Responses of phenolic acid and defensive enzyme activities to mechanical damage in Artemisia frigida
LIUMeng-Meng1, JIALi1, CHENGLu-Yun1, ZHANGHong-Qin1, ZANGXiao-Lin1, BAOYINTaogetao2, ZHANGRu-Min1, GAOYan1,*, 1The Nurturing Station for the State Key Laboratory of Subtropical Silviculture, Zhejiang Agriculture and Forestry University, Lin’an, Zhejiang 311300, China2College of Life Science, Inner Mongolia University, Hohhot 010021, China 通讯作者:* 通信作者Author for correspondence (E-mail:gaoyan1960@sohu.com) 收稿日期:2016-05-7 接受日期:2016-12-25 网络出版日期:2017-02-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家自然科学基金(31270756和31470704)、国家重点基础研究发展计划(973计划) (2014CB138805)和国家科技支撑计划项目(2011BAC07B01)
关键词:冷蒿;机械损伤;酚酸;抗氧化酶;苯丙烷代谢 Abstract Aims The study aims at understanding the effects of feed intake and trample damage on the phenolic acid formation and antioxidant enzyme activities in Artemisia frigida, and elucidating the adaptive mechanisms in A. frigida to grazing in secondary metabolites and their related enzyme activities. Methods We analyzed the phenolic acid content and the activities of polyphenol oxidase (PPO), phenylalanine ammonia-lyase (PAL) and protective enzymes in leaves and roots in A. frigida under three levels (light, moderate, and heavy) of manipulative grazing condition. The measurements of the 9 phenolic acid contents started after 6 h of the mechanical damage of the plants by using the high performance liquid chromatography (HPLC), and the enzyme activities in leaves and roots were measured by a spectrophotometry method. Important findings The light damage treatment induced productions of PPO, PAL and significantly (p < 0.05) increased antioxidant enzyme activities in the leaves and roots of A. frigida. The contents of PPO, PAL and antioxidant enzymes increased with increasing intensity of mechanical damage. Compared to the control, the content of free caffeic, syringic, ferulic and cinnamic acid in the leaves A. frigida were significantly elevated (p < 0.05) by 150.4%, 93.5%, 154.4% and 121.7%, respectively. They were significantly (p < 0.05) positively correlated with PAL activity in the moderate damage treatment. The content of free chlorogenic acid and catechol decreased by 91.1%, and 69.3%, respectively, compared with the control they had a negative correlation with PPO activity in the heavy damage treatment. The contents of gallic and protocatechuic acids increased (p < 0.05) by 280.6% and 215.7%, respectively, in the heavy damage treatment. With increasing intensity of mechanical damage, the content of 9 free phenolic acids significantly increased in roots but the increasing range was less than the one in leaves. Mechanical damage induced an increasing trend in the total amount of free and bounded phenolic acids in the leaves but a decreasing trend in the total amount of bounded phenolic acids in the roots of A. frigida. The results indicated that mechanical damage could firstly induce an increase of antioxidant enzymes and key enzymes in phenolic metabolism in A. frigida, leading to the accumulation of antioxidant substances of phenolic acids, further regulate the biosynthesis of lignins, quinones and tannins, and then enhance the resistance to mechanical damage and improved the tolerance of A. frigida to grazing.
