脉冲标记, 利用茎干和根CO2及其δ13C通量观测箱实时观测, 结果表明, 叶片中标记的碳的储存时间非常短(21-31 h), 其在茎干中的迁移速率为0.20-0.82 m·h-1. ...
Root carbon flux: measurements versus mechanisms 1 2009
... 根据物质守恒方程, 要求待测气体传输只受扩散过程影响, 且全部扩散至气室箱体内, 同时测定过程不能影响CO2产生和传输过程, 否则测定的CO2释放量并不能代表呼吸通量(Midwood et al., 2008; Powers et al., 2010; Kammer et al., 2011; Midwood & Millard, 2011; Sturm et al., 2013; Savage et al., 2014).首先, 应该保证测定过程不影响待测气体的产生和传输, 即不改变温湿度等环境条件以及待测气体源汇底物特征, 并且有效消除气室箱体的边缘效应, 以及不影响气体的扩散和非扩散过程.其次, 客观验证和讨论待测气体传输只受扩散过程影响且全部扩散至气室箱体内的科学假设是否成立.例如, 土壤呼吸产生的CO2主要包括地表释放的CO2和土壤中储存的CO2两部分(Hanson & Gunderson, 2009; Midwood & Millard, 2011).通常假设土壤中储存的CO2通量可以忽略不计, 则箱式通量观测技术和方法(箱式法)直接测定的“土壤CO2释放量”等于“土壤呼吸” (Maier & Schack-Kirchner, 2014).如果土壤CO2在土壤中发生明显的储存(在某些情况下浓度高达数万μmol·mol-1), 那么就会显著影响箱式法的观测结果, 导致数据不具有代表性(Wang et al., 2018a, 2018b).土壤箱式通量法可与土壤通量梯度法相结合, 实现土壤CO2释放量的相互验证以及土壤CO2储存通量的估算(Maier et al., 2011; Maier & Schack-Kirchner, 2014). ...
Changes in the temperature sensitivity of SOM decomposition with grassland succession: implications for soil C sequestration 1 2013
... 由于原理简单且操作方便, 箱式通量观测技术和方法在自然或控制条件下的土壤、植物乃至生态系统尺度温室气体通量观测研究中得到广泛应用(Kuptz et al., 2011a, 2011b; He et al., 2013; Merbold et al., 2013; Savage et al., 2014).从早期的静态箱-碱液吸收滴定(Lundegardh, 1927)和静态箱-气相色谱测定(Midwood & Millard, 2011; Imer et al., 2013)等人工离线测定模式, 发展到目前通常采用的自动箱-红外光谱测定(Plain et al., 2009; Dubbert et al., 2014; Savage et al., 2014)等自动在线测定模式.但是, 通常缺乏对观测系统设计的理论要求和假设等问题的综合考虑和评价.首先, 如何保证系统测定数据的精度和准确度, 即保证系统气密性及气体混合、箱体内外气体浓度和压力保持一致等(Xu et al., 2006; Midwood et al., 2008); 其次, 如何保证系统测定数据的代表性, 即待测气体的产生和传输不受影响等(Midwood et al., 2008; Weiss et al., 2009; Snell et al., 2014). ...
Measuring the ratio of CO2 efflux to O2 influx in tree stem respiration 1 2016
... 茎干箱式通量观测系统通常可以直接使用或借鉴土壤箱式通量观测系统.根据胸径大小制作特定直径的PVC环, 密封固定在树干上直接利用土壤箱式通量观测系统进行观测(Yang et al., 2012; Hilman & Angert, 2016; Salomón et al., 2016), 或者制作适合茎干的特定箱体并与气体分析仪相连构成茎干箱式通量观测系统(Wang et al., 2019; Yang et al., 2019).茎干CO2通量是森林生态系统碳循环中的重要组成部分, 占全球森林生态系统净初级生产力的20% (Yang et al., 2016, 2019; Zhao et al., 2018).研究表明, 茎干呼吸的日变化趋势呈现单峰曲线、双峰曲线和S型曲线3种模式(石新立等, 2010).其中, 气候和植被类型等可能是不同变化趋势的决定因素.Yang等(2016)通过收集全球121篇已发表文章的数据, 研究了茎干呼吸与叶面积指数、年平均气温及年平均降水量的相关关系, 发现茎干呼吸与叶面积指数呈正相关关系, 叶面积指数、年平均气温及降水量可以解释茎干呼吸变异的74%. ...
Ecosystem-level controls on root-rhizosphere respiration 1 2013
... 相对于土壤、植物叶片和茎干箱式通量观测系统, 根系箱式通量观测系统的难点在于适合于根系的气室箱体设计和实现(Marsden et al., 2008; Jia et al., 2013; Lai et al., 2015, 2016).传统根CO2通量观测通常将根系切断, 然后放到根呼吸气室箱体内, 但被切除的根系呼吸速率明显高于未被切除的根系(Makita et al., 2009, 2013; Sun et al., 2017).而通过原位根呼吸气室箱体设计可以很好地解决这一问题, 根呼吸箱通常使用PVC材料, 由两个半圆环或者一个封闭圆环(一端开口)构成, 将不同直径的根系放进根呼吸箱后, 形成封闭系统用于观测根呼吸CO2通量(Marsden et al., 2008; Kuptz et al., 2011a, 2011b).根系呼吸是森林土壤碳排放的重要来源, 其对土壤呼吸的贡献比例介于1/3到50%多之间(Marsden et al., 2008; Makita et al., 2009, 2012; Sun et al., 2017).结合新的高时间分辨率土壤呼吸数据集, 可以为土壤和根系呼吸的生物和环境控制机制提供新的见解(Hopkins et al., 2013).植物根系具有不同直径(粗跟和细根)和不同生理性状, 其呼吸速率模式及其对环境因子的响应具有明显差异.根系呼吸与根系直径、组织组成和特定根长等形态特征密切相关(Sun & Mao, 2011).通过根系呼吸与形态性状之间的相关关系, 可以建立区分粗根和细根呼吸的定量关系.细根对于植物生长、营养元素获取等生理功能具有重要作用, 其呼吸速率通常高于粗根, 且变异较大.在野外原位观测的结果发现, 根系直径与呼吸速率显著相关, 直径小于0.4 mm的根系呼吸速率显著高于其他根系, 且变异较大; 同时, 根系呼吸速率与根长和根的含氮量呈正相关关系(Makita et al., 2009). ...
Vents and seals in non-steady-state chambers used for measuring gas exchange between soil and the atmosphere 2 2001
... 测定过程中, 不能改变待测气体源汇的底物特征及环境条件, 影响待测气体的产生.土壤呼吸依赖于温度和湿度等环境因素, 人为的环境条件改变影响CO2通量(Davidson et al., 2002).测定过程应尽量减少对气室内温度和湿度等条件的影响.土壤环插入土壤的过程会切断土壤中的根系.通常在测定前尽量长的时间将土壤环安装到样地, 使得生态系统能够得到有效恢复; 在不影响系统整体性的前提下, 土壤环插入深度都尽量浅(Midwood & Millard, 2011; 李建军等, 2019), 但存在表层土壤空气流动的问题(Parkin et al., 2005).风不仅能引起气室内压力变化, 也会促进大气CO2与表层土壤CO2的混合(Susfalk et al., 2002; Millard et al., 2008).土壤环插入土壤深度过浅可能会存在大气CO2与表层土壤CO2的混合, 特别是对于干燥、多空隙的土壤来说, 会引入CO2浓度测定误差, 并显著改变测定的δ13C值(Susfalk et al., 2002; Millard et al., 2008).Hutchinson和Livingston (2001)的研究表明, 干燥、多气孔或者粗颗粒的土壤需将土壤环插入土壤超过10 cm; 而细颗粒、湿润或紧实的土壤只需将土壤环插入土壤不超过5 cm.Nickerson和Risk (2009)利用同位素模型模拟了正确的土壤环插入深度对于阻止空气进入气室的重要性, 最差的情况下, 气室内CO2的δ13C富集可达到4‰. ...
Temporal and spatial variations of soil CO2, CH4 and N2O fluxes at three differently managed grasslands 1 2013
... 由于原理简单且操作方便, 箱式通量观测技术和方法在自然或控制条件下的土壤、植物乃至生态系统尺度温室气体通量观测研究中得到广泛应用(Kuptz et al., 2011a, 2011b; He et al., 2013; Merbold et al., 2013; Savage et al., 2014).从早期的静态箱-碱液吸收滴定(Lundegardh, 1927)和静态箱-气相色谱测定(Midwood & Millard, 2011; Imer et al., 2013)等人工离线测定模式, 发展到目前通常采用的自动箱-红外光谱测定(Plain et al., 2009; Dubbert et al., 2014; Savage et al., 2014)等自动在线测定模式.但是, 通常缺乏对观测系统设计的理论要求和假设等问题的综合考虑和评价.首先, 如何保证系统测定数据的精度和准确度, 即保证系统气密性及气体混合、箱体内外气体浓度和压力保持一致等(Xu et al., 2006; Midwood et al., 2008); 其次, 如何保证系统测定数据的代表性, 即待测气体的产生和传输不受影响等(Midwood et al., 2008; Weiss et al., 2009; Snell et al., 2014). ...
Relationships between root respiration rate and root morphology, chemistry and anatomy in Larix gmelinii and Fraxinus mandshurica 1 2013
... 相对于土壤、植物叶片和茎干箱式通量观测系统, 根系箱式通量观测系统的难点在于适合于根系的气室箱体设计和实现(Marsden et al., 2008; Jia et al., 2013; Lai et al., 2015, 2016).传统根CO2通量观测通常将根系切断, 然后放到根呼吸气室箱体内, 但被切除的根系呼吸速率明显高于未被切除的根系(Makita et al., 2009, 2013; Sun et al., 2017).而通过原位根呼吸气室箱体设计可以很好地解决这一问题, 根呼吸箱通常使用PVC材料, 由两个半圆环或者一个封闭圆环(一端开口)构成, 将不同直径的根系放进根呼吸箱后, 形成封闭系统用于观测根呼吸CO2通量(Marsden et al., 2008; Kuptz et al., 2011a, 2011b).根系呼吸是森林土壤碳排放的重要来源, 其对土壤呼吸的贡献比例介于1/3到50%多之间(Marsden et al., 2008; Makita et al., 2009, 2012; Sun et al., 2017).结合新的高时间分辨率土壤呼吸数据集, 可以为土壤和根系呼吸的生物和环境控制机制提供新的见解(Hopkins et al., 2013).植物根系具有不同直径(粗跟和细根)和不同生理性状, 其呼吸速率模式及其对环境因子的响应具有明显差异.根系呼吸与根系直径、组织组成和特定根长等形态特征密切相关(Sun & Mao, 2011).通过根系呼吸与形态性状之间的相关关系, 可以建立区分粗根和细根呼吸的定量关系.细根对于植物生长、营养元素获取等生理功能具有重要作用, 其呼吸速率通常高于粗根, 且变异较大.在野外原位观测的结果发现, 根系直径与呼吸速率显著相关, 直径小于0.4 mm的根系呼吸速率显著高于其他根系, 且变异较大; 同时, 根系呼吸速率与根长和根的含氮量呈正相关关系(Makita et al., 2009). ...
Application of a quantum cascade laser- based spectrometer in a closed chamber system for real- time δ13C and δ18O measurements of soil-respired CO2 3 2011
... 箱式通量观测系统需要保证气体分析仪以及系统气路的气密性, 即不存在与外界大气的气体交换过程.测定过程中, 需要箱体内以及系统内气体充分混合才能代表待测气体.通常采取在气室箱体顶端安装风扇方式进行气体混匀(Midwood et al., 2008).但是由于风扇可能导致气室内压力波动, 影响CO2扩散梯度, 进而导致CO2排放通量的高估或低估(Midwood et al., 2008).通常仅通过气室箱体与气体分析仪间的气体循环过程实现系统内气体充分混合(Pumpanen et al., 2004; Kammer et al., 2011).需要注意的是, 开始测量时, 系统内均会残留上一个测定的气体, 需要短暂的系统平衡时间(图1B、1D), 才能实现系统内的气体充分混合.需要保证不同气室箱体间的有效切换才能代表待测气体, 即需要剔除气路切换后的无效数据后保证足够的有效数据量.无效数据量所对应的时间越短代表切换效率越高.同时, 对于CO2 δ13C通量测定, 需要选用不会造成同位素分馏效应的材料构建系统. ...
... 所有箱式通量观测系统均需要对气体分析仪进行定期标定.目前常用的CO2及其δ13C气体分析仪主要基于稳定同位素红外光谱技术(Midwood & Millard, 2011; Griffis, 2013; Wen et al., 2013; Pang et al., 2016a), 但该技术自身也存在缺点, 主要表现为δ13C测量值对CO2浓度变化的非线性响应(浓度依赖性)以及对环境条件变化的敏感性导致的漂移(时间漂移).二者均对气体分析仪的精度和准确度产生影响, 需要进行适当校正和数据溯源以保证数据的可靠性(Kammer et al., 2011; Pang et al., 2016b; Chen et al., 2017, 2019).庞家平和温学发(2018)推荐利用3个或3个以上已知CO2浓度和δ13C真值的CO2标准气体涵盖待测气体CO2浓度, 进行气体分析仪的浓度依赖性校正, 同时通过设置适当的校正频率, 校正时间漂移并进行数据溯源. ...
... 根据物质守恒方程, 要求待测气体传输只受扩散过程影响, 且全部扩散至气室箱体内, 同时测定过程不能影响CO2产生和传输过程, 否则测定的CO2释放量并不能代表呼吸通量(Midwood et al., 2008; Powers et al., 2010; Kammer et al., 2011; Midwood & Millard, 2011; Sturm et al., 2013; Savage et al., 2014).首先, 应该保证测定过程不影响待测气体的产生和传输, 即不改变温湿度等环境条件以及待测气体源汇底物特征, 并且有效消除气室箱体的边缘效应, 以及不影响气体的扩散和非扩散过程.其次, 客观验证和讨论待测气体传输只受扩散过程影响且全部扩散至气室箱体内的科学假设是否成立.例如, 土壤呼吸产生的CO2主要包括地表释放的CO2和土壤中储存的CO2两部分(Hanson & Gunderson, 2009; Midwood & Millard, 2011).通常假设土壤中储存的CO2通量可以忽略不计, 则箱式通量观测技术和方法(箱式法)直接测定的“土壤CO2释放量”等于“土壤呼吸” (Maier & Schack-Kirchner, 2014).如果土壤CO2在土壤中发生明显的储存(在某些情况下浓度高达数万μmol·mol-1), 那么就会显著影响箱式法的观测结果, 导致数据不具有代表性(Wang et al., 2018a, 2018b).土壤箱式通量法可与土壤通量梯度法相结合, 实现土壤CO2释放量的相互验证以及土壤CO2储存通量的估算(Maier et al., 2011; Maier & Schack-Kirchner, 2014). ...
The concentration and isotopic abundances of atmospheric carbon dioxide in rural areas 1 1958
... 由于气体分析仪只能测定单位体积湿空气的CO2物质的量, 而不能直接测定单位体积干空气的CO2物质的量, 这里湿空气密度等于干空气密度、水汽密度和CO2密度的和.因此, 需要校正水汽密度变化对CO2密度的影响.非稳态系统CO2通量计算是根据箱体内CO2、水汽和空气的3个质量守恒方程联立推导获得.非稳态系统CO2 δ13C通量计算是根据同位素物质守恒原理, 利用Keeling Plot曲线(KP) (Keeling, 1958), Miller-Tans曲线(MT)(Miller & Tans, 2003)等同位素混合模型以及同位素通量比值(FR)模型(Griffis et al., 2004)计算.上述模型的估算偏差和不确定性受到CO2跨度(KP和MT)和d13CO2/dz (FR)降低的影响(Chen et al., 2017).高斯噪声模拟表明不同模型估算结果的不确定性主要来源于δ13C观测误差, 而非CO2观测误差.确定低CO2跨度或FR阈值是保证不同模型计算精度的有效途径(Chen et al., 2017). ...
Seasonal patterns of carbon allocation to respiratory pools in 60-yr-old deciduous (Fagus sylvatica) and evergreen (Picea abies) trees assessed via whole-tree stable carbon isotope labeling 4 2011a
... 由于原理简单且操作方便, 箱式通量观测技术和方法在自然或控制条件下的土壤、植物乃至生态系统尺度温室气体通量观测研究中得到广泛应用(Kuptz et al., 2011a, 2011b; He et al., 2013; Merbold et al., 2013; Savage et al., 2014).从早期的静态箱-碱液吸收滴定(Lundegardh, 1927)和静态箱-气相色谱测定(Midwood & Millard, 2011; Imer et al., 2013)等人工离线测定模式, 发展到目前通常采用的自动箱-红外光谱测定(Plain et al., 2009; Dubbert et al., 2014; Savage et al., 2014)等自动在线测定模式.但是, 通常缺乏对观测系统设计的理论要求和假设等问题的综合考虑和评价.首先, 如何保证系统测定数据的精度和准确度, 即保证系统气密性及气体混合、箱体内外气体浓度和压力保持一致等(Xu et al., 2006; Midwood et al., 2008); 其次, 如何保证系统测定数据的代表性, 即待测气体的产生和传输不受影响等(Midwood et al., 2008; Weiss et al., 2009; Snell et al., 2014). ...
... 相对于土壤、植物叶片和茎干箱式通量观测系统, 根系箱式通量观测系统的难点在于适合于根系的气室箱体设计和实现(Marsden et al., 2008; Jia et al., 2013; Lai et al., 2015, 2016).传统根CO2通量观测通常将根系切断, 然后放到根呼吸气室箱体内, 但被切除的根系呼吸速率明显高于未被切除的根系(Makita et al., 2009, 2013; Sun et al., 2017).而通过原位根呼吸气室箱体设计可以很好地解决这一问题, 根呼吸箱通常使用PVC材料, 由两个半圆环或者一个封闭圆环(一端开口)构成, 将不同直径的根系放进根呼吸箱后, 形成封闭系统用于观测根呼吸CO2通量(Marsden et al., 2008; Kuptz et al., 2011a, 2011b).根系呼吸是森林土壤碳排放的重要来源, 其对土壤呼吸的贡献比例介于1/3到50%多之间(Marsden et al., 2008; Makita et al., 2009, 2012; Sun et al., 2017).结合新的高时间分辨率土壤呼吸数据集, 可以为土壤和根系呼吸的生物和环境控制机制提供新的见解(Hopkins et al., 2013).植物根系具有不同直径(粗跟和细根)和不同生理性状, 其呼吸速率模式及其对环境因子的响应具有明显差异.根系呼吸与根系直径、组织组成和特定根长等形态特征密切相关(Sun & Mao, 2011).通过根系呼吸与形态性状之间的相关关系, 可以建立区分粗根和细根呼吸的定量关系.细根对于植物生长、营养元素获取等生理功能具有重要作用, 其呼吸速率通常高于粗根, 且变异较大.在野外原位观测的结果发现, 根系直径与呼吸速率显著相关, 直径小于0.4 mm的根系呼吸速率显著高于其他根系, 且变异较大; 同时, 根系呼吸速率与根长和根的含氮量呈正相关关系(Makita et al., 2009). ...
... 根据研究需要, 研究者通常将上述土壤、植物叶片、茎干和根CO2及其δ13C通量观测箱体进行组合, 即可实现土壤和植物CO2及其δ13C通量的协同观测(Wingate et al., 2010; Kuptz et al., 2011a, 2011b).叶片(小枝)、茎干和土壤呼吸CO2及其δ13C协同观测系统可用于植物光合作用新合成碳水化合物的迁移转化过程研究(Bowling et al., 2008; Wingate et al., 2010; Kuptz et al., 2011a, 2011b).叶片(小枝)、茎干和土壤呼吸CO2及其δ13C间具有明显的时间滞后效应(Wingate et al., 2010; Bowling et al., 2015).对20-25 m高的大树研究发现, 土壤CO2 δ13C与冠层光合产物δ13C之间存在1-4天的滞后效应, 并且显著受控于空气相对湿度的变化(Ekblad & H?gberg, 2001).Wingate等(2010)研究发现, 与光合判别相比, 叶片呼吸释放CO2 δ13C的短期变异更小, 并且存在2-10天的延迟效应, 这可能是由于后光合分馏效应和新老碳的混合作用共同导致的. ...
... ., 2010; Kuptz et al., 2011a, 2011b).叶片(小枝)、茎干和土壤呼吸CO2及其δ13C间具有明显的时间滞后效应(Wingate et al., 2010; Bowling et al., 2015).对20-25 m高的大树研究发现, 土壤CO2 δ13C与冠层光合产物δ13C之间存在1-4天的滞后效应, 并且显著受控于空气相对湿度的变化(Ekblad & H?gberg, 2001).Wingate等(2010)研究发现, 与光合判别相比, 叶片呼吸释放CO2 δ13C的短期变异更小, 并且存在2-10天的延迟效应, 这可能是由于后光合分馏效应和新老碳的混合作用共同导致的. ...
Seasonal dynamics in the stable carbon isotope composition δ13C from non-leafy branch, trunk and coarse root CO2 efflux of adult deciduous (Fagus sylvatica) and evergreen (Picea abies) trees 4 2011b
... 由于原理简单且操作方便, 箱式通量观测技术和方法在自然或控制条件下的土壤、植物乃至生态系统尺度温室气体通量观测研究中得到广泛应用(Kuptz et al., 2011a, 2011b; He et al., 2013; Merbold et al., 2013; Savage et al., 2014).从早期的静态箱-碱液吸收滴定(Lundegardh, 1927)和静态箱-气相色谱测定(Midwood & Millard, 2011; Imer et al., 2013)等人工离线测定模式, 发展到目前通常采用的自动箱-红外光谱测定(Plain et al., 2009; Dubbert et al., 2014; Savage et al., 2014)等自动在线测定模式.但是, 通常缺乏对观测系统设计的理论要求和假设等问题的综合考虑和评价.首先, 如何保证系统测定数据的精度和准确度, 即保证系统气密性及气体混合、箱体内外气体浓度和压力保持一致等(Xu et al., 2006; Midwood et al., 2008); 其次, 如何保证系统测定数据的代表性, 即待测气体的产生和传输不受影响等(Midwood et al., 2008; Weiss et al., 2009; Snell et al., 2014). ...
... 相对于土壤、植物叶片和茎干箱式通量观测系统, 根系箱式通量观测系统的难点在于适合于根系的气室箱体设计和实现(Marsden et al., 2008; Jia et al., 2013; Lai et al., 2015, 2016).传统根CO2通量观测通常将根系切断, 然后放到根呼吸气室箱体内, 但被切除的根系呼吸速率明显高于未被切除的根系(Makita et al., 2009, 2013; Sun et al., 2017).而通过原位根呼吸气室箱体设计可以很好地解决这一问题, 根呼吸箱通常使用PVC材料, 由两个半圆环或者一个封闭圆环(一端开口)构成, 将不同直径的根系放进根呼吸箱后, 形成封闭系统用于观测根呼吸CO2通量(Marsden et al., 2008; Kuptz et al., 2011a, 2011b).根系呼吸是森林土壤碳排放的重要来源, 其对土壤呼吸的贡献比例介于1/3到50%多之间(Marsden et al., 2008; Makita et al., 2009, 2012; Sun et al., 2017).结合新的高时间分辨率土壤呼吸数据集, 可以为土壤和根系呼吸的生物和环境控制机制提供新的见解(Hopkins et al., 2013).植物根系具有不同直径(粗跟和细根)和不同生理性状, 其呼吸速率模式及其对环境因子的响应具有明显差异.根系呼吸与根系直径、组织组成和特定根长等形态特征密切相关(Sun & Mao, 2011).通过根系呼吸与形态性状之间的相关关系, 可以建立区分粗根和细根呼吸的定量关系.细根对于植物生长、营养元素获取等生理功能具有重要作用, 其呼吸速率通常高于粗根, 且变异较大.在野外原位观测的结果发现, 根系直径与呼吸速率显著相关, 直径小于0.4 mm的根系呼吸速率显著高于其他根系, 且变异较大; 同时, 根系呼吸速率与根长和根的含氮量呈正相关关系(Makita et al., 2009). ...
... 根据研究需要, 研究者通常将上述土壤、植物叶片、茎干和根CO2及其δ13C通量观测箱体进行组合, 即可实现土壤和植物CO2及其δ13C通量的协同观测(Wingate et al., 2010; Kuptz et al., 2011a, 2011b).叶片(小枝)、茎干和土壤呼吸CO2及其δ13C协同观测系统可用于植物光合作用新合成碳水化合物的迁移转化过程研究(Bowling et al., 2008; Wingate et al., 2010; Kuptz et al., 2011a, 2011b).叶片(小枝)、茎干和土壤呼吸CO2及其δ13C间具有明显的时间滞后效应(Wingate et al., 2010; Bowling et al., 2015).对20-25 m高的大树研究发现, 土壤CO2 δ13C与冠层光合产物δ13C之间存在1-4天的滞后效应, 并且显著受控于空气相对湿度的变化(Ekblad & H?gberg, 2001).Wingate等(2010)研究发现, 与光合判别相比, 叶片呼吸释放CO2 δ13C的短期变异更小, 并且存在2-10天的延迟效应, 这可能是由于后光合分馏效应和新老碳的混合作用共同导致的. ...
Diel patterns of fine root respiration in a dryland shrub, measured in situ over different phenological stages 1 2016
... 相对于土壤、植物叶片和茎干箱式通量观测系统, 根系箱式通量观测系统的难点在于适合于根系的气室箱体设计和实现(Marsden et al., 2008; Jia et al., 2013; Lai et al., 2015, 2016).传统根CO2通量观测通常将根系切断, 然后放到根呼吸气室箱体内, 但被切除的根系呼吸速率明显高于未被切除的根系(Makita et al., 2009, 2013; Sun et al., 2017).而通过原位根呼吸气室箱体设计可以很好地解决这一问题, 根呼吸箱通常使用PVC材料, 由两个半圆环或者一个封闭圆环(一端开口)构成, 将不同直径的根系放进根呼吸箱后, 形成封闭系统用于观测根呼吸CO2通量(Marsden et al., 2008; Kuptz et al., 2011a, 2011b).根系呼吸是森林土壤碳排放的重要来源, 其对土壤呼吸的贡献比例介于1/3到50%多之间(Marsden et al., 2008; Makita et al., 2009, 2012; Sun et al., 2017).结合新的高时间分辨率土壤呼吸数据集, 可以为土壤和根系呼吸的生物和环境控制机制提供新的见解(Hopkins et al., 2013).植物根系具有不同直径(粗跟和细根)和不同生理性状, 其呼吸速率模式及其对环境因子的响应具有明显差异.根系呼吸与根系直径、组织组成和特定根长等形态特征密切相关(Sun & Mao, 2011).通过根系呼吸与形态性状之间的相关关系, 可以建立区分粗根和细根呼吸的定量关系.细根对于植物生长、营养元素获取等生理功能具有重要作用, 其呼吸速率通常高于粗根, 且变异较大.在野外原位观测的结果发现, 根系直径与呼吸速率显著相关, 直径小于0.4 mm的根系呼吸速率显著高于其他根系, 且变异较大; 同时, 根系呼吸速率与根长和根的含氮量呈正相关关系(Makita et al., 2009). ...
Impacts of morphological traits and temperature on fine root respiration during dormancy of 1 2015
... 相对于土壤、植物叶片和茎干箱式通量观测系统, 根系箱式通量观测系统的难点在于适合于根系的气室箱体设计和实现(Marsden et al., 2008; Jia et al., 2013; Lai et al., 2015, 2016).传统根CO2通量观测通常将根系切断, 然后放到根呼吸气室箱体内, 但被切除的根系呼吸速率明显高于未被切除的根系(Makita et al., 2009, 2013; Sun et al., 2017).而通过原位根呼吸气室箱体设计可以很好地解决这一问题, 根呼吸箱通常使用PVC材料, 由两个半圆环或者一个封闭圆环(一端开口)构成, 将不同直径的根系放进根呼吸箱后, 形成封闭系统用于观测根呼吸CO2通量(Marsden et al., 2008; Kuptz et al., 2011a, 2011b).根系呼吸是森林土壤碳排放的重要来源, 其对土壤呼吸的贡献比例介于1/3到50%多之间(Marsden et al., 2008; Makita et al., 2009, 2012; Sun et al., 2017).结合新的高时间分辨率土壤呼吸数据集, 可以为土壤和根系呼吸的生物和环境控制机制提供新的见解(Hopkins et al., 2013).植物根系具有不同直径(粗跟和细根)和不同生理性状, 其呼吸速率模式及其对环境因子的响应具有明显差异.根系呼吸与根系直径、组织组成和特定根长等形态特征密切相关(Sun & Mao, 2011).通过根系呼吸与形态性状之间的相关关系, 可以建立区分粗根和细根呼吸的定量关系.细根对于植物生长、营养元素获取等生理功能具有重要作用, 其呼吸速率通常高于粗根, 且变异较大.在野外原位观测的结果发现, 根系直径与呼吸速率显著相关, 直径小于0.4 mm的根系呼吸速率显著高于其他根系, 且变异较大; 同时, 根系呼吸速率与根长和根的含氮量呈正相关关系(Makita et al., 2009). ...
底座入土深度和面积对典型草原土壤呼吸测定结果的影响 1 2019
... 测定过程中, 不能改变待测气体源汇的底物特征及环境条件, 影响待测气体的产生.土壤呼吸依赖于温度和湿度等环境因素, 人为的环境条件改变影响CO2通量(Davidson et al., 2002).测定过程应尽量减少对气室内温度和湿度等条件的影响.土壤环插入土壤的过程会切断土壤中的根系.通常在测定前尽量长的时间将土壤环安装到样地, 使得生态系统能够得到有效恢复; 在不影响系统整体性的前提下, 土壤环插入深度都尽量浅(Midwood & Millard, 2011; 李建军等, 2019), 但存在表层土壤空气流动的问题(Parkin et al., 2005).风不仅能引起气室内压力变化, 也会促进大气CO2与表层土壤CO2的混合(Susfalk et al., 2002; Millard et al., 2008).土壤环插入土壤深度过浅可能会存在大气CO2与表层土壤CO2的混合, 特别是对于干燥、多空隙的土壤来说, 会引入CO2浓度测定误差, 并显著改变测定的δ13C值(Susfalk et al., 2002; Millard et al., 2008).Hutchinson和Livingston (2001)的研究表明, 干燥、多气孔或者粗颗粒的土壤需将土壤环插入土壤超过10 cm; 而细颗粒、湿润或紧实的土壤只需将土壤环插入土壤不超过5 cm.Nickerson和Risk (2009)利用同位素模型模拟了正确的土壤环插入深度对于阻止空气进入气室的重要性, 最差的情况下, 气室内CO2的δ13C富集可达到4‰. ...
底座入土深度和面积对典型草原土壤呼吸测定结果的影响 1 2019
... 测定过程中, 不能改变待测气体源汇的底物特征及环境条件, 影响待测气体的产生.土壤呼吸依赖于温度和湿度等环境因素, 人为的环境条件改变影响CO2通量(Davidson et al., 2002).测定过程应尽量减少对气室内温度和湿度等条件的影响.土壤环插入土壤的过程会切断土壤中的根系.通常在测定前尽量长的时间将土壤环安装到样地, 使得生态系统能够得到有效恢复; 在不影响系统整体性的前提下, 土壤环插入深度都尽量浅(Midwood & Millard, 2011; 李建军等, 2019), 但存在表层土壤空气流动的问题(Parkin et al., 2005).风不仅能引起气室内压力变化, 也会促进大气CO2与表层土壤CO2的混合(Susfalk et al., 2002; Millard et al., 2008).土壤环插入土壤深度过浅可能会存在大气CO2与表层土壤CO2的混合, 特别是对于干燥、多空隙的土壤来说, 会引入CO2浓度测定误差, 并显著改变测定的δ13C值(Susfalk et al., 2002; Millard et al., 2008).Hutchinson和Livingston (2001)的研究表明, 干燥、多气孔或者粗颗粒的土壤需将土壤环插入土壤超过10 cm; 而细颗粒、湿润或紧实的土壤只需将土壤环插入土壤不超过5 cm.Nickerson和Risk (2009)利用同位素模型模拟了正确的土壤环插入深度对于阻止空气进入气室的重要性, 最差的情况下, 气室内CO2的δ13C富集可达到4‰. ...