Keywords:Artemisia frigida;mechanical damage;phenolic acid;antioxidant enzyme;phenylpropanoid pathway -->0 PDF (3538KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 刘盟盟, 贾丽, 程路芸, 张洪芹, 臧晓琳, 宝音陶格涛, 张汝民, 高岩. 冷蒿酚酸及其抗氧化防御酶活性对机械损伤的响应. 植物生态学报, 2017, 41(2): 219-230 https://doi.org/10.17521/cjpe.2016.0161 LIUMeng-Meng, JIALi, CHENGLu-Yun, ZHANGHong-Qin, ZANGXiao-Lin, BAOYINTaogetao, ZHANGRu-Min, GAOYan. Responses of phenolic acid and defensive enzyme activities to mechanical damage in Artemisia frigida. Chinese Journal of Plant Ecology, 2017, 41(2): 219-230 https://doi.org/10.17521/cjpe.2016.0161 诱导防御机制在植物自我保护过程中发挥着重要的作用, 机械损伤或动物取食都会诱导植物体内产生大量的防御物质(Kessler & Baldwin, 2002; van Poecke & Dicke, 2004)。植物通过次生代谢途径产生大量挥发性有机化合物(VOCs)、酚酸、单宁等重要的化学防御物质, 增加植物的防御能力, 对外界刺激做出响应(Agrawal, 2001)。酚酸是植物体内广泛分布的一类次生代谢产物, 其结构复杂, 种类繁多, 游离态酚酸参与很多生理过程, 如抗氧化、清除自由基、形成木质素、刺激反应和毒素活性的反应等(Brandolini et al., 2013), 结合态作为储藏形式。酚酸是植物体内通过苯丙烷酸代谢途径产生的一类重要物质, 直接参与植物的防御反应(Matkowski, 2006)。多酚氧化酶(PPO)和苯丙氨酸氨裂合酶(PAL)是苯丙烷酸代谢途径中两个重要的关键酶, 直接调控酚酸形成量和增加细胞壁木质素抵抗机械压力。因而, PAL和PPO活性的提高是诱导防御物质产生和增加防御能力的重要前提(Winkel-Shirley, 1999; Latha et al., 2009)。诸多研究表明, 植物受到损伤或动物采食后, 能够激活植物的防御反应, 诱导体内PAL和PPO活性显著升高(Chaman et al., 2003; Koussevitzky et al., 2004), 合成并积累酚类化合物(Gogoi et al., 2001; 胡增辉等, 2009; 严善春等, 2010)。超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)是植物体内主要的抗氧化酶和活性氧清除剂, 参与植物防御反应(Sharma et al., 2012)。目前, 机械损伤诱导植物酚酸积累和抗氧化酶活性研究多限于地上部分, 但是, 植物生长过程中, 地上和地下是一个协调、统一的生命整体, 丛生禾草的再生生长研究表明, 根系会对刈割做出生理响应, 通过调节生长和生理代谢, 增强植物的抵抗能力(章家恩等, 2005; Puthoff & Smigocki, 2007)。 冷蒿(Artemisia frigida)为菊科蒿属多年生小半灌木, 广泛分布于内蒙古草原带和荒漠草原带, 是草原群落的主要建群种。在连续多年过度放牧压力下草原退化演替为以冷蒿为主的单一优势群落(王炜等, 1996; 汪诗平等, 2001)。近年来, 对冷蒿的研究主要集中在群落结构特征(李衍青等, 2011; 赵康和宝音陶格涛, 2014)、营养胁迫(刘娜娜等, 2014)、化感作用(张汝民等, 2010; 左照江等, 2010)以及冷蒿挥发物对家畜采食的影响(Zhang et al., 2014)等方面, 但是关于冷蒿耐损伤机理以及酚类化合物代谢对放牧损伤的响应机制鲜见报道。本研究以冷蒿为试验材料, 研究了不同强度机械损伤对冷蒿枝叶和根系中9种游离态和结合态酚酸含量的影响, 以及苯丙烷酸代谢调控酶(PPO、PAL)和抗氧化酶(SOD、POD和CAT)活性的变化, 揭示冷蒿应激防御反应系统中酚酸合成和抗氧化酶的生理响应调控机制, 以及冷蒿抗机械损伤的适应对策; 同时为牧场的合理利用、退化草地恢复、草地生物多样性保护以及可持续发展利用提供参考。
1 材料和方法
1.1 试验材料
试验材料冷蒿采自内蒙古自治区锡林浩特毛登牧场内蒙古大学草地生态学研究基地, 地理位置44.17° N, 116.48° E, 海拔1160 m。全年平均气温为-0.4 ℃, 最冷月(1月)平均气温-22.3 ℃, 最热月(7月)平均气温18.8 ℃, ≥ 0 ℃年积温为2410 ℃, ≥ 10 ℃积温为1597.9 ℃, 无霜期91天, 草原植物生长期为150天左右。年降水量为365.6 mm, 集中于6-9月, 占全年降水量的80%左右。土壤为栗钙土。本研究区域主要草原植物为羊草(Leymus chinensis)、糙隐子草(Cleistogenes squarrosa)、克氏针茅(Stipa sareptana var. krylovii)、大针茅(Stipa grandis)、防风(Saposhnikovia divaricata)、冷蒿、瓣蕊唐松草(Thalictrum petaloideum)、阿尔泰狗哇花(Heterop- appus altaicus)等。
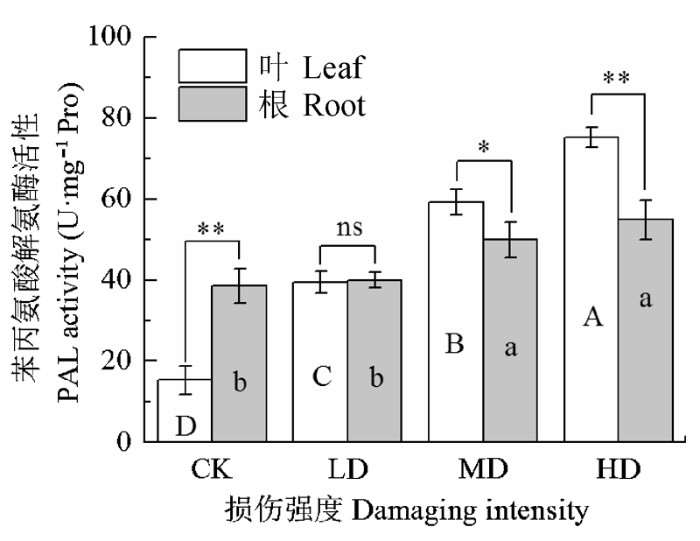
PAL是苯丙烷酸代谢途径上的调控酶, 参与酚酸和木质素的生物合成, 增加植物的抗逆性。不同机械损伤冷蒿体内PAL活性变化见图1。轻度、中度和重度处理, 枝叶PAL活性比对照分别升高了2.88倍、3.88倍和4.92倍( p < 0.05), 表明轻度损伤处理枝叶, 已经能够达到诱导阈值, 激活枝叶PAL活性, 使其迅速增加; 而根系PAL活性在中度损伤后, 才显著增加, 表明中度损伤才能够达到根系PAL活性诱导阈值。对照组枝叶PAL活性低于根系, 损伤处理后, 枝叶中PAL活性增加比根系中迅速, 表明枝叶中PAL活性对刺激响应迅速, 而根系中响应迟缓。机械损伤能够促进枝叶中酚酸的合成。 显示原图|下载原图ZIP|生成PPT 图1机械损伤对冷蒿苯丙氨酸氨裂合酶活性的影响(平均值±标准误差, n = 5)。不同大写字母表示冷蒿枝叶的差异显著, 不同小写字母表示根系的差异显著(p < 0.05, 根据LSD测验)。*, p < 0.05; **, p < 0.01。ns, 不显著。CK, 对照; LD为轻度损伤; MD为中度损伤; HD为重度损伤。 -->Fig. 1Effect of mechanical damage on the activity of phenylalanine ammonia-lyase (PAL) in Artemisia frigid (mean ± SE, n = 5). Different uppercase letters indicate significant differences of leaves and different lowercase letters denote statistically significant differences of roots from Artemisia frigida (p < 0.05, according to LSD test). *, p < 0.05; **, p < 0.01; ns, non-significant. CK, control; LD, light damaging; MD, moderate damaging; HD, heavy damaging. -->
PPO可以把酚酸氧化成醌类的物质, 防止病原菌感染。图2显示了不同损伤程度对冷蒿体内PPO活性的影响。轻度、中度和重度处理下, 枝叶中PPO活性比对照分别升高了41.7%、103.7%和197.8% (p < 0.05), 而根系PPO活性分别升高了7.0%、99.3%和163.9%, 表明轻度损伤处理冷蒿枝叶, 已经能够达到诱导阈值, 激活枝叶和根系中PPO, 使其活性迅速增加。各处理中枝叶PPO活性均高于根系, 且差异极显著, 表明枝叶PPO氧化酚酸的能力强于根系, 形成的醌类物质含量高于根系, 增强了枝叶的保护能力。 显示原图|下载原图ZIP|生成PPT 图2机械损伤对冷蒿多酚氧化酶活性的影响(平均值±标准误差, n = 5)。不同大写字母表示冷蒿枝叶的差异显著, 不同小写字母表示根系的差异显著(p < 0.05, 根据LSD测验)。*, p < 0.05; **, p < 0.01。ns, 不显著。CK, 对照; LD为轻度损伤; MD为中度损伤; HD为重度损伤。 -->Fig. 2Effect of mechanical damage on the activity of polyphenol oxidase (PPO) in Artemisia frigid (mean ± SE, n = 5). Different uppercase letters indicate significant differences of leaves and different lowercase letters denote statistically significant differences of roots from Artemisia frigida (p < 0.05, according to LSD test). *, p < 0.05; **, p <<0.01; ns, non-significant. CK, control; LD, light damaging; MD, moderate damaging; HD, heavy damaging. -->