Widespread asymmetric response of soil heterotrophic respiration to warming and cooling 1 2018
... 土壤CO2及其δ13C通量的原位长期连续观测, 有助于解析土壤呼吸随植物生理和环境等因素变化过程中的微小波动(Midwood et al., 2008; Bowling et al., 2015).土壤呼吸包括自养呼吸和异养呼吸, 而δ13C已经被证明可以用于拆分这两种组分(Midwood et al., 2008; Albanito et al., 2012).Midwood等(2008)利用土壤CO2稳态系统与稳定同位素质谱仪(Isotope Ratio Mass Spectrometry, IRMS)连用测定, 通过两元混合模型计算表明大麦(Hordeum vulgare)幼苗根中50%的碳可能来自于光合作用新合成的碳水化合物.Albanito等(2012)通过设计的参考室和双气室的稳态系统, 利用三元混合模型计算出土壤有机质、凋落物和根对土壤呼吸的相对贡献比例分别为37%、33%和30%.Liu等(2018)利用非稳态系统进行周期性升温和降温(5-30 ℃)培养试验, 结果表明土壤微生物呼吸对升温和降温具有明显的非对称响应, 升温阶段土壤微生物呼吸的增长速度更快. ...
Trace gas emission in chambers: a non-steady-state diffusion model 2 2006
... C显著偏负, 幅度最高可达4.4‰ (Livingston et al., 2006). ...
Carbon dioxide evolution of soil and crop growth 1 1927
... 由于原理简单且操作方便, 箱式通量观测技术和方法在自然或控制条件下的土壤、植物乃至生态系统尺度温室气体通量观测研究中得到广泛应用(Kuptz et al., 2011a, 2011b; He et al., 2013; Merbold et al., 2013; Savage et al., 2014).从早期的静态箱-碱液吸收滴定(Lundegardh, 1927)和静态箱-气相色谱测定(Midwood & Millard, 2011; Imer et al., 2013)等人工离线测定模式, 发展到目前通常采用的自动箱-红外光谱测定(Plain et al., 2009; Dubbert et al., 2014; Savage et al., 2014)等自动在线测定模式.但是, 通常缺乏对观测系统设计的理论要求和假设等问题的综合考虑和评价.首先, 如何保证系统测定数据的精度和准确度, 即保证系统气密性及气体混合、箱体内外气体浓度和压力保持一致等(Xu et al., 2006; Midwood et al., 2008); 其次, 如何保证系统测定数据的代表性, 即待测气体的产生和传输不受影响等(Midwood et al., 2008; Weiss et al., 2009; Snell et al., 2014). ...
Using the gradient method to determine soil gas flux: a review 2 2014
... 根据物质守恒方程, 要求待测气体传输只受扩散过程影响, 且全部扩散至气室箱体内, 同时测定过程不能影响CO2产生和传输过程, 否则测定的CO2释放量并不能代表呼吸通量(Midwood et al., 2008; Powers et al., 2010; Kammer et al., 2011; Midwood & Millard, 2011; Sturm et al., 2013; Savage et al., 2014).首先, 应该保证测定过程不影响待测气体的产生和传输, 即不改变温湿度等环境条件以及待测气体源汇底物特征, 并且有效消除气室箱体的边缘效应, 以及不影响气体的扩散和非扩散过程.其次, 客观验证和讨论待测气体传输只受扩散过程影响且全部扩散至气室箱体内的科学假设是否成立.例如, 土壤呼吸产生的CO2主要包括地表释放的CO2和土壤中储存的CO2两部分(Hanson & Gunderson, 2009; Midwood & Millard, 2011).通常假设土壤中储存的CO2通量可以忽略不计, 则箱式通量观测技术和方法(箱式法)直接测定的“土壤CO2释放量”等于“土壤呼吸” (Maier & Schack-Kirchner, 2014).如果土壤CO2在土壤中发生明显的储存(在某些情况下浓度高达数万μmol·mol-1), 那么就会显著影响箱式法的观测结果, 导致数据不具有代表性(Wang et al., 2018a, 2018b).土壤箱式通量法可与土壤通量梯度法相结合, 实现土壤CO2释放量的相互验证以及土壤CO2储存通量的估算(Maier et al., 2011; Maier & Schack-Kirchner, 2014). ...
... ; Maier & Schack-Kirchner, 2014). ...
Soil CO2 efflux vs. soil respiration: implications for flux models 1 2011
... 根据物质守恒方程, 要求待测气体传输只受扩散过程影响, 且全部扩散至气室箱体内, 同时测定过程不能影响CO2产生和传输过程, 否则测定的CO2释放量并不能代表呼吸通量(Midwood et al., 2008; Powers et al., 2010; Kammer et al., 2011; Midwood & Millard, 2011; Sturm et al., 2013; Savage et al., 2014).首先, 应该保证测定过程不影响待测气体的产生和传输, 即不改变温湿度等环境条件以及待测气体源汇底物特征, 并且有效消除气室箱体的边缘效应, 以及不影响气体的扩散和非扩散过程.其次, 客观验证和讨论待测气体传输只受扩散过程影响且全部扩散至气室箱体内的科学假设是否成立.例如, 土壤呼吸产生的CO2主要包括地表释放的CO2和土壤中储存的CO2两部分(Hanson & Gunderson, 2009; Midwood & Millard, 2011).通常假设土壤中储存的CO2通量可以忽略不计, 则箱式通量观测技术和方法(箱式法)直接测定的“土壤CO2释放量”等于“土壤呼吸” (Maier & Schack-Kirchner, 2014).如果土壤CO2在土壤中发生明显的储存(在某些情况下浓度高达数万μmol·mol-1), 那么就会显著影响箱式法的观测结果, 导致数据不具有代表性(Wang et al., 2018a, 2018b).土壤箱式通量法可与土壤通量梯度法相结合, 实现土壤CO2释放量的相互验证以及土壤CO2储存通量的估算(Maier et al., 2011; Maier & Schack-Kirchner, 2014). ...
Fine root morphological traits determine variation in root respiration of 3 2009
... 相对于土壤、植物叶片和茎干箱式通量观测系统, 根系箱式通量观测系统的难点在于适合于根系的气室箱体设计和实现(Marsden et al., 2008; Jia et al., 2013; Lai et al., 2015, 2016).传统根CO2通量观测通常将根系切断, 然后放到根呼吸气室箱体内, 但被切除的根系呼吸速率明显高于未被切除的根系(Makita et al., 2009, 2013; Sun et al., 2017).而通过原位根呼吸气室箱体设计可以很好地解决这一问题, 根呼吸箱通常使用PVC材料, 由两个半圆环或者一个封闭圆环(一端开口)构成, 将不同直径的根系放进根呼吸箱后, 形成封闭系统用于观测根呼吸CO2通量(Marsden et al., 2008; Kuptz et al., 2011a, 2011b).根系呼吸是森林土壤碳排放的重要来源, 其对土壤呼吸的贡献比例介于1/3到50%多之间(Marsden et al., 2008; Makita et al., 2009, 2012; Sun et al., 2017).结合新的高时间分辨率土壤呼吸数据集, 可以为土壤和根系呼吸的生物和环境控制机制提供新的见解(Hopkins et al., 2013).植物根系具有不同直径(粗跟和细根)和不同生理性状, 其呼吸速率模式及其对环境因子的响应具有明显差异.根系呼吸与根系直径、组织组成和特定根长等形态特征密切相关(Sun & Mao, 2011).通过根系呼吸与形态性状之间的相关关系, 可以建立区分粗根和细根呼吸的定量关系.细根对于植物生长、营养元素获取等生理功能具有重要作用, 其呼吸速率通常高于粗根, 且变异较大.在野外原位观测的结果发现, 根系直径与呼吸速率显著相关, 直径小于0.4 mm的根系呼吸速率显著高于其他根系, 且变异较大; 同时, 根系呼吸速率与根长和根的含氮量呈正相关关系(Makita et al., 2009). ...
... ; Makita et al., 2009, 2012; Sun et al., 2017).结合新的高时间分辨率土壤呼吸数据集, 可以为土壤和根系呼吸的生物和环境控制机制提供新的见解(Hopkins et al., 2013).植物根系具有不同直径(粗跟和细根)和不同生理性状, 其呼吸速率模式及其对环境因子的响应具有明显差异.根系呼吸与根系直径、组织组成和特定根长等形态特征密切相关(Sun & Mao, 2011).通过根系呼吸与形态性状之间的相关关系, 可以建立区分粗根和细根呼吸的定量关系.细根对于植物生长、营养元素获取等生理功能具有重要作用, 其呼吸速率通常高于粗根, 且变异较大.在野外原位观测的结果发现, 根系直径与呼吸速率显著相关, 直径小于0.4 mm的根系呼吸速率显著高于其他根系, 且变异较大; 同时, 根系呼吸速率与根长和根的含氮量呈正相关关系(Makita et al., 2009). ...
Patterns of root respiration rates and morphological traits in 13 tree species in a tropical forest 2012
Effects of excising and washing treatments on the root respiration rates of Japanese cedar (Cryptomeria japonica) seedlings 1 2013
... 相对于土壤、植物叶片和茎干箱式通量观测系统, 根系箱式通量观测系统的难点在于适合于根系的气室箱体设计和实现(Marsden et al., 2008; Jia et al., 2013; Lai et al., 2015, 2016).传统根CO2通量观测通常将根系切断, 然后放到根呼吸气室箱体内, 但被切除的根系呼吸速率明显高于未被切除的根系(Makita et al., 2009, 2013; Sun et al., 2017).而通过原位根呼吸气室箱体设计可以很好地解决这一问题, 根呼吸箱通常使用PVC材料, 由两个半圆环或者一个封闭圆环(一端开口)构成, 将不同直径的根系放进根呼吸箱后, 形成封闭系统用于观测根呼吸CO2通量(Marsden et al., 2008; Kuptz et al., 2011a, 2011b).根系呼吸是森林土壤碳排放的重要来源, 其对土壤呼吸的贡献比例介于1/3到50%多之间(Marsden et al., 2008; Makita et al., 2009, 2012; Sun et al., 2017).结合新的高时间分辨率土壤呼吸数据集, 可以为土壤和根系呼吸的生物和环境控制机制提供新的见解(Hopkins et al., 2013).植物根系具有不同直径(粗跟和细根)和不同生理性状, 其呼吸速率模式及其对环境因子的响应具有明显差异.根系呼吸与根系直径、组织组成和特定根长等形态特征密切相关(Sun & Mao, 2011).通过根系呼吸与形态性状之间的相关关系, 可以建立区分粗根和细根呼吸的定量关系.细根对于植物生长、营养元素获取等生理功能具有重要作用, 其呼吸速率通常高于粗根, 且变异较大.在野外原位观测的结果发现, 根系直径与呼吸速率显著相关, 直径小于0.4 mm的根系呼吸速率显著高于其他根系, 且变异较大; 同时, 根系呼吸速率与根长和根的含氮量呈正相关关系(Makita et al., 2009). ...
Seasonal and daily time course of the 13C composition in soil CO2 efflux recorded with a tunable diode laser spectrophotometer (TDLS) 4 2009
... 稳态箱式通量观测系统是通过使箱体内气体形成开放气路, 保持箱体内外CO2浓度一致或仅略高于大气, 通过测定箱体进气口和出气口CO2浓度差计算通量, 如图1C和1D所示.根据是否增加CO2浓度调节装置, 通常可以分为两类: (1)不增加CO2浓度调节装置, 仅通过将气泵和电磁阀切换实现利用气体分析仪测定进气口(参考气)和出气口(样品气) CO2浓度并计算差值.在测定过程中, 箱体内总CO2浓度略高于大气, 同时12CO2和13CO2浓度均略高于大气(Bahn et al., 2009; Plain et al., 2009; Subke et al., 2009).(2)增加CO2浓度调节装置, 通过向箱体内注入无CO2的空气, 保持箱体进气口和出气口气体流速相同, 使箱体内总CO2浓度始终与箱体外总CO2浓度基本保持一致, 仅略高于大气, 但设计和操作较难(Midwood et al., 2006; Marron et al., 2009).稳态系统通常没有明显的CO2累积(一般低于100 μmol·mol-1)(Midwood & Milllard, 2011), 因此, 测定过程中通常不会对CO2扩散产生明显影响, 这使得稳态系统在测定δ13C时比非稳态系统更有优势(Bahn et al., 2009; Plain et al., 2009; Subke et al., 2009). ...
... 与非稳态系统类似, 稳态系统CO2通量计算同样基于质量守恒方程的基本原理, 测定箱体进气口和出气口的CO2浓度、气体流速及箱体的底面积和高度(Marron et al., 2009; Powers et al., 2010).稳态系统CO2 δ13C通量计算也是根据同位素质量守恒的基本原理, 需要精确测定进气口和出气口的CO2浓度及δ13C值(Marron et al., 2009; Powers et al., 2010).稳态系统比非稳态系统更复杂, 特别在排放通量低的情况下, 进气口和出气口的差异明显减小, 进气口和出气口CO2浓度和δ13C的测定误差最终都会影响稳态系统CO2 δ13C通量的精度(Midwood et al., 2008). ...
... C值(Marron et al., 2009; Powers et al., 2010).稳态系统比非稳态系统更复杂, 特别在排放通量低的情况下, 进气口和出气口的差异明显减小, 进气口和出气口CO2浓度和δ13C的测定误差最终都会影响稳态系统CO2 δ13C通量的精度(Midwood et al., 2008). ...
... 稳态系统满足上述假设的方法通常也包括两种.首先, 通过增加CO2浓度调节装置向箱体内注入无CO2的空气, 保持箱体进气口和出气口气体流速相同, 使箱体内总CO2浓度始终与箱体外总CO2浓度保持动态平衡(Midwood et al., 2008; Marron et al., 2009; Powers et al., 2010).其次, 通过参考室与双层气室系统设计达到箱体内总CO2浓度始终与箱体外总CO2浓度保持动态平衡(Albanito et al., 2012), 其中参考气室上表面为CO2自由渗透膜, 双层气室的上层气室相当于上述第一种途径的稳态气室, 中间利用CO2渗透膜隔开, 下层气室为非稳态气室; 参考气室和上层气室利用红外光谱技术测定, 当二者CO2浓度一致时, 利用稳定同位素红外光谱技术测定下层气室CO2 δ13C值. ...
Two independent estimations of stand-level root respiration on clonal Eucalyptus stands in Congo: up scaling of direct measurements on roots versus the trenched-plot technique 3 2008
... 相对于土壤、植物叶片和茎干箱式通量观测系统, 根系箱式通量观测系统的难点在于适合于根系的气室箱体设计和实现(Marsden et al., 2008; Jia et al., 2013; Lai et al., 2015, 2016).传统根CO2通量观测通常将根系切断, 然后放到根呼吸气室箱体内, 但被切除的根系呼吸速率明显高于未被切除的根系(Makita et al., 2009, 2013; Sun et al., 2017).而通过原位根呼吸气室箱体设计可以很好地解决这一问题, 根呼吸箱通常使用PVC材料, 由两个半圆环或者一个封闭圆环(一端开口)构成, 将不同直径的根系放进根呼吸箱后, 形成封闭系统用于观测根呼吸CO2通量(Marsden et al., 2008; Kuptz et al., 2011a, 2011b).根系呼吸是森林土壤碳排放的重要来源, 其对土壤呼吸的贡献比例介于1/3到50%多之间(Marsden et al., 2008; Makita et al., 2009, 2012; Sun et al., 2017).结合新的高时间分辨率土壤呼吸数据集, 可以为土壤和根系呼吸的生物和环境控制机制提供新的见解(Hopkins et al., 2013).植物根系具有不同直径(粗跟和细根)和不同生理性状, 其呼吸速率模式及其对环境因子的响应具有明显差异.根系呼吸与根系直径、组织组成和特定根长等形态特征密切相关(Sun & Mao, 2011).通过根系呼吸与形态性状之间的相关关系, 可以建立区分粗根和细根呼吸的定量关系.细根对于植物生长、营养元素获取等生理功能具有重要作用, 其呼吸速率通常高于粗根, 且变异较大.在野外原位观测的结果发现, 根系直径与呼吸速率显著相关, 直径小于0.4 mm的根系呼吸速率显著高于其他根系, 且变异较大; 同时, 根系呼吸速率与根长和根的含氮量呈正相关关系(Makita et al., 2009). ...
... 通量(Marsden et al., 2008; Kuptz et al., 2011a, 2011b).根系呼吸是森林土壤碳排放的重要来源, 其对土壤呼吸的贡献比例介于1/3到50%多之间(Marsden et al., 2008; Makita et al., 2009, 2012; Sun et al., 2017).结合新的高时间分辨率土壤呼吸数据集, 可以为土壤和根系呼吸的生物和环境控制机制提供新的见解(Hopkins et al., 2013).植物根系具有不同直径(粗跟和细根)和不同生理性状, 其呼吸速率模式及其对环境因子的响应具有明显差异.根系呼吸与根系直径、组织组成和特定根长等形态特征密切相关(Sun & Mao, 2011).通过根系呼吸与形态性状之间的相关关系, 可以建立区分粗根和细根呼吸的定量关系.细根对于植物生长、营养元素获取等生理功能具有重要作用, 其呼吸速率通常高于粗根, 且变异较大.在野外原位观测的结果发现, 根系直径与呼吸速率显著相关, 直径小于0.4 mm的根系呼吸速率显著高于其他根系, 且变异较大; 同时, 根系呼吸速率与根长和根的含氮量呈正相关关系(Makita et al., 2009). ...
... ).根系呼吸是森林土壤碳排放的重要来源, 其对土壤呼吸的贡献比例介于1/3到50%多之间(Marsden et al., 2008; Makita et al., 2009, 2012; Sun et al., 2017).结合新的高时间分辨率土壤呼吸数据集, 可以为土壤和根系呼吸的生物和环境控制机制提供新的见解(Hopkins et al., 2013).植物根系具有不同直径(粗跟和细根)和不同生理性状, 其呼吸速率模式及其对环境因子的响应具有明显差异.根系呼吸与根系直径、组织组成和特定根长等形态特征密切相关(Sun & Mao, 2011).通过根系呼吸与形态性状之间的相关关系, 可以建立区分粗根和细根呼吸的定量关系.细根对于植物生长、营养元素获取等生理功能具有重要作用, 其呼吸速率通常高于粗根, 且变异较大.在野外原位观测的结果发现, 根系直径与呼吸速率显著相关, 直径小于0.4 mm的根系呼吸速率显著高于其他根系, 且变异较大; 同时, 根系呼吸速率与根长和根的含氮量呈正相关关系(Makita et al., 2009). ...
Permanent storage of carbon dioxide in geological reservoirs by mineral carbonation 1 2009
Winter greenhouse gas emissions (CO2, CH4 and N2O) from a sub-alpine grassland 1 2013
... 由于原理简单且操作方便, 箱式通量观测技术和方法在自然或控制条件下的土壤、植物乃至生态系统尺度温室气体通量观测研究中得到广泛应用(Kuptz et al., 2011a, 2011b; He et al., 2013; Merbold et al., 2013; Savage et al., 2014).从早期的静态箱-碱液吸收滴定(Lundegardh, 1927)和静态箱-气相色谱测定(Midwood & Millard, 2011; Imer et al., 2013)等人工离线测定模式, 发展到目前通常采用的自动箱-红外光谱测定(Plain et al., 2009; Dubbert et al., 2014; Savage et al., 2014)等自动在线测定模式.但是, 通常缺乏对观测系统设计的理论要求和假设等问题的综合考虑和评价.首先, 如何保证系统测定数据的精度和准确度, 即保证系统气密性及气体混合、箱体内外气体浓度和压力保持一致等(Xu et al., 2006; Midwood et al., 2008); 其次, 如何保证系统测定数据的代表性, 即待测气体的产生和传输不受影响等(Midwood et al., 2008; Weiss et al., 2009; Snell et al., 2014). ...
Collection and storage of CO2 for 13C analysis: an application to separate soil CO2 efflux into root- and soil-derived components 1 2006
... 稳态箱式通量观测系统是通过使箱体内气体形成开放气路, 保持箱体内外CO2浓度一致或仅略高于大气, 通过测定箱体进气口和出气口CO2浓度差计算通量, 如图1C和1D所示.根据是否增加CO2浓度调节装置, 通常可以分为两类: (1)不增加CO2浓度调节装置, 仅通过将气泵和电磁阀切换实现利用气体分析仪测定进气口(参考气)和出气口(样品气) CO2浓度并计算差值.在测定过程中, 箱体内总CO2浓度略高于大气, 同时12CO2和13CO2浓度均略高于大气(Bahn et al., 2009; Plain et al., 2009; Subke et al., 2009).(2)增加CO2浓度调节装置, 通过向箱体内注入无CO2的空气, 保持箱体进气口和出气口气体流速相同, 使箱体内总CO2浓度始终与箱体外总CO2浓度基本保持一致, 仅略高于大气, 但设计和操作较难(Midwood et al., 2006; Marron et al., 2009).稳态系统通常没有明显的CO2累积(一般低于100 μmol·mol-1)(Midwood & Milllard, 2011), 因此, 测定过程中通常不会对CO2扩散产生明显影响, 这使得稳态系统在测定δ13C时比非稳态系统更有优势(Bahn et al., 2009; Plain et al., 2009; Subke et al., 2009). ...
Challenges in measuring the δ13C of the soil surface CO2 efflux 10 2011
... 由于原理简单且操作方便, 箱式通量观测技术和方法在自然或控制条件下的土壤、植物乃至生态系统尺度温室气体通量观测研究中得到广泛应用(Kuptz et al., 2011a, 2011b; He et al., 2013; Merbold et al., 2013; Savage et al., 2014).从早期的静态箱-碱液吸收滴定(Lundegardh, 1927)和静态箱-气相色谱测定(Midwood & Millard, 2011; Imer et al., 2013)等人工离线测定模式, 发展到目前通常采用的自动箱-红外光谱测定(Plain et al., 2009; Dubbert et al., 2014; Savage et al., 2014)等自动在线测定模式.但是, 通常缺乏对观测系统设计的理论要求和假设等问题的综合考虑和评价.首先, 如何保证系统测定数据的精度和准确度, 即保证系统气密性及气体混合、箱体内外气体浓度和压力保持一致等(Xu et al., 2006; Midwood et al., 2008); 其次, 如何保证系统测定数据的代表性, 即待测气体的产生和传输不受影响等(Midwood et al., 2008; Weiss et al., 2009; Snell et al., 2014). ...
... 测定过程中, 不能改变待测气体源汇的底物特征及环境条件, 影响待测气体的产生.土壤呼吸依赖于温度和湿度等环境因素, 人为的环境条件改变影响CO2通量(Davidson et al., 2002).测定过程应尽量减少对气室内温度和湿度等条件的影响.土壤环插入土壤的过程会切断土壤中的根系.通常在测定前尽量长的时间将土壤环安装到样地, 使得生态系统能够得到有效恢复; 在不影响系统整体性的前提下, 土壤环插入深度都尽量浅(Midwood & Millard, 2011; 李建军等, 2019), 但存在表层土壤空气流动的问题(Parkin et al., 2005).风不仅能引起气室内压力变化, 也会促进大气CO2与表层土壤CO2的混合(Susfalk et al., 2002; Millard et al., 2008).土壤环插入土壤深度过浅可能会存在大气CO2与表层土壤CO2的混合, 特别是对于干燥、多空隙的土壤来说, 会引入CO2浓度测定误差, 并显著改变测定的δ13C值(Susfalk et al., 2002; Millard et al., 2008).Hutchinson和Livingston (2001)的研究表明, 干燥、多气孔或者粗颗粒的土壤需将土壤环插入土壤超过10 cm; 而细颗粒、湿润或紧实的土壤只需将土壤环插入土壤不超过5 cm.Nickerson和Risk (2009)利用同位素模型模拟了正确的土壤环插入深度对于阻止空气进入气室的重要性, 最差的情况下, 气室内CO2的δ13C富集可达到4‰. ...
... 所有箱式通量观测系统均需要对气体分析仪进行定期标定.目前常用的CO2及其δ13C气体分析仪主要基于稳定同位素红外光谱技术(Midwood & Millard, 2011; Griffis, 2013; Wen et al., 2013; Pang et al., 2016a), 但该技术自身也存在缺点, 主要表现为δ13C测量值对CO2浓度变化的非线性响应(浓度依赖性)以及对环境条件变化的敏感性导致的漂移(时间漂移).二者均对气体分析仪的精度和准确度产生影响, 需要进行适当校正和数据溯源以保证数据的可靠性(Kammer et al., 2011; Pang et al., 2016b; Chen et al., 2017, 2019).庞家平和温学发(2018)推荐利用3个或3个以上已知CO2浓度和δ13C真值的CO2标准气体涵盖待测气体CO2浓度, 进行气体分析仪的浓度依赖性校正, 同时通过设置适当的校正频率, 校正时间漂移并进行数据溯源. ...
... 根据物质守恒方程, 要求待测气体传输只受扩散过程影响, 且全部扩散至气室箱体内, 同时测定过程不能影响CO2产生和传输过程, 否则测定的CO2释放量并不能代表呼吸通量(Midwood et al., 2008; Powers et al., 2010; Kammer et al., 2011; Midwood & Millard, 2011; Sturm et al., 2013; Savage et al., 2014).首先, 应该保证测定过程不影响待测气体的产生和传输, 即不改变温湿度等环境条件以及待测气体源汇底物特征, 并且有效消除气室箱体的边缘效应, 以及不影响气体的扩散和非扩散过程.其次, 客观验证和讨论待测气体传输只受扩散过程影响且全部扩散至气室箱体内的科学假设是否成立.例如, 土壤呼吸产生的CO2主要包括地表释放的CO2和土壤中储存的CO2两部分(Hanson & Gunderson, 2009; Midwood & Millard, 2011).通常假设土壤中储存的CO2通量可以忽略不计, 则箱式通量观测技术和方法(箱式法)直接测定的“土壤CO2释放量”等于“土壤呼吸” (Maier & Schack-Kirchner, 2014).如果土壤CO2在土壤中发生明显的储存(在某些情况下浓度高达数万μmol·mol-1), 那么就会显著影响箱式法的观测结果, 导致数据不具有代表性(Wang et al., 2018a, 2018b).土壤箱式通量法可与土壤通量梯度法相结合, 实现土壤CO2释放量的相互验证以及土壤CO2储存通量的估算(Maier et al., 2011; Maier & Schack-Kirchner, 2014). ...
Measuring the 13C content of soil-respired CO2 using a novel open chamber system 11 2008
... 由于原理简单且操作方便, 箱式通量观测技术和方法在自然或控制条件下的土壤、植物乃至生态系统尺度温室气体通量观测研究中得到广泛应用(Kuptz et al., 2011a, 2011b; He et al., 2013; Merbold et al., 2013; Savage et al., 2014).从早期的静态箱-碱液吸收滴定(Lundegardh, 1927)和静态箱-气相色谱测定(Midwood & Millard, 2011; Imer et al., 2013)等人工离线测定模式, 发展到目前通常采用的自动箱-红外光谱测定(Plain et al., 2009; Dubbert et al., 2014; Savage et al., 2014)等自动在线测定模式.但是, 通常缺乏对观测系统设计的理论要求和假设等问题的综合考虑和评价.首先, 如何保证系统测定数据的精度和准确度, 即保证系统气密性及气体混合、箱体内外气体浓度和压力保持一致等(Xu et al., 2006; Midwood et al., 2008); 其次, 如何保证系统测定数据的代表性, 即待测气体的产生和传输不受影响等(Midwood et al., 2008; Weiss et al., 2009; Snell et al., 2014). ...
... ); 其次, 如何保证系统测定数据的代表性, 即待测气体的产生和传输不受影响等(Midwood et al., 2008; Weiss et al., 2009; Snell et al., 2014). ...
... 与非稳态系统类似, 稳态系统CO2通量计算同样基于质量守恒方程的基本原理, 测定箱体进气口和出气口的CO2浓度、气体流速及箱体的底面积和高度(Marron et al., 2009; Powers et al., 2010).稳态系统CO2 δ13C通量计算也是根据同位素质量守恒的基本原理, 需要精确测定进气口和出气口的CO2浓度及δ13C值(Marron et al., 2009; Powers et al., 2010).稳态系统比非稳态系统更复杂, 特别在排放通量低的情况下, 进气口和出气口的差异明显减小, 进气口和出气口CO2浓度和δ13C的测定误差最终都会影响稳态系统CO2 δ13C通量的精度(Midwood et al., 2008). ...
... 箱式通量观测系统需要保证气体分析仪以及系统气路的气密性, 即不存在与外界大气的气体交换过程.测定过程中, 需要箱体内以及系统内气体充分混合才能代表待测气体.通常采取在气室箱体顶端安装风扇方式进行气体混匀(Midwood et al., 2008).但是由于风扇可能导致气室内压力波动, 影响CO2扩散梯度, 进而导致CO2排放通量的高估或低估(Midwood et al., 2008).通常仅通过气室箱体与气体分析仪间的气体循环过程实现系统内气体充分混合(Pumpanen et al., 2004; Kammer et al., 2011).需要注意的是, 开始测量时, 系统内均会残留上一个测定的气体, 需要短暂的系统平衡时间(图1B、1D), 才能实现系统内的气体充分混合.需要保证不同气室箱体间的有效切换才能代表待测气体, 即需要剔除气路切换后的无效数据后保证足够的有效数据量.无效数据量所对应的时间越短代表切换效率越高.同时, 对于CO2 δ13C通量测定, 需要选用不会造成同位素分馏效应的材料构建系统. ...
... 排放通量的高估或低估(Midwood et al., 2008).通常仅通过气室箱体与气体分析仪间的气体循环过程实现系统内气体充分混合(Pumpanen et al., 2004; Kammer et al., 2011).需要注意的是, 开始测量时, 系统内均会残留上一个测定的气体, 需要短暂的系统平衡时间(图1B、1D), 才能实现系统内的气体充分混合.需要保证不同气室箱体间的有效切换才能代表待测气体, 即需要剔除气路切换后的无效数据后保证足够的有效数据量.无效数据量所对应的时间越短代表切换效率越高.同时, 对于CO2 δ13C通量测定, 需要选用不会造成同位素分馏效应的材料构建系统. ...
... 稳态系统满足上述假设的方法通常也包括两种.首先, 通过增加CO2浓度调节装置向箱体内注入无CO2的空气, 保持箱体进气口和出气口气体流速相同, 使箱体内总CO2浓度始终与箱体外总CO2浓度保持动态平衡(Midwood et al., 2008; Marron et al., 2009; Powers et al., 2010).其次, 通过参考室与双层气室系统设计达到箱体内总CO2浓度始终与箱体外总CO2浓度保持动态平衡(Albanito et al., 2012), 其中参考气室上表面为CO2自由渗透膜, 双层气室的上层气室相当于上述第一种途径的稳态气室, 中间利用CO2渗透膜隔开, 下层气室为非稳态气室; 参考气室和上层气室利用红外光谱技术测定, 当二者CO2浓度一致时, 利用稳定同位素红外光谱技术测定下层气室CO2 δ13C值. ...
... 土壤CO2及其δ13C通量的原位长期连续观测, 有助于解析土壤呼吸随植物生理和环境等因素变化过程中的微小波动(Midwood et al., 2008; Bowling et al., 2015).土壤呼吸包括自养呼吸和异养呼吸, 而δ13C已经被证明可以用于拆分这两种组分(Midwood et al., 2008; Albanito et al., 2012).Midwood等(2008)利用土壤CO2稳态系统与稳定同位素质谱仪(Isotope Ratio Mass Spectrometry, IRMS)连用测定, 通过两元混合模型计算表明大麦(Hordeum vulgare)幼苗根中50%的碳可能来自于光合作用新合成的碳水化合物.Albanito等(2012)通过设计的参考室和双气室的稳态系统, 利用三元混合模型计算出土壤有机质、凋落物和根对土壤呼吸的相对贡献比例分别为37%、33%和30%.Liu等(2018)利用非稳态系统进行周期性升温和降温(5-30 ℃)培养试验, 结果表明土壤微生物呼吸对升温和降温具有明显的非对称响应, 升温阶段土壤微生物呼吸的增长速度更快. ...
... C已经被证明可以用于拆分这两种组分(Midwood et al., 2008; Albanito et al., 2012).Midwood等(2008)利用土壤CO2稳态系统与稳定同位素质谱仪(Isotope Ratio Mass Spectrometry, IRMS)连用测定, 通过两元混合模型计算表明大麦(Hordeum vulgare)幼苗根中50%的碳可能来自于光合作用新合成的碳水化合物.Albanito等(2012)通过设计的参考室和双气室的稳态系统, 利用三元混合模型计算出土壤有机质、凋落物和根对土壤呼吸的相对贡献比例分别为37%、33%和30%.Liu等(2018)利用非稳态系统进行周期性升温和降温(5-30 ℃)培养试验, 结果表明土壤微生物呼吸对升温和降温具有明显的非对称响应, 升温阶段土壤微生物呼吸的增长速度更快. ...