2.3 机械损伤对冷蒿抗氧化酶活性的影响
随着机械损伤强度的增加, SOD、CAT和POD活性逐渐增强(图3), 轻度、中度和重度损伤处理, 枝叶中SOD活性比对照分别增加了1.96倍、2.36倍和4.98倍(p < 0.05); 中度和重度损伤, 根系中SOD活性比对照分别增加了68.3%和147.2% (p < 0.05) (图3A)。表明轻度损伤能够诱导枝叶SOD活性迅速增加, 达到诱导阈值; 中度损伤根系达到诱导阈值。各处理中枝叶SOD活性显著高于根系, 表明枝叶SOD活性对机械损伤的响应明显高于根系, 消除O2-·的能力很强。 显示原图|下载原图ZIP|生成PPT 图3机械损伤对冷蒿抗氧化酶活性的影响(平均值±标准误差, n = 5)。不同大写字母表示冷蒿枝叶的差异显著, 不同小写字母表示根系的差异显著(p < 0.05, 根据LSD测验)。*, p < 0.05; **, p < 0.01。ns, 不显著。CK, 对照; LD为轻度损伤; MD为中度损伤; HD为重度损伤。 -->Fig. 3Effect of mechanical damage on activity of antioxidant enzyme in Artemisia frigid (mean ± SE, n = 5). Different uppercase letters indicate significant differences of leaves and different lowercase letters denote statistically significant differences of roots from Artemisia frigida (p < 0.05, according to LSD test). *, p < 0.05; **, p < 0.01; ns, non-significant. CK, control; LD, light damaging; MD, moderate damaging; HD, heavy damaging. -->
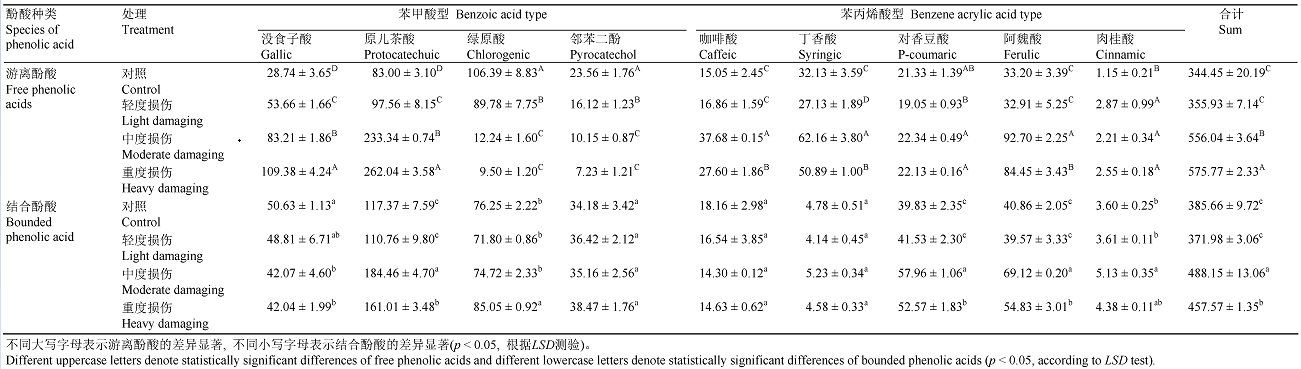
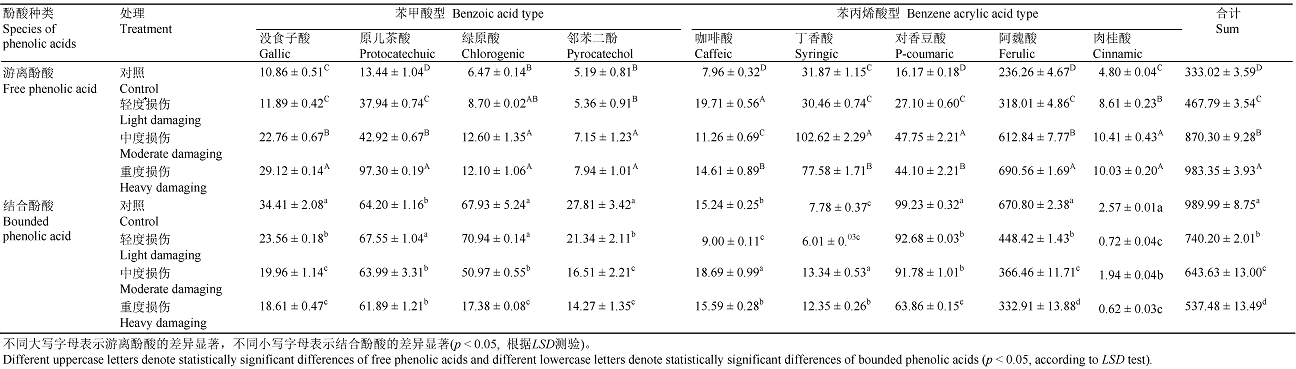
为进一步研究冷蒿体内酚酸种类和含量与抗机械损伤强度的关系, 对9种酚酸和总酚酸含量与PPO、PAL活性间进行相关性分析。由表3可见, 枝叶中游离态没食子酸、原儿茶酸、酚酸总量和阿魏酸、咖啡酸、肉桂酸含量与PAL酶活性有极显著正相关和显著正相关关系, 表明PAL酶活性增加, 促进这几种酚酸的合成, 导致含量积累, PAL酶有明显的贡献; 绿原酸和邻苯二酚含量与PPO酶活性呈极显著负相关关系, 表明PPO酶活性的增加促进绿原酸和邻苯二酚转化成醌类物质, PPO有显著贡献。枝叶中结合态原儿茶酸、肉桂酸和绿原酸、邻苯二酚、阿魏酸、对香豆酸、丁香酸、酚酸总量与PAL酶活性有显著正相关和正相关关系, 表明PAL酶活性的增加, 促进这几种结合酚酸的合成, 导致含量积累, PAL酶有贡献; 绿原酸、邻苯二酚和对香豆酸与PPO酶活性呈显著正相关关系, 没食子酸与PPO和PAL酶活性呈负相关关系。从表4可以看出, 冷蒿根系游离态原儿茶酸和没食子酸、阿魏酸和酚酸总量与PAL酶活性呈极显著正相关和显著正相关关系, 咖啡酸、丁香酸、对香豆酸、肉桂酸与PAL酶活性有正相关关系, 表明PAL酶活性的增加促进这几种酚酸的合成, 导致其含量积累, PAL酶有明显贡献; 没食子酸、原儿茶酸、绿原酸、邻苯二酚、对香豆酸、阿魏酸、肉桂酸和酚酸总量与PPO酶活性呈极显著正相关关系, 丁香酸与PPO酶活性呈显著正相关关系, 表明这几种酚酸含量增加, 在细胞内积累。结合态的7种酚酸与PAL和PPO活性呈极显著负相关和显著负相关关系, 表明这几种酚酸进行了分解, 含量下降。 Table 3 表3 表3冷蒿枝叶酚酸含量与酶活性之间相关性分析 Table 3Correlation analysis between the content of phenolic acids and related enzyme activities in the leaves of Artemisia frigid
酶 Enzyme
没食子酸 Gallic
原儿茶酸 Protocatechuic
绿原酸 Chlorogenic
邻苯二酚 Pyrocatechol
咖啡酸 Caffeic
丁香酸 Syringic
对香豆酸 P-coumaric
阿魏酸 Ferulic
肉桂酸 Cinnamic
合计 Sum
游离酚酸 Free phenolic acid
PPO
0.968**
0.861**
-0.835**
-0.871**
0.548
0.556
-0.194
0.723*
0.559
0.831**
PAL
0.859**
0.763**
-0.785**
-0.793**
0.641*
0.531
-0.069
0.690*
0.688*
0.752**
结合酚酸 Bounded phenolic acid
PPO
-0.635*
0.564
0.670*
0.742*
-0.493
-0.050
0.579*
0.487
0.480
0.559
PAL
-0.646*
0.576*
0.413
0.542
-0.489
0.126
0.572
0.551
0.617*
0.558
PAL, phenylalanine ammonia-lyase; PPO, polyphenol oxidase. *, p < 0.05; **, p < 0.01。PAL, 苯丙氨酸氨裂合酶; PPO, 多酚氧化酶。 新窗口打开 Table 4 表4 表4冷蒿根系酚酸含量与酶活性之间相关性分析 Table 4Correlation analysis between the content of phenolic acids and related enzyme activities in the roots of Artemisia frigid
酶 Enzyme
没食子酸 Gallic
原儿茶酸 Protocatechuic
绿原酸 Chlorogenic
邻苯二酚 Pyrocatechol
咖啡酸 Caffeic
丁香酸 Syringic
对香豆酸 P-coumaric
阿魏酸 Ferulic
肉桂酸 Cinnamic
合计 Sum
游离酚酸 Free phenolic acids
PPO
0.974**
0.953**
0.782**
0.861**
0.113
0.696*
0.797**
0.933**
0.711**
0.933**
PAL
0.678*
0.774**
0.478
0.511
0.254
0.346
0.483
0.618*
0.473
0.622*
结合酚酸 Bounded phenolic acids
PPO
-0.757**
-0.581*
-0.977**
-0.912**
0.419
0.772**
-0.938**
-0.611*
-0.529
-0.709**
PAL
-0.540
-0.465
-0.725**
-0.561
0.106
0.425
-0.789**
-0.635*
-0.544
-0.689*
PAL, phenylalanine ammonia-lyase; PPO, polyphenol oxidase. *, p < 0.05; **, p < 0.01。PAL, 苯丙氨酸氨裂合酶; PPO, 多酚氧化酶。 新窗口打开
AdomKK, LiuRH (2002). Antioxidant activity of grains. Journal of Agricultural and Food Chemistry, 50, 6182-6187. [本文引用: 1]
[2]
AgrawalAA (2001). Phenotypic plasticity in the interaction and evolution of species. Science, 294, 321-326. [本文引用: 1]
[3]
BaqueMA, LeeEJ, PaekKY (2010). Medium salt strength induced changes in growth, physiology and secondary metabolite contentin adventitious roots of Morinda citrifolia: The role of antioxidant enzymes and phenylalanine ammonia lyase. Plant Cell Reports, 29, 685-694. [本文引用: 1]
[4]
BradfordMM (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, 72, 248-254. [本文引用: 1]
[5]
BrandoliniA, CastoldiP, PlizzariL, HidalgoA (2013). Phenolic acids composition, total polyphenols content and antioxidant activity of Triticum monococcum, Triticum turgidum and Triticum aestivum: A two-years evaluation. Journal of Cereal Science, 58, 123-131. [本文引用: 3]
[6]
Campos-VargasR, SaltveitME (2002). Involvement of putative chemical wound signals in the induction of phenolic metabolism in wounded lettuce. Physiologia Plantarum, 114, 73-84. [本文引用: 2]
[7]
ChamanME, CopajaSV, ArgandoñaVH (2003). Relationships between salicylic acid content, phenylalanine ammonialyase (PAL) activity, and resistance of barley to aphid infestation. Journal of Agricultural and Food Chemistry, 51, 2227-2231. [本文引用: 1]
[8]
DickeM, van PoeckeRM, de BoerJG (2003). Inducible indirect defence of plants: From mechanisms to ecological functions. Basic and Applied Ecology, 4, 27-42. [本文引用: 1]
FuJM, HuangBG (2001). Involvement of antioxidants and lipid peroxidation in the adaptation of two cool-season grasses to localized drought stress. Environmental and Experimental Botany, 45, 105-114. [本文引用: 1]
[11]
GiannopolitisCN, RiesSK (1977). Superoxide dismutases I. Occurrence in higher plants. Plant Physiology, 59, 309-314. [本文引用: 1]
[12]
GillSS, TutejaN (2010). Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiology and Biochemistry, 48, 909-930. [本文引用: 1]
[13]
GogoiR, SinghDV, SrivastavaKD (2001). Phenols as a biochemical basis of resistance in wheat against Karnal bunt. Plant Pathology, 50, 470-476. [本文引用: 1]
[14]
HarutaM, PedersenJA, ConstabelCP (2001). Polyphenol oxidase and herbivore defense in trembling aspen (Populus tremuloides): cDNA cloning, expression, and potential substrates. Physiologia Plantarum, 112, 552-558. [本文引用: 1]
[15]
HuZH, YangD, ShenYB (2009). Difference of phenolic contents in leaves of Populus simonii × P. pyramidalis “Opera 8277” cuttings induced by various damages . Acta Botanica Boreali-Occidentalia Sinica, 29, 332-337. (in Chinese with English abstract)[胡增辉, 杨迪, 沈应柏 (2009). 不同损伤形式诱导合作杨叶片中酚类物质含量的差异 . 西北植物学报, 29, 332-337.] [本文引用: 2]
[16]
JiangY, JoyceDC (2003). ABA effects on ethylene production, PAL activity, anthocyanin and phenolic contents of strawberry fruit. Plant Growth Regulation, 39, 171-174. [本文引用: 2]
[17]
KesslerA, BaldwinIT (2002). Plant responses to insect herbivory: The emerging molecular analysis. Annual Review of Plant Biology, 53, 299-328. [本文引用: 1]
[18]
KestringD, MenezesLCCR, TomazCA, LimaG, RossiMN (2009). Relationship among phenolic contents, seed predation, and physical seed traits in Mimosa bimucronata plants. Journal of Plant Biology, 52, 569-576. [本文引用: 1]
[19]
KoussevitzkyS, Ne’emanE, HarelE (2004). Import of polyphenol oxidase by chloroplasts is enhanced by methyl jasmonate. Planta, 219, 412-419. [本文引用: 2]
[20]
KováčikJ, KlejdusB, HedbavnyJ, BačkorM (2010). Effect of copper and salicylic acid on phenolic metabolites and free amino acids in Scenedesmus quadricauda (Chlorophyceae). Plant Science, 178, 307-311. [本文引用: 1]
[21]
LachmanJ, AmouzK, ŠulcM, OrsákM, PivecV, HejtmánkováA, DvorákP, CeplJ (2009). Cultivar differences of total anthocyanins and anthocyanidins in red and purple- fleshed potatoes and their relation to antioxidant activity. Food Chemistry, 114, 836-843. [本文引用: 1]
[22]
LathaP, AnandT, RagupathiN, PrakasamV, SamiyappanR (2009). Antimicrobial activity of plant extracts and induction of systemic resistance in tomato plants by mixtures of PGPR strains and Zimmu leaf extract against Alternaria solani. Biological Control, 50, 85-93. [本文引用: 1]