... ).Midwood等(2008)利用土壤CO2稳态系统与稳定同位素质谱仪(Isotope Ratio Mass Spectrometry, IRMS)连用测定, 通过两元混合模型计算表明大麦(Hordeum vulgare)幼苗根中50%的碳可能来自于光合作用新合成的碳水化合物.Albanito等(2012)通过设计的参考室和双气室的稳态系统, 利用三元混合模型计算出土壤有机质、凋落物和根对土壤呼吸的相对贡献比例分别为37%、33%和30%.Liu等(2018)利用非稳态系统进行周期性升温和降温(5-30 ℃)培养试验, 结果表明土壤微生物呼吸对升温和降温具有明显的非对称响应, 升温阶段土壤微生物呼吸的增长速度更快. ...
... 所有箱式通量观测系统均需要对系统测定数据的精度和准确度进行评价.可以通过零通量和模拟通量测试对箱式通量观测系统测定数据的精度和准度进行评价(Midwood et al., 2008; Powers et al., 2010; 高峻等, 2011; 于贵瑞等, 2015).零通量测试与系统漏率测试类似, 但不需要使用标气, 只要将气室箱体密封进行正常测定即可, 然后计算CO2通量评价系统是否满足气密性的基本需求.模拟通量测试需要利用注射泵向模拟装置中匀速地注入标准气体来模拟土壤气体的排放过程, 将实际测定排放速率与模拟通量的设定值对比分析, 即可验证观测系统的准确度和精度. ...
... 根据物质守恒方程, 要求待测气体传输只受扩散过程影响, 且全部扩散至气室箱体内, 同时测定过程不能影响CO2产生和传输过程, 否则测定的CO2释放量并不能代表呼吸通量(Midwood et al., 2008; Powers et al., 2010; Kammer et al., 2011; Midwood & Millard, 2011; Sturm et al., 2013; Savage et al., 2014).首先, 应该保证测定过程不影响待测气体的产生和传输, 即不改变温湿度等环境条件以及待测气体源汇底物特征, 并且有效消除气室箱体的边缘效应, 以及不影响气体的扩散和非扩散过程.其次, 客观验证和讨论待测气体传输只受扩散过程影响且全部扩散至气室箱体内的科学假设是否成立.例如, 土壤呼吸产生的CO2主要包括地表释放的CO2和土壤中储存的CO2两部分(Hanson & Gunderson, 2009; Midwood & Millard, 2011).通常假设土壤中储存的CO2通量可以忽略不计, 则箱式通量观测技术和方法(箱式法)直接测定的“土壤CO2释放量”等于“土壤呼吸” (Maier & Schack-Kirchner, 2014).如果土壤CO2在土壤中发生明显的储存(在某些情况下浓度高达数万μmol·mol-1), 那么就会显著影响箱式法的观测结果, 导致数据不具有代表性(Wang et al., 2018a, 2018b).土壤箱式通量法可与土壤通量梯度法相结合, 实现土壤CO2释放量的相互验证以及土壤CO2储存通量的估算(Maier et al., 2011; Maier & Schack-Kirchner, 2014). ...
Partitioning soil surface CO2 efflux into autotrophic and heterotrophic components, using natural gradients in soil δ13C in an undisturbed savannah soil 2 2008
... 测定过程中, 不能改变待测气体源汇的底物特征及环境条件, 影响待测气体的产生.土壤呼吸依赖于温度和湿度等环境因素, 人为的环境条件改变影响CO2通量(Davidson et al., 2002).测定过程应尽量减少对气室内温度和湿度等条件的影响.土壤环插入土壤的过程会切断土壤中的根系.通常在测定前尽量长的时间将土壤环安装到样地, 使得生态系统能够得到有效恢复; 在不影响系统整体性的前提下, 土壤环插入深度都尽量浅(Midwood & Millard, 2011; 李建军等, 2019), 但存在表层土壤空气流动的问题(Parkin et al., 2005).风不仅能引起气室内压力变化, 也会促进大气CO2与表层土壤CO2的混合(Susfalk et al., 2002; Millard et al., 2008).土壤环插入土壤深度过浅可能会存在大气CO2与表层土壤CO2的混合, 特别是对于干燥、多空隙的土壤来说, 会引入CO2浓度测定误差, 并显著改变测定的δ13C值(Susfalk et al., 2002; Millard et al., 2008).Hutchinson和Livingston (2001)的研究表明, 干燥、多气孔或者粗颗粒的土壤需将土壤环插入土壤超过10 cm; 而细颗粒、湿润或紧实的土壤只需将土壤环插入土壤不超过5 cm.Nickerson和Risk (2009)利用同位素模型模拟了正确的土壤环插入深度对于阻止空气进入气室的重要性, 最差的情况下, 气室内CO2的δ13C富集可达到4‰. ...
... 所有箱式通量观测系统均需要对气体分析仪进行定期标定.目前常用的CO2及其δ13C气体分析仪主要基于稳定同位素红外光谱技术(Midwood & Millard, 2011; Griffis, 2013; Wen et al., 2013; Pang et al., 2016a), 但该技术自身也存在缺点, 主要表现为δ13C测量值对CO2浓度变化的非线性响应(浓度依赖性)以及对环境条件变化的敏感性导致的漂移(时间漂移).二者均对气体分析仪的精度和准确度产生影响, 需要进行适当校正和数据溯源以保证数据的可靠性(Kammer et al., 2011; Pang et al., 2016b; Chen et al., 2017, 2019).庞家平和温学发(2018)推荐利用3个或3个以上已知CO2浓度和δ13C真值的CO2标准气体涵盖待测气体CO2浓度, 进行气体分析仪的浓度依赖性校正, 同时通过设置适当的校正频率, 校正时间漂移并进行数据溯源. ...
稳定同位素红外光谱技术测定CO2同位素校正方法的研究进展 1 2018
... 所有箱式通量观测系统均需要对气体分析仪进行定期标定.目前常用的CO2及其δ13C气体分析仪主要基于稳定同位素红外光谱技术(Midwood & Millard, 2011; Griffis, 2013; Wen et al., 2013; Pang et al., 2016a), 但该技术自身也存在缺点, 主要表现为δ13C测量值对CO2浓度变化的非线性响应(浓度依赖性)以及对环境条件变化的敏感性导致的漂移(时间漂移).二者均对气体分析仪的精度和准确度产生影响, 需要进行适当校正和数据溯源以保证数据的可靠性(Kammer et al., 2011; Pang et al., 2016b; Chen et al., 2017, 2019).庞家平和温学发(2018)推荐利用3个或3个以上已知CO2浓度和δ13C真值的CO2标准气体涵盖待测气体CO2浓度, 进行气体分析仪的浓度依赖性校正, 同时通过设置适当的校正频率, 校正时间漂移并进行数据溯源. ...
Mixing ratio and carbon isotopic composition investigation of atmospheric CO2 in Beijing, China 1 2016a
... 所有箱式通量观测系统均需要对气体分析仪进行定期标定.目前常用的CO2及其δ13C气体分析仪主要基于稳定同位素红外光谱技术(Midwood & Millard, 2011; Griffis, 2013; Wen et al., 2013; Pang et al., 2016a), 但该技术自身也存在缺点, 主要表现为δ13C测量值对CO2浓度变化的非线性响应(浓度依赖性)以及对环境条件变化的敏感性导致的漂移(时间漂移).二者均对气体分析仪的精度和准确度产生影响, 需要进行适当校正和数据溯源以保证数据的可靠性(Kammer et al., 2011; Pang et al., 2016b; Chen et al., 2017, 2019).庞家平和温学发(2018)推荐利用3个或3个以上已知CO2浓度和δ13C真值的CO2标准气体涵盖待测气体CO2浓度, 进行气体分析仪的浓度依赖性校正, 同时通过设置适当的校正频率, 校正时间漂移并进行数据溯源. ...
Intercomparison of two cavity ring-down spectroscopy analyzers for atmospheric 13CO2/12CO2 measurement 1 2016b
... 所有箱式通量观测系统均需要对气体分析仪进行定期标定.目前常用的CO2及其δ13C气体分析仪主要基于稳定同位素红外光谱技术(Midwood & Millard, 2011; Griffis, 2013; Wen et al., 2013; Pang et al., 2016a), 但该技术自身也存在缺点, 主要表现为δ13C测量值对CO2浓度变化的非线性响应(浓度依赖性)以及对环境条件变化的敏感性导致的漂移(时间漂移).二者均对气体分析仪的精度和准确度产生影响, 需要进行适当校正和数据溯源以保证数据的可靠性(Kammer et al., 2011; Pang et al., 2016b; Chen et al., 2017, 2019).庞家平和温学发(2018)推荐利用3个或3个以上已知CO2浓度和δ13C真值的CO2标准气体涵盖待测气体CO2浓度, 进行气体分析仪的浓度依赖性校正, 同时通过设置适当的校正频率, 校正时间漂移并进行数据溯源. ...
Relationship of soil respiration to crop and landscape in the walnut creek watershed 1 2005
... 测定过程中, 不能改变待测气体源汇的底物特征及环境条件, 影响待测气体的产生.土壤呼吸依赖于温度和湿度等环境因素, 人为的环境条件改变影响CO2通量(Davidson et al., 2002).测定过程应尽量减少对气室内温度和湿度等条件的影响.土壤环插入土壤的过程会切断土壤中的根系.通常在测定前尽量长的时间将土壤环安装到样地, 使得生态系统能够得到有效恢复; 在不影响系统整体性的前提下, 土壤环插入深度都尽量浅(Midwood & Millard, 2011; 李建军等, 2019), 但存在表层土壤空气流动的问题(Parkin et al., 2005).风不仅能引起气室内压力变化, 也会促进大气CO2与表层土壤CO2的混合(Susfalk et al., 2002; Millard et al., 2008).土壤环插入土壤深度过浅可能会存在大气CO2与表层土壤CO2的混合, 特别是对于干燥、多空隙的土壤来说, 会引入CO2浓度测定误差, 并显著改变测定的δ13C值(Susfalk et al., 2002; Millard et al., 2008).Hutchinson和Livingston (2001)的研究表明, 干燥、多气孔或者粗颗粒的土壤需将土壤环插入土壤超过10 cm; 而细颗粒、湿润或紧实的土壤只需将土壤环插入土壤不超过5 cm.Nickerson和Risk (2009)利用同位素模型模拟了正确的土壤环插入深度对于阻止空气进入气室的重要性, 最差的情况下, 气室内CO2的δ13C富集可达到4‰. ...
Tracing of recently assimilated carbon in respiration at high temporal resolution in the field with a tuneable diode laser absorption spectrometer after in situ13CO2 pulse labelling of 20-year-old beech trees 5 2009
... 由于原理简单且操作方便, 箱式通量观测技术和方法在自然或控制条件下的土壤、植物乃至生态系统尺度温室气体通量观测研究中得到广泛应用(Kuptz et al., 2011a, 2011b; He et al., 2013; Merbold et al., 2013; Savage et al., 2014).从早期的静态箱-碱液吸收滴定(Lundegardh, 1927)和静态箱-气相色谱测定(Midwood & Millard, 2011; Imer et al., 2013)等人工离线测定模式, 发展到目前通常采用的自动箱-红外光谱测定(Plain et al., 2009; Dubbert et al., 2014; Savage et al., 2014)等自动在线测定模式.但是, 通常缺乏对观测系统设计的理论要求和假设等问题的综合考虑和评价.首先, 如何保证系统测定数据的精度和准确度, 即保证系统气密性及气体混合、箱体内外气体浓度和压力保持一致等(Xu et al., 2006; Midwood et al., 2008); 其次, 如何保证系统测定数据的代表性, 即待测气体的产生和传输不受影响等(Midwood et al., 2008; Weiss et al., 2009; Snell et al., 2014). ...
... 稳态箱式通量观测系统是通过使箱体内气体形成开放气路, 保持箱体内外CO2浓度一致或仅略高于大气, 通过测定箱体进气口和出气口CO2浓度差计算通量, 如图1C和1D所示.根据是否增加CO2浓度调节装置, 通常可以分为两类: (1)不增加CO2浓度调节装置, 仅通过将气泵和电磁阀切换实现利用气体分析仪测定进气口(参考气)和出气口(样品气) CO2浓度并计算差值.在测定过程中, 箱体内总CO2浓度略高于大气, 同时12CO2和13CO2浓度均略高于大气(Bahn et al., 2009; Plain et al., 2009; Subke et al., 2009).(2)增加CO2浓度调节装置, 通过向箱体内注入无CO2的空气, 保持箱体进气口和出气口气体流速相同, 使箱体内总CO2浓度始终与箱体外总CO2浓度基本保持一致, 仅略高于大气, 但设计和操作较难(Midwood et al., 2006; Marron et al., 2009).稳态系统通常没有明显的CO2累积(一般低于100 μmol·mol-1)(Midwood & Milllard, 2011), 因此, 测定过程中通常不会对CO2扩散产生明显影响, 这使得稳态系统在测定δ13C时比非稳态系统更有优势(Bahn et al., 2009; Plain et al., 2009; Subke et al., 2009). ...
... ; Plain et al., 2009; Subke et al., 2009). ...
... 生态系统箱式通量观测系统主要分为两种: 冠层通量测定(不包括土壤)和生态系统通量测定(包括土壤), 可以采用明箱或暗箱测定.生态系统通量观测多应用于草本或小灌木生态系统通量观测研究(Wu et al., 2010; Hafner et al., 2012; Wei et al., 2016).高大乔木根据研究目标可以选择进行冠层或生态系统CO2及其δ13C通量观测.但由于难度较高而选择进行13CO2脉冲标记研究(Plain et al., 2009; Epron et al., 2016).通常利用不同高度的钢制脚手架环绕树的冠层, 利用高透光率的聚合物膜或有机玻璃等制作冠层箱(明箱或暗箱), 箱体的底座由两个半圆形不锈钢板制成, 钢板中间开口, 以便容纳茎干, 并固定在脚手架上(Plain et al., 2009; Epron et al., 2016).为了维持箱体内的温湿度条件, 通常配套安装空调, 利用风扇混匀冠层箱内的气体(Epron et al., 2016).Epron等(2016)利用27或38 m3的高透光率膜罩住整个桉树(Eucalyptus robusta)冠层进行脉冲标记, 利用茎干和根CO2及其δ13C通量观测箱实时观测, 结果表明, 叶片中标记的碳的储存时间非常短(21-31 h), 其在茎干中的迁移速率为0.20-0.82 m·h-1. ...
... ).通常利用不同高度的钢制脚手架环绕树的冠层, 利用高透光率的聚合物膜或有机玻璃等制作冠层箱(明箱或暗箱), 箱体的底座由两个半圆形不锈钢板制成, 钢板中间开口, 以便容纳茎干, 并固定在脚手架上(Plain et al., 2009; Epron et al., 2016).为了维持箱体内的温湿度条件, 通常配套安装空调, 利用风扇混匀冠层箱内的气体(Epron et al., 2016).Epron等(2016)利用27或38 m3的高透光率膜罩住整个桉树(Eucalyptus robusta)冠层进行脉冲标记, 利用茎干和根CO2及其δ13C通量观测箱实时观测, 结果表明, 叶片中标记的碳的储存时间非常短(21-31 h), 其在茎干中的迁移速率为0.20-0.82 m·h-1. ...
A dynamic soil chamber system coupled with a tunable diode laser for online measurements of δ13C, δ18O, and efflux rate of soil-respired CO2 5 2010
... 与非稳态系统类似, 稳态系统CO2通量计算同样基于质量守恒方程的基本原理, 测定箱体进气口和出气口的CO2浓度、气体流速及箱体的底面积和高度(Marron et al., 2009; Powers et al., 2010).稳态系统CO2 δ13C通量计算也是根据同位素质量守恒的基本原理, 需要精确测定进气口和出气口的CO2浓度及δ13C值(Marron et al., 2009; Powers et al., 2010).稳态系统比非稳态系统更复杂, 特别在排放通量低的情况下, 进气口和出气口的差异明显减小, 进气口和出气口CO2浓度和δ13C的测定误差最终都会影响稳态系统CO2 δ13C通量的精度(Midwood et al., 2008). ...
... ; Powers et al., 2010).稳态系统比非稳态系统更复杂, 特别在排放通量低的情况下, 进气口和出气口的差异明显减小, 进气口和出气口CO2浓度和δ13C的测定误差最终都会影响稳态系统CO2 δ13C通量的精度(Midwood et al., 2008). ...