[23]
LiYQ, SunYJ, ZhangTH, ZhaoAF, LianJ (2011). Structural characteristics of Artemisia frigid community at different succession stages in Horqin Sandy Land. Chinese Journal of Applied Ecology, 22, 1725-1730. (in Chinese with English abstract)[李衍青, 孙英杰, 张铜会, 赵爱芬, 连杰 (2011). 科尔沁沙地不同演替阶段冷蒿群落的结构特征 . 应用生态学报, 22, 1725-1730.] [本文引用: 1]
[24]
LinKM, YeFM, LinY, LiQS (2010). Research advances of phenolic functional mechanisms in soils and plants. Chinese Journal of Eco-Agriculture, 18, 1130-1137. (in Chinese with English abstract)[林开敏, 叶发茂, 林艳, 李卿叁 (2010). 酚类物质对土壤和植物的作用机制研究进展 . 中国生态农业学报, 18, 1130-1137.] [本文引用: 1]
[25]
LiuNN, TianQY, ZhangWH (2014). Comparison of adaptive strategies to phosphorus-deficient soil between dominant species Artemisia frigida and Stipa krylovii in typical steppe of Nei Mongol. Chinese Journal of Plant Ecology, 38, 905-915. (in Chinese with English abstract)[刘娜娜, 田秋英, 张文浩 (2014). 内蒙古典型草原优势种冷蒿和克氏针茅对土壤低磷环境适应策略的比较 . 植物生态学报, 38, 905-915.] [本文引用: 1]
[26]
LiuR, WangZY, LiTT, WangF, AnJ (2014). The role of chitosan in polyphenols accumulation and induction of defense enzymes in Pinus koraiensis seedlings. Chinese Journal of Plant Ecology, 38, 749-756. (in Chinese with English abstract)[刘冉, 王振宇, 李婷婷, 王芳, 安静 (2014). 壳聚糖对红松幼苗多酚积累和抗氧化防御酶的诱导作用 . 植物生态学报, 38, 749-756.] [本文引用: 2]
[27]
MatkowskiA (2006). Plant Phenolic Metabolites as Antioxidants and Mutagenesis Inhibitors. IOS Press, Amsterdam. [本文引用: 2]
[28]
MengJF, FangYL, QinMY, ZhuangXF, ZhangZW (2012). Varietal differences among the phenolic profiles and antioxidant properties of four cultivars of spine grape (Vitis davidii Foex) in Chongyi County (China). Food Chemistry, 134, 2049-2056. [本文引用: 2]
[29]
NicholsonRL, HammerschmidtR (1992). Phenolic compounds and their role in disease resistance. Annual Review of Phytopathology, 30, 369-389. [本文引用: 1]
[30]
NordbergJ, ArnerES (2001). Reactive oxygen species, antioxidants, and the mammalian thioredoxin system. Free Radical Biology and Medicine, 31, 1287-1312. [本文引用: 1]
[31]
PaulS, GoodingCB (2001). Molecular cloning and characterization of banana fruit polyphenol oxidase. Planta, 213, 748-757. [本文引用: 1]
[32]
PuthoffDP, SmigockiAC (2007). Insect feeding-induced differential expression of Beta vulgaris root genes and their regulation by defense-associated signals. Plant Cell Reports, 26, 71-84. [本文引用: 2]
[33]
RobbinsRJ (2003). Phenolic acids in foods: An overview of analytical methodology. Journal of Agricultural and Food Chemistry, 51, 2866-2887. [本文引用: 1]
[34]
SharmaP, JhaAB, DubeyRS, PessarakliM (2012). Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. Journal of Botany, 2012, 1-26. [本文引用: 2]
van PoeckeRMP, DickeM (2004). Indirect defence of plants against herbivores: Using Arabidopsis thaliana as a model plant. Plant Biology, 6, 387-401. [本文引用: 1]
[37]
WangJ, YangC, HanWQ, LiuML (2003). Effects on water- soluble carbohydrate of Artemisia frigida under different defoliation intensities. Acta Ecologica Sinica, 23, 908-913. (in Chinese with English abstract)[王静, 杨持, 韩文权, 刘美玲 (2003). 刈割强度对冷蒿可溶性碳水化合物的影响 . 生态学报, 23, 908-913.] [本文引用: 2]
[38]
WangQ, YanSC, JinH, WangYJ (2012). Systemic induction of defensive enzymes in needles of Larix gmelinii seedlings in wounded conditions. Journal of Northeast Forestry University, 40(9), 77-80. (in Chinese with English abstract)[王琪, 严善春, 金虎, 王艳军 (2012). 叶损伤对兴安落叶松防御酶活性的系统诱导 . 东北林业大学学报, 40(9), 77-80.] [本文引用: 1]
[39]
WangSP, LiYH, WangYF, ChenZZ (2001). Influence of different stocking rates on plant diversity of Artemisia frigida community in inner mongolia steppe. Acta Botanica Sinica, 43, 89-96. (in Chinese with English abstract)[汪诗平, 李永宏, 王艳芬, 陈佐忠 (2001). 不同放牧率对内蒙古冷蒿草原植物多样性的影响 . 植物学报, 43, 89-96.] [本文引用: 1]
[40]
WangW, LiuZL, HaoDY, LiangCZ (1996). Research on the restoring succession of the degenerated grassland in Inner Mongolia II. Analysis of the restoring processes. Acta Phytoecologica Sinica, 20, 460-471. (in Chinese with English abstract)[王炜, 刘钟龄, 郝敦元, 梁存柱 (1996). 内蒙古草原退化群落恢复演替的研究II. 恢复演替时间进程的分析 . 植物生态学报, 20, 460-471.] [本文引用: 1]
[41]
Winkel-ShirleyB (1999). Evidence for enzyme complexes in the phenylpropanoid and flavonoid pathways. Physiologia Plantarum, 107, 142-149. [本文引用: 2]
[42]
XuGH, GuanRF, YeXQ, ChenJC, LiuDH (2008). Composition and distribution of phenolic acids in satsuma mandarin (Citrus unshiu Marc.) during maturity. Food Science, 29(2), 137-141. (in Chinese with English abstract)[徐贵华, 关荣发, 叶兴乾, 陈健初, 刘东红 (2008). 不同成熟期蜜桔中酚酸的组成与分布 . 食品科学, 29(2), 137-141.] [本文引用: 1]
[43]
YanSC, YuanHE, WangQ, WangYJ (2010). Quantitative changes of phenolic acids in Larix gmelinii needles induced by leaf damage. Chinese Journal of Applied Ecology, 21, 1000-1006. (in Chinese with English abstract)[严善春, 袁红娥, 王琪, 王艳军 (2010). 叶损伤诱导兴安落叶松针叶中10种酚酸的变化 . 应用生态学报, 21, 1000-1006.] [本文引用: 3]
[44]
ZhangJE, LiuWG, ChenJQ, ShiYC, CaiYF (2005). Effects of different cutting intensities on above- and underground growth of Stylosanthes guianensis. Chinese Journal of Applied Ecology, 16, 1740-1744. (in Chinese with English abstract)[章家恩, 刘文高, 陈景青, 施耀才, 蔡燕飞 (2005). 不同刈割强度对牧草地上部和地下部生长性状的影响 . 应用生态学报, 16, 1740-1744.] [本文引用: 1]
[45]
ZhangRM, WangYZ, HouP, WenGS, GaoY (2010). Physiological responses to allelopathy of aquatic stem and leaf extract of Artemisia frigida in seedling of several pasture plants. Acta Ecologica Sinica, 30, 2197-2204. (in Chinese with English abstract)[张汝民, 王玉芝, 侯平, 温国胜, 高岩 (2010). 几种牧草幼苗对冷蒿茎叶水浸提液化感作用的生理响应 . 生态学报, 30, 2197-2204.] [本文引用: 1]
[46]
ZhangRM, ZhangWG, ZuoZJ, LiR, WuJH, GaoY (2014). Inhibition effects of volatile organic compounds from Artemisia frigida Willd. on the pasture grass intake by lambs. Small Ruminant Research, 121, 248-254. [本文引用: 1]
[47]
ZhaoK, BaoyinT (2014). Effect of seasonal grazing use on productivity of grassland community. Chinese Journal of Grassland, 36(1), 109-115. (in Chinese with English abstract)[赵康, 宝音陶格涛 (2014). 季节性放牧利用对典型草原群落生产力的影响 . 中国草地学报, 36(1), 109-115.] [本文引用: 2]
[48]
ZuoZJ, ZhangRM, WangY, WenGS, HouP, GaoY (2010). The composition of volatile organic compounds (VOCs) emitted from damaged Artemisia frigida Willd. plants and their effects on root growth and development of pasture plants. Acta Ecologica Sinica, 30, 5131-5139. (in Chinese with English abstract)[左照江, 张汝民, 王勇, 温国胜, 侯平, 高岩 (2010). 损伤冷蒿挥发性有机化合物(VOCs)成分分析及其对牧草根系生长发育的影响 . 生态学报, 30, 5131-5139.] [本文引用: 1]
Phenotypic plasticity in the interaction and evolution of species. 1 2001
... 诱导防御机制在植物自我保护过程中发挥着重要的作用, 机械损伤或动物取食都会诱导植物体内产生大量的防御物质(Kessler & Baldwin, 2002; van Poecke & Dicke, 2004).植物通过次生代谢途径产生大量挥发性有机化合物(VOCs)、酚酸、单宁等重要的化学防御物质, 增加植物的防御能力, 对外界刺激做出响应(Agrawal, 2001).酚酸是植物体内广泛分布的一类次生代谢产物, 其结构复杂, 种类繁多, 游离态酚酸参与很多生理过程, 如抗氧化、清除自由基、形成木质素、刺激反应和毒素活性的反应等(Brandolini et al., 2013), 结合态作为储藏形式.酚酸是植物体内通过苯丙烷酸代谢途径产生的一类重要物质, 直接参与植物的防御反应(Matkowski, 2006).多酚氧化酶(PPO)和苯丙氨酸氨裂合酶(PAL)是苯丙烷酸代谢途径中两个重要的关键酶, 直接调控酚酸形成量和增加细胞壁木质素抵抗机械压力.因而, PAL和PPO活性的提高是诱导防御物质产生和增加防御能力的重要前提(Winkel-Shirley, 1999; Latha et al., 2009).诸多研究表明, 植物受到损伤或动物采食后, 能够激活植物的防御反应, 诱导体内PAL和PPO活性显著升高(Chaman et al., 2003; Koussevitzky et al., 2004), 合成并积累酚类化合物(Gogoi et al., 2001; 胡增辉等, 2009; 严善春等, 2010).超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)是植物体内主要的抗氧化酶和活性氧清除剂, 参与植物防御反应(Sharma et al., 2012).目前, 机械损伤诱导植物酚酸积累和抗氧化酶活性研究多限于地上部分, 但是, 植物生长过程中, 地上和地下是一个协调、统一的生命整体, 丛生禾草的再生生长研究表明, 根系会对刈割做出生理响应, 通过调节生长和生理代谢, 增强植物的抵抗能力(章家恩等, 2005; Puthoff & Smigocki, 2007). ...