... 稳态系统满足上述假设的方法通常也包括两种.首先, 通过增加CO2浓度调节装置向箱体内注入无CO2的空气, 保持箱体进气口和出气口气体流速相同, 使箱体内总CO2浓度始终与箱体外总CO2浓度保持动态平衡(Midwood et al., 2008; Marron et al., 2009; Powers et al., 2010).其次, 通过参考室与双层气室系统设计达到箱体内总CO2浓度始终与箱体外总CO2浓度保持动态平衡(Albanito et al., 2012), 其中参考气室上表面为CO2自由渗透膜, 双层气室的上层气室相当于上述第一种途径的稳态气室, 中间利用CO2渗透膜隔开, 下层气室为非稳态气室; 参考气室和上层气室利用红外光谱技术测定, 当二者CO2浓度一致时, 利用稳定同位素红外光谱技术测定下层气室CO2 δ13C值. ...
... 所有箱式通量观测系统均需要对系统测定数据的精度和准确度进行评价.可以通过零通量和模拟通量测试对箱式通量观测系统测定数据的精度和准度进行评价(Midwood et al., 2008; Powers et al., 2010; 高峻等, 2011; 于贵瑞等, 2015).零通量测试与系统漏率测试类似, 但不需要使用标气, 只要将气室箱体密封进行正常测定即可, 然后计算CO2通量评价系统是否满足气密性的基本需求.模拟通量测试需要利用注射泵向模拟装置中匀速地注入标准气体来模拟土壤气体的排放过程, 将实际测定排放速率与模拟通量的设定值对比分析, 即可验证观测系统的准确度和精度. ...
... 根据物质守恒方程, 要求待测气体传输只受扩散过程影响, 且全部扩散至气室箱体内, 同时测定过程不能影响CO2产生和传输过程, 否则测定的CO2释放量并不能代表呼吸通量(Midwood et al., 2008; Powers et al., 2010; Kammer et al., 2011; Midwood & Millard, 2011; Sturm et al., 2013; Savage et al., 2014).首先, 应该保证测定过程不影响待测气体的产生和传输, 即不改变温湿度等环境条件以及待测气体源汇底物特征, 并且有效消除气室箱体的边缘效应, 以及不影响气体的扩散和非扩散过程.其次, 客观验证和讨论待测气体传输只受扩散过程影响且全部扩散至气室箱体内的科学假设是否成立.例如, 土壤呼吸产生的CO2主要包括地表释放的CO2和土壤中储存的CO2两部分(Hanson & Gunderson, 2009; Midwood & Millard, 2011).通常假设土壤中储存的CO2通量可以忽略不计, 则箱式通量观测技术和方法(箱式法)直接测定的“土壤CO2释放量”等于“土壤呼吸” (Maier & Schack-Kirchner, 2014).如果土壤CO2在土壤中发生明显的储存(在某些情况下浓度高达数万μmol·mol-1), 那么就会显著影响箱式法的观测结果, 导致数据不具有代表性(Wang et al., 2018a, 2018b).土壤箱式通量法可与土壤通量梯度法相结合, 实现土壤CO2释放量的相互验证以及土壤CO2储存通量的估算(Maier et al., 2011; Maier & Schack-Kirchner, 2014). ...
Comparison of different chamber techniques for measuring soil CO2 efflux 1 2004
... 箱式通量观测系统需要保证气体分析仪以及系统气路的气密性, 即不存在与外界大气的气体交换过程.测定过程中, 需要箱体内以及系统内气体充分混合才能代表待测气体.通常采取在气室箱体顶端安装风扇方式进行气体混匀(Midwood et al., 2008).但是由于风扇可能导致气室内压力波动, 影响CO2扩散梯度, 进而导致CO2排放通量的高估或低估(Midwood et al., 2008).通常仅通过气室箱体与气体分析仪间的气体循环过程实现系统内气体充分混合(Pumpanen et al., 2004; Kammer et al., 2011).需要注意的是, 开始测量时, 系统内均会残留上一个测定的气体, 需要短暂的系统平衡时间(图1B、1D), 才能实现系统内的气体充分混合.需要保证不同气室箱体间的有效切换才能代表待测气体, 即需要剔除气路切换后的无效数据后保证足够的有效数据量.无效数据量所对应的时间越短代表切换效率越高.同时, 对于CO2 δ13C通量测定, 需要选用不会造成同位素分馏效应的材料构建系统. ...
Mind the gap: non-biological processes contributing to soil CO2 efflux 2 2015
Temporal and spatial patterns of internal and external stem CO2 fluxes in a sub-Mediterranean oak 1 2016
... 茎干箱式通量观测系统通常可以直接使用或借鉴土壤箱式通量观测系统.根据胸径大小制作特定直径的PVC环, 密封固定在树干上直接利用土壤箱式通量观测系统进行观测(Yang et al., 2012; Hilman & Angert, 2016; Salomón et al., 2016), 或者制作适合茎干的特定箱体并与气体分析仪相连构成茎干箱式通量观测系统(Wang et al., 2019; Yang et al., 2019).茎干CO2通量是森林生态系统碳循环中的重要组成部分, 占全球森林生态系统净初级生产力的20% (Yang et al., 2016, 2019; Zhao et al., 2018).研究表明, 茎干呼吸的日变化趋势呈现单峰曲线、双峰曲线和S型曲线3种模式(石新立等, 2010).其中, 气候和植被类型等可能是不同变化趋势的决定因素.Yang等(2016)通过收集全球121篇已发表文章的数据, 研究了茎干呼吸与叶面积指数、年平均气温及年平均降水量的相关关系, 发现茎干呼吸与叶面积指数呈正相关关系, 叶面积指数、年平均气温及降水量可以解释茎干呼吸变异的74%. ...
High temporal frequency measurements of greenhouse gas emissions from soils 3 2014
... 由于原理简单且操作方便, 箱式通量观测技术和方法在自然或控制条件下的土壤、植物乃至生态系统尺度温室气体通量观测研究中得到广泛应用(Kuptz et al., 2011a, 2011b; He et al., 2013; Merbold et al., 2013; Savage et al., 2014).从早期的静态箱-碱液吸收滴定(Lundegardh, 1927)和静态箱-气相色谱测定(Midwood & Millard, 2011; Imer et al., 2013)等人工离线测定模式, 发展到目前通常采用的自动箱-红外光谱测定(Plain et al., 2009; Dubbert et al., 2014; Savage et al., 2014)等自动在线测定模式.但是, 通常缺乏对观测系统设计的理论要求和假设等问题的综合考虑和评价.首先, 如何保证系统测定数据的精度和准确度, 即保证系统气密性及气体混合、箱体内外气体浓度和压力保持一致等(Xu et al., 2006; Midwood et al., 2008); 其次, 如何保证系统测定数据的代表性, 即待测气体的产生和传输不受影响等(Midwood et al., 2008; Weiss et al., 2009; Snell et al., 2014). ...
... ; Savage et al., 2014)等自动在线测定模式.但是, 通常缺乏对观测系统设计的理论要求和假设等问题的综合考虑和评价.首先, 如何保证系统测定数据的精度和准确度, 即保证系统气密性及气体混合、箱体内外气体浓度和压力保持一致等(Xu et al., 2006; Midwood et al., 2008); 其次, 如何保证系统测定数据的代表性, 即待测气体的产生和传输不受影响等(Midwood et al., 2008; Weiss et al., 2009; Snell et al., 2014). ...
... 根据物质守恒方程, 要求待测气体传输只受扩散过程影响, 且全部扩散至气室箱体内, 同时测定过程不能影响CO2产生和传输过程, 否则测定的CO2释放量并不能代表呼吸通量(Midwood et al., 2008; Powers et al., 2010; Kammer et al., 2011; Midwood & Millard, 2011; Sturm et al., 2013; Savage et al., 2014).首先, 应该保证测定过程不影响待测气体的产生和传输, 即不改变温湿度等环境条件以及待测气体源汇底物特征, 并且有效消除气室箱体的边缘效应, 以及不影响气体的扩散和非扩散过程.其次, 客观验证和讨论待测气体传输只受扩散过程影响且全部扩散至气室箱体内的科学假设是否成立.例如, 土壤呼吸产生的CO2主要包括地表释放的CO2和土壤中储存的CO2两部分(Hanson & Gunderson, 2009; Midwood & Millard, 2011).通常假设土壤中储存的CO2通量可以忽略不计, 则箱式通量观测技术和方法(箱式法)直接测定的“土壤CO2释放量”等于“土壤呼吸” (Maier & Schack-Kirchner, 2014).如果土壤CO2在土壤中发生明显的储存(在某些情况下浓度高达数万μmol·mol-1), 那么就会显著影响箱式法的观测结果, 导致数据不具有代表性(Wang et al., 2018a, 2018b).土壤箱式通量法可与土壤通量梯度法相结合, 实现土壤CO2释放量的相互验证以及土壤CO2储存通量的估算(Maier et al., 2011; Maier & Schack-Kirchner, 2014). ...
四个温带树种树干呼吸的时间动态及其影响因子 1 2010
... 茎干箱式通量观测系统通常可以直接使用或借鉴土壤箱式通量观测系统.根据胸径大小制作特定直径的PVC环, 密封固定在树干上直接利用土壤箱式通量观测系统进行观测(Yang et al., 2012; Hilman & Angert, 2016; Salomón et al., 2016), 或者制作适合茎干的特定箱体并与气体分析仪相连构成茎干箱式通量观测系统(Wang et al., 2019; Yang et al., 2019).茎干CO2通量是森林生态系统碳循环中的重要组成部分, 占全球森林生态系统净初级生产力的20% (Yang et al., 2016, 2019; Zhao et al., 2018).研究表明, 茎干呼吸的日变化趋势呈现单峰曲线、双峰曲线和S型曲线3种模式(石新立等, 2010).其中, 气候和植被类型等可能是不同变化趋势的决定因素.Yang等(2016)通过收集全球121篇已发表文章的数据, 研究了茎干呼吸与叶面积指数、年平均气温及年平均降水量的相关关系, 发现茎干呼吸与叶面积指数呈正相关关系, 叶面积指数、年平均气温及降水量可以解释茎干呼吸变异的74%. ...
四个温带树种树干呼吸的时间动态及其影响因子 1 2010
... 茎干箱式通量观测系统通常可以直接使用或借鉴土壤箱式通量观测系统.根据胸径大小制作特定直径的PVC环, 密封固定在树干上直接利用土壤箱式通量观测系统进行观测(Yang et al., 2012; Hilman & Angert, 2016; Salomón et al., 2016), 或者制作适合茎干的特定箱体并与气体分析仪相连构成茎干箱式通量观测系统(Wang et al., 2019; Yang et al., 2019).茎干CO2通量是森林生态系统碳循环中的重要组成部分, 占全球森林生态系统净初级生产力的20% (Yang et al., 2016, 2019; Zhao et al., 2018).研究表明, 茎干呼吸的日变化趋势呈现单峰曲线、双峰曲线和S型曲线3种模式(石新立等, 2010).其中, 气候和植被类型等可能是不同变化趋势的决定因素.Yang等(2016)通过收集全球121篇已发表文章的数据, 研究了茎干呼吸与叶面积指数、年平均气温及年平均降水量的相关关系, 发现茎干呼吸与叶面积指数呈正相关关系, 叶面积指数、年平均气温及降水量可以解释茎干呼吸变异的74%. ...
Minimising methodological biases to improve the accuracy of partitioning soil respiration using natural abundance 13C 1 2014
... 由于原理简单且操作方便, 箱式通量观测技术和方法在自然或控制条件下的土壤、植物乃至生态系统尺度温室气体通量观测研究中得到广泛应用(Kuptz et al., 2011a, 2011b; He et al., 2013; Merbold et al., 2013; Savage et al., 2014).从早期的静态箱-碱液吸收滴定(Lundegardh, 1927)和静态箱-气相色谱测定(Midwood & Millard, 2011; Imer et al., 2013)等人工离线测定模式, 发展到目前通常采用的自动箱-红外光谱测定(Plain et al., 2009; Dubbert et al., 2014; Savage et al., 2014)等自动在线测定模式.但是, 通常缺乏对观测系统设计的理论要求和假设等问题的综合考虑和评价.首先, 如何保证系统测定数据的精度和准确度, 即保证系统气密性及气体混合、箱体内外气体浓度和压力保持一致等(Xu et al., 2006; Midwood et al., 2008); 其次, 如何保证系统测定数据的代表性, 即待测气体的产生和传输不受影响等(Midwood et al., 2008; Weiss et al., 2009; Snell et al., 2014). ...
Tracking isotopic signatures of CO2 at the high altitude site Jungfraujoch with laser spectroscopy: analytical improvements and representative results 1 2013
... 根据物质守恒方程, 要求待测气体传输只受扩散过程影响, 且全部扩散至气室箱体内, 同时测定过程不能影响CO2产生和传输过程, 否则测定的CO2释放量并不能代表呼吸通量(Midwood et al., 2008; Powers et al., 2010; Kammer et al., 2011; Midwood & Millard, 2011; Sturm et al., 2013; Savage et al., 2014).首先, 应该保证测定过程不影响待测气体的产生和传输, 即不改变温湿度等环境条件以及待测气体源汇底物特征, 并且有效消除气室箱体的边缘效应, 以及不影响气体的扩散和非扩散过程.其次, 客观验证和讨论待测气体传输只受扩散过程影响且全部扩散至气室箱体内的科学假设是否成立.例如, 土壤呼吸产生的CO2主要包括地表释放的CO2和土壤中储存的CO2两部分(Hanson & Gunderson, 2009; Midwood & Millard, 2011).通常假设土壤中储存的CO2通量可以忽略不计, 则箱式通量观测技术和方法(箱式法)直接测定的“土壤CO2释放量”等于“土壤呼吸” (Maier & Schack-Kirchner, 2014).如果土壤CO2在土壤中发生明显的储存(在某些情况下浓度高达数万μmol·mol-1), 那么就会显著影响箱式法的观测结果, 导致数据不具有代表性(Wang et al., 2018a, 2018b).土壤箱式通量法可与土壤通量梯度法相结合, 实现土壤CO2释放量的相互验证以及土壤CO2储存通量的估算(Maier et al., 2011; Maier & Schack-Kirchner, 2014). ...
Short-term dynamics of abiotic and biotic soil 13CO2 effluxes after in situ13CO2 pulse labelling of a boreal pine forest 2 2009
... 稳态箱式通量观测系统是通过使箱体内气体形成开放气路, 保持箱体内外CO2浓度一致或仅略高于大气, 通过测定箱体进气口和出气口CO2浓度差计算通量, 如图1C和1D所示.根据是否增加CO2浓度调节装置, 通常可以分为两类: (1)不增加CO2浓度调节装置, 仅通过将气泵和电磁阀切换实现利用气体分析仪测定进气口(参考气)和出气口(样品气) CO2浓度并计算差值.在测定过程中, 箱体内总CO2浓度略高于大气, 同时12CO2和13CO2浓度均略高于大气(Bahn et al., 2009; Plain et al., 2009; Subke et al., 2009).(2)增加CO2浓度调节装置, 通过向箱体内注入无CO2的空气, 保持箱体进气口和出气口气体流速相同, 使箱体内总CO2浓度始终与箱体外总CO2浓度基本保持一致, 仅略高于大气, 但设计和操作较难(Midwood et al., 2006; Marron et al., 2009).稳态系统通常没有明显的CO2累积(一般低于100 μmol·mol-1)(Midwood & Milllard, 2011), 因此, 测定过程中通常不会对CO2扩散产生明显影响, 这使得稳态系统在测定δ13C时比非稳态系统更有优势(Bahn et al., 2009; Plain et al., 2009; Subke et al., 2009). ...
... ; Subke et al., 2009). ...