Medium salt strength induced changes in growth, physiology and secondary metabolite contentin adventitious roots of Morinda citrifolia: The role of antioxidant enzymes and phenylalanine ammonia lyase. 1 2010
A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. 1 1976
... 可溶性蛋白质含量测定参照Bradford (1976)的方法. ...
Phenolic acids composition, total polyphenols content and antioxidant activity of Triticum monococcum, Triticum turgidum and Triticum aestivum: A two-years evaluation. 3 2013
... 诱导防御机制在植物自我保护过程中发挥着重要的作用, 机械损伤或动物取食都会诱导植物体内产生大量的防御物质(Kessler & Baldwin, 2002; van Poecke & Dicke, 2004).植物通过次生代谢途径产生大量挥发性有机化合物(VOCs)、酚酸、单宁等重要的化学防御物质, 增加植物的防御能力, 对外界刺激做出响应(Agrawal, 2001).酚酸是植物体内广泛分布的一类次生代谢产物, 其结构复杂, 种类繁多, 游离态酚酸参与很多生理过程, 如抗氧化、清除自由基、形成木质素、刺激反应和毒素活性的反应等(Brandolini et al., 2013), 结合态作为储藏形式.酚酸是植物体内通过苯丙烷酸代谢途径产生的一类重要物质, 直接参与植物的防御反应(Matkowski, 2006).多酚氧化酶(PPO)和苯丙氨酸氨裂合酶(PAL)是苯丙烷酸代谢途径中两个重要的关键酶, 直接调控酚酸形成量和增加细胞壁木质素抵抗机械压力.因而, PAL和PPO活性的提高是诱导防御物质产生和增加防御能力的重要前提(Winkel-Shirley, 1999; Latha et al., 2009).诸多研究表明, 植物受到损伤或动物采食后, 能够激活植物的防御反应, 诱导体内PAL和PPO活性显著升高(Chaman et al., 2003; Koussevitzky et al., 2004), 合成并积累酚类化合物(Gogoi et al., 2001; 胡增辉等, 2009; 严善春等, 2010).超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)是植物体内主要的抗氧化酶和活性氧清除剂, 参与植物防御反应(Sharma et al., 2012).目前, 机械损伤诱导植物酚酸积累和抗氧化酶活性研究多限于地上部分, 但是, 植物生长过程中, 地上和地下是一个协调、统一的生命整体, 丛生禾草的再生生长研究表明, 根系会对刈割做出生理响应, 通过调节生长和生理代谢, 增强植物的抵抗能力(章家恩等, 2005; Puthoff & Smigocki, 2007). ...
Effect of copper and salicylic acid on phenolic metabolites and free amino acids in Scenedesmus quadricauda (Chlorophyceae). 1 2010
... 诸多研究表明, 游离态酚酸都具有很强的抗氧化性能, 能有效地清除自由基, 抑制膜脂过氧化和清除重金属, 植物抗氧化活性与体内酚酸含量有显著的相关关系(Matkowski, 2006; Brandolini et al., 2013; 刘冉等, 2014).植物产生防御反应的特征之一, 是酚酸含量显著升高, 植物受到机械损伤、昆虫取食等均能诱导体内酚酸含量迅速增加, 以增加植物的抵抗能力(Nicholson & Hammerschmidt, 1992; Campos & Saltveit, 2002; Kováčik et al., 2010).进一步研究发现, 黄肉马铃薯(Solanum tuberosum)抗氧化能力的主要贡献者是绿原酸、没食子酸、咖啡酸和儿茶素, 而紫肉马铃薯的主要贡献者是花青素(Lachman et al., 2009; Meng et al., 2012).苯丙烯酸类酚酸的抗氧化活性高于羟基苯甲酸类, 可能是由于苯丙烯酸类中存在CH═CH—COOH, 而羟基苯甲酸类存在COOH, 前者表现出较强的供氢能力和清除自由基的能力.因此, 咖啡酸、阿魏酸和对香豆酸的抗氧化活性高于原儿茶酸、香草酸和对羟基苯甲酸(Meng et al., 2012).本研究显示, 机械损伤后冷蒿枝叶和根系中羟基苯甲酸类和苯丙烯酸类酚酸都显著增加, 表明冷蒿体内抗氧化活性很强. ...