Relationship between fine-root exudation and respiration of two Quercus species in a Japanese temperate forest 2 2017
... 相对于土壤、植物叶片和茎干箱式通量观测系统, 根系箱式通量观测系统的难点在于适合于根系的气室箱体设计和实现(Marsden et al., 2008; Jia et al., 2013; Lai et al., 2015, 2016).传统根CO2通量观测通常将根系切断, 然后放到根呼吸气室箱体内, 但被切除的根系呼吸速率明显高于未被切除的根系(Makita et al., 2009, 2013; Sun et al., 2017).而通过原位根呼吸气室箱体设计可以很好地解决这一问题, 根呼吸箱通常使用PVC材料, 由两个半圆环或者一个封闭圆环(一端开口)构成, 将不同直径的根系放进根呼吸箱后, 形成封闭系统用于观测根呼吸CO2通量(Marsden et al., 2008; Kuptz et al., 2011a, 2011b).根系呼吸是森林土壤碳排放的重要来源, 其对土壤呼吸的贡献比例介于1/3到50%多之间(Marsden et al., 2008; Makita et al., 2009, 2012; Sun et al., 2017).结合新的高时间分辨率土壤呼吸数据集, 可以为土壤和根系呼吸的生物和环境控制机制提供新的见解(Hopkins et al., 2013).植物根系具有不同直径(粗跟和细根)和不同生理性状, 其呼吸速率模式及其对环境因子的响应具有明显差异.根系呼吸与根系直径、组织组成和特定根长等形态特征密切相关(Sun & Mao, 2011).通过根系呼吸与形态性状之间的相关关系, 可以建立区分粗根和细根呼吸的定量关系.细根对于植物生长、营养元素获取等生理功能具有重要作用, 其呼吸速率通常高于粗根, 且变异较大.在野外原位观测的结果发现, 根系直径与呼吸速率显著相关, 直径小于0.4 mm的根系呼吸速率显著高于其他根系, 且变异较大; 同时, 根系呼吸速率与根长和根的含氮量呈正相关关系(Makita et al., 2009). ...
... , 2012; Sun et al., 2017).结合新的高时间分辨率土壤呼吸数据集, 可以为土壤和根系呼吸的生物和环境控制机制提供新的见解(Hopkins et al., 2013).植物根系具有不同直径(粗跟和细根)和不同生理性状, 其呼吸速率模式及其对环境因子的响应具有明显差异.根系呼吸与根系直径、组织组成和特定根长等形态特征密切相关(Sun & Mao, 2011).通过根系呼吸与形态性状之间的相关关系, 可以建立区分粗根和细根呼吸的定量关系.细根对于植物生长、营养元素获取等生理功能具有重要作用, 其呼吸速率通常高于粗根, 且变异较大.在野外原位观测的结果发现, 根系直径与呼吸速率显著相关, 直径小于0.4 mm的根系呼吸速率显著高于其他根系, 且变异较大; 同时, 根系呼吸速率与根长和根的含氮量呈正相关关系(Makita et al., 2009). ...
Functional relationships between morphology and respiration of fine roots in two Chinese temperate tree species 1 2011
... 相对于土壤、植物叶片和茎干箱式通量观测系统, 根系箱式通量观测系统的难点在于适合于根系的气室箱体设计和实现(Marsden et al., 2008; Jia et al., 2013; Lai et al., 2015, 2016).传统根CO2通量观测通常将根系切断, 然后放到根呼吸气室箱体内, 但被切除的根系呼吸速率明显高于未被切除的根系(Makita et al., 2009, 2013; Sun et al., 2017).而通过原位根呼吸气室箱体设计可以很好地解决这一问题, 根呼吸箱通常使用PVC材料, 由两个半圆环或者一个封闭圆环(一端开口)构成, 将不同直径的根系放进根呼吸箱后, 形成封闭系统用于观测根呼吸CO2通量(Marsden et al., 2008; Kuptz et al., 2011a, 2011b).根系呼吸是森林土壤碳排放的重要来源, 其对土壤呼吸的贡献比例介于1/3到50%多之间(Marsden et al., 2008; Makita et al., 2009, 2012; Sun et al., 2017).结合新的高时间分辨率土壤呼吸数据集, 可以为土壤和根系呼吸的生物和环境控制机制提供新的见解(Hopkins et al., 2013).植物根系具有不同直径(粗跟和细根)和不同生理性状, 其呼吸速率模式及其对环境因子的响应具有明显差异.根系呼吸与根系直径、组织组成和特定根长等形态特征密切相关(Sun & Mao, 2011).通过根系呼吸与形态性状之间的相关关系, 可以建立区分粗根和细根呼吸的定量关系.细根对于植物生长、营养元素获取等生理功能具有重要作用, 其呼吸速率通常高于粗根, 且变异较大.在野外原位观测的结果发现, 根系直径与呼吸速率显著相关, 直径小于0.4 mm的根系呼吸速率显著高于其他根系, 且变异较大; 同时, 根系呼吸速率与根长和根的含氮量呈正相关关系(Makita et al., 2009). ...
Lateral diffusion and atmospheric CO2 mixing compromise estimates of rhizosphere respiration in a forest soil 2 2002
... 测定过程中, 不能改变待测气体源汇的底物特征及环境条件, 影响待测气体的产生.土壤呼吸依赖于温度和湿度等环境因素, 人为的环境条件改变影响CO2通量(Davidson et al., 2002).测定过程应尽量减少对气室内温度和湿度等条件的影响.土壤环插入土壤的过程会切断土壤中的根系.通常在测定前尽量长的时间将土壤环安装到样地, 使得生态系统能够得到有效恢复; 在不影响系统整体性的前提下, 土壤环插入深度都尽量浅(Midwood & Millard, 2011; 李建军等, 2019), 但存在表层土壤空气流动的问题(Parkin et al., 2005).风不仅能引起气室内压力变化, 也会促进大气CO2与表层土壤CO2的混合(Susfalk et al., 2002; Millard et al., 2008).土壤环插入土壤深度过浅可能会存在大气CO2与表层土壤CO2的混合, 特别是对于干燥、多空隙的土壤来说, 会引入CO2浓度测定误差, 并显著改变测定的δ13C值(Susfalk et al., 2002; Millard et al., 2008).Hutchinson和Livingston (2001)的研究表明, 干燥、多气孔或者粗颗粒的土壤需将土壤环插入土壤超过10 cm; 而细颗粒、湿润或紧实的土壤只需将土壤环插入土壤不超过5 cm.Nickerson和Risk (2009)利用同位素模型模拟了正确的土壤环插入深度对于阻止空气进入气室的重要性, 最差的情况下, 气室内CO2的δ13C富集可达到4‰. ...
... C值(Susfalk et al., 2002; Millard et al., 2008).Hutchinson和Livingston (2001)的研究表明, 干燥、多气孔或者粗颗粒的土壤需将土壤环插入土壤超过10 cm; 而细颗粒、湿润或紧实的土壤只需将土壤环插入土壤不超过5 cm.Nickerson和Risk (2009)利用同位素模型模拟了正确的土壤环插入深度对于阻止空气进入气室的重要性, 最差的情况下, 气室内CO2的δ13C富集可达到4‰. ...
Profile distribution of CO2 in an arid saline-alkali soil with gypsum and wheat straw amendments: a two-year incubation experiment 1 2018a
... 根据物质守恒方程, 要求待测气体传输只受扩散过程影响, 且全部扩散至气室箱体内, 同时测定过程不能影响CO2产生和传输过程, 否则测定的CO2释放量并不能代表呼吸通量(Midwood et al., 2008; Powers et al., 2010; Kammer et al., 2011; Midwood & Millard, 2011; Sturm et al., 2013; Savage et al., 2014).首先, 应该保证测定过程不影响待测气体的产生和传输, 即不改变温湿度等环境条件以及待测气体源汇底物特征, 并且有效消除气室箱体的边缘效应, 以及不影响气体的扩散和非扩散过程.其次, 客观验证和讨论待测气体传输只受扩散过程影响且全部扩散至气室箱体内的科学假设是否成立.例如, 土壤呼吸产生的CO2主要包括地表释放的CO2和土壤中储存的CO2两部分(Hanson & Gunderson, 2009; Midwood & Millard, 2011).通常假设土壤中储存的CO2通量可以忽略不计, 则箱式通量观测技术和方法(箱式法)直接测定的“土壤CO2释放量”等于“土壤呼吸” (Maier & Schack-Kirchner, 2014).如果土壤CO2在土壤中发生明显的储存(在某些情况下浓度高达数万μmol·mol-1), 那么就会显著影响箱式法的观测结果, 导致数据不具有代表性(Wang et al., 2018a, 2018b).土壤箱式通量法可与土壤通量梯度法相结合, 实现土壤CO2释放量的相互验证以及土壤CO2储存通量的估算(Maier et al., 2011; Maier & Schack-Kirchner, 2014). ...
Stem radial CO2 conductance affects stem respiratory CO2 fluxes in ash and birch trees 1 2019
... 茎干箱式通量观测系统通常可以直接使用或借鉴土壤箱式通量观测系统.根据胸径大小制作特定直径的PVC环, 密封固定在树干上直接利用土壤箱式通量观测系统进行观测(Yang et al., 2012; Hilman & Angert, 2016; Salomón et al., 2016), 或者制作适合茎干的特定箱体并与气体分析仪相连构成茎干箱式通量观测系统(Wang et al., 2019; Yang et al., 2019).茎干CO2通量是森林生态系统碳循环中的重要组成部分, 占全球森林生态系统净初级生产力的20% (Yang et al., 2016, 2019; Zhao et al., 2018).研究表明, 茎干呼吸的日变化趋势呈现单峰曲线、双峰曲线和S型曲线3种模式(石新立等, 2010).其中, 气候和植被类型等可能是不同变化趋势的决定因素.Yang等(2016)通过收集全球121篇已发表文章的数据, 研究了茎干呼吸与叶面积指数、年平均气温及年平均降水量的相关关系, 发现茎干呼吸与叶面积指数呈正相关关系, 叶面积指数、年平均气温及降水量可以解释茎干呼吸变异的74%. ...
Depth-dependent greenhouse gas production and consumption in an upland cropping system in northern China 1 2018b
... 根据物质守恒方程, 要求待测气体传输只受扩散过程影响, 且全部扩散至气室箱体内, 同时测定过程不能影响CO2产生和传输过程, 否则测定的CO2释放量并不能代表呼吸通量(Midwood et al., 2008; Powers et al., 2010; Kammer et al., 2011; Midwood & Millard, 2011; Sturm et al., 2013; Savage et al., 2014).首先, 应该保证测定过程不影响待测气体的产生和传输, 即不改变温湿度等环境条件以及待测气体源汇底物特征, 并且有效消除气室箱体的边缘效应, 以及不影响气体的扩散和非扩散过程.其次, 客观验证和讨论待测气体传输只受扩散过程影响且全部扩散至气室箱体内的科学假设是否成立.例如, 土壤呼吸产生的CO2主要包括地表释放的CO2和土壤中储存的CO2两部分(Hanson & Gunderson, 2009; Midwood & Millard, 2011).通常假设土壤中储存的CO2通量可以忽略不计, 则箱式通量观测技术和方法(箱式法)直接测定的“土壤CO2释放量”等于“土壤呼吸” (Maier & Schack-Kirchner, 2014).如果土壤CO2在土壤中发生明显的储存(在某些情况下浓度高达数万μmol·mol-1), 那么就会显著影响箱式法的观测结果, 导致数据不具有代表性(Wang et al., 2018a, 2018b).土壤箱式通量法可与土壤通量梯度法相结合, 实现土壤CO2释放量的相互验证以及土壤CO2储存通量的估算(Maier et al., 2011; Maier & Schack-Kirchner, 2014). ...
Differential allocation of carbon in fenced and clipped grasslands: a 13C tracer study in the semiarid Chinese Loess Plateau 1 2016
... 生态系统箱式通量观测系统主要分为两种: 冠层通量测定(不包括土壤)和生态系统通量测定(包括土壤), 可以采用明箱或暗箱测定.生态系统通量观测多应用于草本或小灌木生态系统通量观测研究(Wu et al., 2010; Hafner et al., 2012; Wei et al., 2016).高大乔木根据研究目标可以选择进行冠层或生态系统CO2及其δ13C通量观测.但由于难度较高而选择进行13CO2脉冲标记研究(Plain et al., 2009; Epron et al., 2016).通常利用不同高度的钢制脚手架环绕树的冠层, 利用高透光率的聚合物膜或有机玻璃等制作冠层箱(明箱或暗箱), 箱体的底座由两个半圆形不锈钢板制成, 钢板中间开口, 以便容纳茎干, 并固定在脚手架上(Plain et al., 2009; Epron et al., 2016).为了维持箱体内的温湿度条件, 通常配套安装空调, 利用风扇混匀冠层箱内的气体(Epron et al., 2016).Epron等(2016)利用27或38 m3的高透光率膜罩住整个桉树(Eucalyptus robusta)冠层进行脉冲标记, 利用茎干和根CO2及其δ13C通量观测箱实时观测, 结果表明, 叶片中标记的碳的储存时间非常短(21-31 h), 其在茎干中的迁移速率为0.20-0.82 m·h-1. ...
Chamber response time: a neglected issue in gas exchange measurements 1 2009
... 由于原理简单且操作方便, 箱式通量观测技术和方法在自然或控制条件下的土壤、植物乃至生态系统尺度温室气体通量观测研究中得到广泛应用(Kuptz et al., 2011a, 2011b; He et al., 2013; Merbold et al., 2013; Savage et al., 2014).从早期的静态箱-碱液吸收滴定(Lundegardh, 1927)和静态箱-气相色谱测定(Midwood & Millard, 2011; Imer et al., 2013)等人工离线测定模式, 发展到目前通常采用的自动箱-红外光谱测定(Plain et al., 2009; Dubbert et al., 2014; Savage et al., 2014)等自动在线测定模式.但是, 通常缺乏对观测系统设计的理论要求和假设等问题的综合考虑和评价.首先, 如何保证系统测定数据的精度和准确度, 即保证系统气密性及气体混合、箱体内外气体浓度和压力保持一致等(Xu et al., 2006; Midwood et al., 2008); 其次, 如何保证系统测定数据的代表性, 即待测气体的产生和传输不受影响等(Midwood et al., 2008; Weiss et al., 2009; Snell et al., 2014). ...
Evaluating calibration strategies for isotope ratio infrared spectroscopy for atmospheric 13CO2/12CO2 measurement 3 2013
... 生态系统各种生源元素在土壤、植物、动物、微生物和大气间迁移和转化的物质循环过程研究是生态系统生态学的前沿性科学问题(于贵瑞, 2009; 于振良, 2017).碳循环、氮循环和水循环是最重要的物质循环, 碳(CO2、CH4)、氮(N2O)和水汽(H2O)等温室气体的交换通量是生态系统物质循环的核心, 是地圈-生物圈-大气圈相互作用的纽带(于贵瑞和孙晓敏, 2017).同位素技术具有指示、示踪和整合功能, 可以辅助解析生态系统碳氮水的生物地球化学循环过程与机制(温学发等, 2019).稳定同位素红外光谱技术和方法的进步使碳稳定同位素比值(δ13C)和氧稳定同位素比值(δ18O)(CO2)、δ13C (CH4)、氮稳定同位素比值(δ15N)和δ18O (N2O)、氢稳定同位素比值(δD)和δ18O (H2O)等温室气体同位素组成的连续观测成为可能(Wen et al., 2008, 2013), 与箱式通量观测技术和方法结合可以实现温室气体及其同位素通量观测研究, 有助于揭示温室气体在土壤-植物-大气连续体的产生、传输和转化的相关过程与机制等(Wingate et al., 2010; Bowling et al., 2015; Epron et al., 2016). ...
... 所有箱式通量观测系统均需要对气体分析仪进行定期标定.目前常用的CO2及其δ13C气体分析仪主要基于稳定同位素红外光谱技术(Midwood & Millard, 2011; Griffis, 2013; Wen et al., 2013; Pang et al., 2016a), 但该技术自身也存在缺点, 主要表现为δ13C测量值对CO2浓度变化的非线性响应(浓度依赖性)以及对环境条件变化的敏感性导致的漂移(时间漂移).二者均对气体分析仪的精度和准确度产生影响, 需要进行适当校正和数据溯源以保证数据的可靠性(Kammer et al., 2011; Pang et al., 2016b; Chen et al., 2017, 2019).庞家平和温学发(2018)推荐利用3个或3个以上已知CO2浓度和δ13C真值的CO2标准气体涵盖待测气体CO2浓度, 进行气体分析仪的浓度依赖性校正, 同时通过设置适当的校正频率, 校正时间漂移并进行数据溯源. ...
Continuous measurement of water vapor D/H and 18O/16O isotope ratios in the atmosphere 1 2008
... 生态系统各种生源元素在土壤、植物、动物、微生物和大气间迁移和转化的物质循环过程研究是生态系统生态学的前沿性科学问题(于贵瑞, 2009; 于振良, 2017).碳循环、氮循环和水循环是最重要的物质循环, 碳(CO2、CH4)、氮(N2O)和水汽(H2O)等温室气体的交换通量是生态系统物质循环的核心, 是地圈-生物圈-大气圈相互作用的纽带(于贵瑞和孙晓敏, 2017).同位素技术具有指示、示踪和整合功能, 可以辅助解析生态系统碳氮水的生物地球化学循环过程与机制(温学发等, 2019).稳定同位素红外光谱技术和方法的进步使碳稳定同位素比值(δ13C)和氧稳定同位素比值(δ18O)(CO2)、δ13C (CH4)、氮稳定同位素比值(δ15N)和δ18O (N2O)、氢稳定同位素比值(δD)和δ18O (H2O)等温室气体同位素组成的连续观测成为可能(Wen et al., 2008, 2013), 与箱式通量观测技术和方法结合可以实现温室气体及其同位素通量观测研究, 有助于揭示温室气体在土壤-植物-大气连续体的产生、传输和转化的相关过程与机制等(Wingate et al., 2010; Bowling et al., 2015; Epron et al., 2016). ...
地球关键带视角理解生态系统碳生物地球化学过程与机制 1 2019
... 生态系统各种生源元素在土壤、植物、动物、微生物和大气间迁移和转化的物质循环过程研究是生态系统生态学的前沿性科学问题(于贵瑞, 2009; 于振良, 2017).碳循环、氮循环和水循环是最重要的物质循环, 碳(CO2、CH4)、氮(N2O)和水汽(H2O)等温室气体的交换通量是生态系统物质循环的核心, 是地圈-生物圈-大气圈相互作用的纽带(于贵瑞和孙晓敏, 2017).同位素技术具有指示、示踪和整合功能, 可以辅助解析生态系统碳氮水的生物地球化学循环过程与机制(温学发等, 2019).稳定同位素红外光谱技术和方法的进步使碳稳定同位素比值(δ13C)和氧稳定同位素比值(δ18O)(CO2)、δ13C (CH4)、氮稳定同位素比值(δ15N)和δ18O (N2O)、氢稳定同位素比值(δD)和δ18O (H2O)等温室气体同位素组成的连续观测成为可能(Wen et al., 2008, 2013), 与箱式通量观测技术和方法结合可以实现温室气体及其同位素通量观测研究, 有助于揭示温室气体在土壤-植物-大气连续体的产生、传输和转化的相关过程与机制等(Wingate et al., 2010; Bowling et al., 2015; Epron et al., 2016). ...
地球关键带视角理解生态系统碳生物地球化学过程与机制 1 2019
... 生态系统各种生源元素在土壤、植物、动物、微生物和大气间迁移和转化的物质循环过程研究是生态系统生态学的前沿性科学问题(于贵瑞, 2009; 于振良, 2017).碳循环、氮循环和水循环是最重要的物质循环, 碳(CO2、CH4)、氮(N2O)和水汽(H2O)等温室气体的交换通量是生态系统物质循环的核心, 是地圈-生物圈-大气圈相互作用的纽带(于贵瑞和孙晓敏, 2017).同位素技术具有指示、示踪和整合功能, 可以辅助解析生态系统碳氮水的生物地球化学循环过程与机制(温学发等, 2019).稳定同位素红外光谱技术和方法的进步使碳稳定同位素比值(δ13C)和氧稳定同位素比值(δ18O)(CO2)、δ13C (CH4)、氮稳定同位素比值(δ15N)和δ18O (N2O)、氢稳定同位素比值(δD)和δ18O (H2O)等温室气体同位素组成的连续观测成为可能(Wen et al., 2008, 2013), 与箱式通量观测技术和方法结合可以实现温室气体及其同位素通量观测研究, 有助于揭示温室气体在土壤-植物-大气连续体的产生、传输和转化的相关过程与机制等(Wingate et al., 2010; Bowling et al., 2015; Epron et al., 2016). ...
Photosynthetic carbon isotope discrimination and its relationship to the carbon isotope signals of stem, soil and ecosystem respiration 2 2010
... 生态系统各种生源元素在土壤、植物、动物、微生物和大气间迁移和转化的物质循环过程研究是生态系统生态学的前沿性科学问题(于贵瑞, 2009; 于振良, 2017).碳循环、氮循环和水循环是最重要的物质循环, 碳(CO2、CH4)、氮(N2O)和水汽(H2O)等温室气体的交换通量是生态系统物质循环的核心, 是地圈-生物圈-大气圈相互作用的纽带(于贵瑞和孙晓敏, 2017).同位素技术具有指示、示踪和整合功能, 可以辅助解析生态系统碳氮水的生物地球化学循环过程与机制(温学发等, 2019).稳定同位素红外光谱技术和方法的进步使碳稳定同位素比值(δ13C)和氧稳定同位素比值(δ18O)(CO2)、δ13C (CH4)、氮稳定同位素比值(δ15N)和δ18O (N2O)、氢稳定同位素比值(δD)和δ18O (H2O)等温室气体同位素组成的连续观测成为可能(Wen et al., 2008, 2013), 与箱式通量观测技术和方法结合可以实现温室气体及其同位素通量观测研究, 有助于揭示温室气体在土壤-植物-大气连续体的产生、传输和转化的相关过程与机制等(Wingate et al., 2010; Bowling et al., 2015; Epron et al., 2016). ...
... 根据研究需要, 研究者通常将上述土壤、植物叶片、茎干和根CO2及其δ13C通量观测箱体进行组合, 即可实现土壤和植物CO2及其δ13C通量的协同观测(Wingate et al., 2010; Kuptz et al., 2011a, 2011b).叶片(小枝)、茎干和土壤呼吸CO2及其δ13C协同观测系统可用于植物光合作用新合成碳水化合物的迁移转化过程研究(Bowling et al., 2008; Wingate et al., 2010; Kuptz et al., 2011a, 2011b).叶片(小枝)、茎干和土壤呼吸CO2及其δ13C间具有明显的时间滞后效应(Wingate et al., 2010; Bowling et al., 2015).对20-25 m高的大树研究发现, 土壤CO2 δ13C与冠层光合产物δ13C之间存在1-4天的滞后效应, 并且显著受控于空气相对湿度的变化(Ekblad & H?gberg, 2001).Wingate等(2010)研究发现, 与光合判别相比, 叶片呼吸释放CO2 δ13C的短期变异更小, 并且存在2-10天的延迟效应, 这可能是由于后光合分馏效应和新老碳的混合作用共同导致的. ...
Partitioning pattern of carbon flux in a Kobresia grassland on the Qinghai-Tibetan Plateau revealed by field13C pulse-labeling 1 2010
... 生态系统箱式通量观测系统主要分为两种: 冠层通量测定(不包括土壤)和生态系统通量测定(包括土壤), 可以采用明箱或暗箱测定.生态系统通量观测多应用于草本或小灌木生态系统通量观测研究(Wu et al., 2010; Hafner et al., 2012; Wei et al., 2016).高大乔木根据研究目标可以选择进行冠层或生态系统CO2及其δ13C通量观测.但由于难度较高而选择进行13CO2脉冲标记研究(Plain et al., 2009; Epron et al., 2016).通常利用不同高度的钢制脚手架环绕树的冠层, 利用高透光率的聚合物膜或有机玻璃等制作冠层箱(明箱或暗箱), 箱体的底座由两个半圆形不锈钢板制成, 钢板中间开口, 以便容纳茎干, 并固定在脚手架上(Plain et al., 2009; Epron et al., 2016).为了维持箱体内的温湿度条件, 通常配套安装空调, 利用风扇混匀冠层箱内的气体(Epron et al., 2016).Epron等(2016)利用27或38 m3的高透光率膜罩住整个桉树(Eucalyptus robusta)冠层进行脉冲标记, 利用茎干和根CO2及其δ13C通量观测箱实时观测, 结果表明, 叶片中标记的碳的储存时间非常短(21-31 h), 其在茎干中的迁移速率为0.20-0.82 m·h-1. ...
On maintaining pressure equilibrium between a soil CO2 flux chamber and the ambient air 1 2006
... 由于原理简单且操作方便, 箱式通量观测技术和方法在自然或控制条件下的土壤、植物乃至生态系统尺度温室气体通量观测研究中得到广泛应用(Kuptz et al., 2011a, 2011b; He et al., 2013; Merbold et al., 2013; Savage et al., 2014).从早期的静态箱-碱液吸收滴定(Lundegardh, 1927)和静态箱-气相色谱测定(Midwood & Millard, 2011; Imer et al., 2013)等人工离线测定模式, 发展到目前通常采用的自动箱-红外光谱测定(Plain et al., 2009; Dubbert et al., 2014; Savage et al., 2014)等自动在线测定模式.但是, 通常缺乏对观测系统设计的理论要求和假设等问题的综合考虑和评价.首先, 如何保证系统测定数据的精度和准确度, 即保证系统气密性及气体混合、箱体内外气体浓度和压力保持一致等(Xu et al., 2006; Midwood et al., 2008); 其次, 如何保证系统测定数据的代表性, 即待测气体的产生和传输不受影响等(Midwood et al., 2008; Weiss et al., 2009; Snell et al., 2014). ...
Global patterns and predictors of stem CO2 efflux in forest ecosystems 2 2016
... 茎干箱式通量观测系统通常可以直接使用或借鉴土壤箱式通量观测系统.根据胸径大小制作特定直径的PVC环, 密封固定在树干上直接利用土壤箱式通量观测系统进行观测(Yang et al., 2012; Hilman & Angert, 2016; Salomón et al., 2016), 或者制作适合茎干的特定箱体并与气体分析仪相连构成茎干箱式通量观测系统(Wang et al., 2019; Yang et al., 2019).茎干CO2通量是森林生态系统碳循环中的重要组成部分, 占全球森林生态系统净初级生产力的20% (Yang et al., 2016, 2019; Zhao et al., 2018).研究表明, 茎干呼吸的日变化趋势呈现单峰曲线、双峰曲线和S型曲线3种模式(石新立等, 2010).其中, 气候和植被类型等可能是不同变化趋势的决定因素.Yang等(2016)通过收集全球121篇已发表文章的数据, 研究了茎干呼吸与叶面积指数、年平均气温及年平均降水量的相关关系, 发现茎干呼吸与叶面积指数呈正相关关系, 叶面积指数、年平均气温及降水量可以解释茎干呼吸变异的74%. ...
Temporal and spatial variations of stem CO2 efflux of three species in subtropical China 1 2012
... 茎干箱式通量观测系统通常可以直接使用或借鉴土壤箱式通量观测系统.根据胸径大小制作特定直径的PVC环, 密封固定在树干上直接利用土壤箱式通量观测系统进行观测(Yang et al., 2012; Hilman & Angert, 2016; Salomón et al., 2016), 或者制作适合茎干的特定箱体并与气体分析仪相连构成茎干箱式通量观测系统(Wang et al., 2019; Yang et al., 2019).茎干CO2通量是森林生态系统碳循环中的重要组成部分, 占全球森林生态系统净初级生产力的20% (Yang et al., 2016, 2019; Zhao et al., 2018).研究表明, 茎干呼吸的日变化趋势呈现单峰曲线、双峰曲线和S型曲线3种模式(石新立等, 2010).其中, 气候和植被类型等可能是不同变化趋势的决定因素.Yang等(2016)通过收集全球121篇已发表文章的数据, 研究了茎干呼吸与叶面积指数、年平均气温及年平均降水量的相关关系, 发现茎干呼吸与叶面积指数呈正相关关系, 叶面积指数、年平均气温及降水量可以解释茎干呼吸变异的74%. ...
Effects of girdling on stem CO2 efflux and its temperature sensitivity in Chinese fir and sweetgum trees 2 2019
... 茎干箱式通量观测系统通常可以直接使用或借鉴土壤箱式通量观测系统.根据胸径大小制作特定直径的PVC环, 密封固定在树干上直接利用土壤箱式通量观测系统进行观测(Yang et al., 2012; Hilman & Angert, 2016; Salomón et al., 2016), 或者制作适合茎干的特定箱体并与气体分析仪相连构成茎干箱式通量观测系统(Wang et al., 2019; Yang et al., 2019).茎干CO2通量是森林生态系统碳循环中的重要组成部分, 占全球森林生态系统净初级生产力的20% (Yang et al., 2016, 2019; Zhao et al., 2018).研究表明, 茎干呼吸的日变化趋势呈现单峰曲线、双峰曲线和S型曲线3种模式(石新立等, 2010).其中, 气候和植被类型等可能是不同变化趋势的决定因素.Yang等(2016)通过收集全球121篇已发表文章的数据, 研究了茎干呼吸与叶面积指数、年平均气温及年平均降水量的相关关系, 发现茎干呼吸与叶面积指数呈正相关关系, 叶面积指数、年平均气温及降水量可以解释茎干呼吸变异的74%. ...
Effect of climatic factors on the temporal variation of stem respiration in Larix principis-rupprechtii Mayr 1 2018
... 茎干箱式通量观测系统通常可以直接使用或借鉴土壤箱式通量观测系统.根据胸径大小制作特定直径的PVC环, 密封固定在树干上直接利用土壤箱式通量观测系统进行观测(Yang et al., 2012; Hilman & Angert, 2016; Salomón et al., 2016), 或者制作适合茎干的特定箱体并与气体分析仪相连构成茎干箱式通量观测系统(Wang et al., 2019; Yang et al., 2019).茎干CO2通量是森林生态系统碳循环中的重要组成部分, 占全球森林生态系统净初级生产力的20% (Yang et al., 2016, 2019; Zhao et al., 2018).研究表明, 茎干呼吸的日变化趋势呈现单峰曲线、双峰曲线和S型曲线3种模式(石新立等, 2010).其中, 气候和植被类型等可能是不同变化趋势的决定因素.Yang等(2016)通过收集全球121篇已发表文章的数据, 研究了茎干呼吸与叶面积指数、年平均气温及年平均降水量的相关关系, 发现茎干呼吸与叶面积指数呈正相关关系, 叶面积指数、年平均气温及降水量可以解释茎干呼吸变异的74%. ...
The relationship between soil CO2 efflux and its carbon isotopic composition under non-steady-state conditions 1 2018
... 目前, 非稳态和稳态箱式通量观测系统在自然条件以及控制条件下土壤和植物根-茎-叶乃至生态系统尺度CO2及其δ13C通量观测中均得到应用(Griffis, 2013; Bowling et al., 2015; Zhou et al., 2018).商业化及研究者自制了土壤和植物CO2及其δ13C通量协同观测系统(图2A)和生态系统或冠层CO2及其δ13C通量观测系统(图2B), 通常将分析仪及箱式装置有效整合为一套全自动在线连续观测系统.值得注意的是, 如图2所示, 如果观测系统形成闭路循环气路则为非稳态系统, 而形成开路循环气路则为稳态系统. ...
 ,1,2,*
,1,2,*
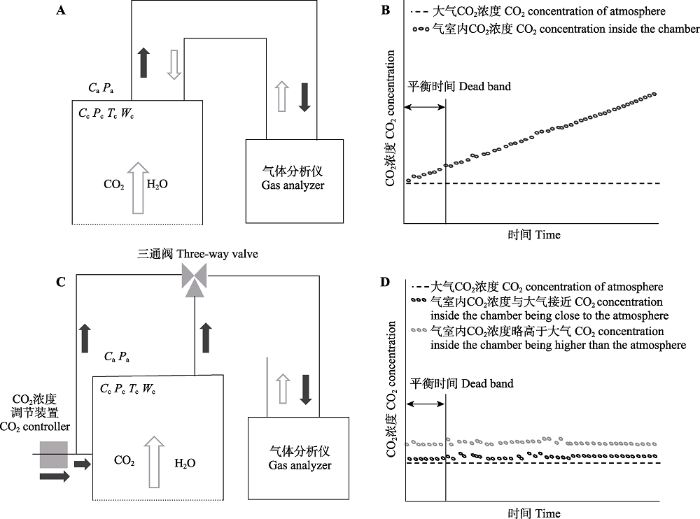 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT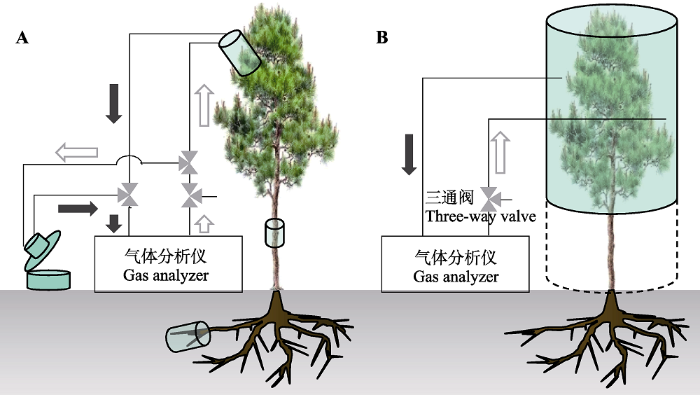 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}