Cultivar differences of total anthocyanins and anthocyanidins in red and purple- fleshed potatoes and their relation to antioxidant activity. 1 2009
... 诸多研究表明, 游离态酚酸都具有很强的抗氧化性能, 能有效地清除自由基, 抑制膜脂过氧化和清除重金属, 植物抗氧化活性与体内酚酸含量有显著的相关关系(Matkowski, 2006; Brandolini et al., 2013; 刘冉等, 2014).植物产生防御反应的特征之一, 是酚酸含量显著升高, 植物受到机械损伤、昆虫取食等均能诱导体内酚酸含量迅速增加, 以增加植物的抵抗能力(Nicholson & Hammerschmidt, 1992; Campos & Saltveit, 2002; Kováčik et al., 2010).进一步研究发现, 黄肉马铃薯(Solanum tuberosum)抗氧化能力的主要贡献者是绿原酸、没食子酸、咖啡酸和儿茶素, 而紫肉马铃薯的主要贡献者是花青素(Lachman et al., 2009; Meng et al., 2012).苯丙烯酸类酚酸的抗氧化活性高于羟基苯甲酸类, 可能是由于苯丙烯酸类中存在CH═CH—COOH, 而羟基苯甲酸类存在COOH, 前者表现出较强的供氢能力和清除自由基的能力.因此, 咖啡酸、阿魏酸和对香豆酸的抗氧化活性高于原儿茶酸、香草酸和对羟基苯甲酸(Meng et al., 2012).本研究显示, 机械损伤后冷蒿枝叶和根系中羟基苯甲酸类和苯丙烯酸类酚酸都显著增加, 表明冷蒿体内抗氧化活性很强. ...
Antimicrobial activity of plant extracts and induction of systemic resistance in tomato plants by mixtures of PGPR strains and Zimmu leaf extract against Alternaria solani. 1 2009
... 诱导防御机制在植物自我保护过程中发挥着重要的作用, 机械损伤或动物取食都会诱导植物体内产生大量的防御物质(Kessler & Baldwin, 2002; van Poecke & Dicke, 2004).植物通过次生代谢途径产生大量挥发性有机化合物(VOCs)、酚酸、单宁等重要的化学防御物质, 增加植物的防御能力, 对外界刺激做出响应(Agrawal, 2001).酚酸是植物体内广泛分布的一类次生代谢产物, 其结构复杂, 种类繁多, 游离态酚酸参与很多生理过程, 如抗氧化、清除自由基、形成木质素、刺激反应和毒素活性的反应等(Brandolini et al., 2013), 结合态作为储藏形式.酚酸是植物体内通过苯丙烷酸代谢途径产生的一类重要物质, 直接参与植物的防御反应(Matkowski, 2006).多酚氧化酶(PPO)和苯丙氨酸氨裂合酶(PAL)是苯丙烷酸代谢途径中两个重要的关键酶, 直接调控酚酸形成量和增加细胞壁木质素抵抗机械压力.因而, PAL和PPO活性的提高是诱导防御物质产生和增加防御能力的重要前提(Winkel-Shirley, 1999; Latha et al., 2009).诸多研究表明, 植物受到损伤或动物采食后, 能够激活植物的防御反应, 诱导体内PAL和PPO活性显著升高(Chaman et al., 2003; Koussevitzky et al., 2004), 合成并积累酚类化合物(Gogoi et al., 2001; 胡增辉等, 2009; 严善春等, 2010).超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)是植物体内主要的抗氧化酶和活性氧清除剂, 参与植物防御反应(Sharma et al., 2012).目前, 机械损伤诱导植物酚酸积累和抗氧化酶活性研究多限于地上部分, 但是, 植物生长过程中, 地上和地下是一个协调、统一的生命整体, 丛生禾草的再生生长研究表明, 根系会对刈割做出生理响应, 通过调节生长和生理代谢, 增强植物的抵抗能力(章家恩等, 2005; Puthoff & Smigocki, 2007). ...
... 诱导防御机制在植物自我保护过程中发挥着重要的作用, 机械损伤或动物取食都会诱导植物体内产生大量的防御物质(Kessler & Baldwin, 2002; van Poecke & Dicke, 2004).植物通过次生代谢途径产生大量挥发性有机化合物(VOCs)、酚酸、单宁等重要的化学防御物质, 增加植物的防御能力, 对外界刺激做出响应(Agrawal, 2001).酚酸是植物体内广泛分布的一类次生代谢产物, 其结构复杂, 种类繁多, 游离态酚酸参与很多生理过程, 如抗氧化、清除自由基、形成木质素、刺激反应和毒素活性的反应等(Brandolini et al., 2013), 结合态作为储藏形式.酚酸是植物体内通过苯丙烷酸代谢途径产生的一类重要物质, 直接参与植物的防御反应(Matkowski, 2006).多酚氧化酶(PPO)和苯丙氨酸氨裂合酶(PAL)是苯丙烷酸代谢途径中两个重要的关键酶, 直接调控酚酸形成量和增加细胞壁木质素抵抗机械压力.因而, PAL和PPO活性的提高是诱导防御物质产生和增加防御能力的重要前提(Winkel-Shirley, 1999; Latha et al., 2009).诸多研究表明, 植物受到损伤或动物采食后, 能够激活植物的防御反应, 诱导体内PAL和PPO活性显著升高(Chaman et al., 2003; Koussevitzky et al., 2004), 合成并积累酚类化合物(Gogoi et al., 2001; 胡增辉等, 2009; 严善春等, 2010).超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)是植物体内主要的抗氧化酶和活性氧清除剂, 参与植物防御反应(Sharma et al., 2012).目前, 机械损伤诱导植物酚酸积累和抗氧化酶活性研究多限于地上部分, 但是, 植物生长过程中, 地上和地下是一个协调、统一的生命整体, 丛生禾草的再生生长研究表明, 根系会对刈割做出生理响应, 通过调节生长和生理代谢, 增强植物的抵抗能力(章家恩等, 2005; Puthoff & Smigocki, 2007). ...
... 诸多研究表明, 游离态酚酸都具有很强的抗氧化性能, 能有效地清除自由基, 抑制膜脂过氧化和清除重金属, 植物抗氧化活性与体内酚酸含量有显著的相关关系(Matkowski, 2006; Brandolini et al., 2013; 刘冉等, 2014).植物产生防御反应的特征之一, 是酚酸含量显著升高, 植物受到机械损伤、昆虫取食等均能诱导体内酚酸含量迅速增加, 以增加植物的抵抗能力(Nicholson & Hammerschmidt, 1992; Campos & Saltveit, 2002; Kováčik et al., 2010).进一步研究发现, 黄肉马铃薯(Solanum tuberosum)抗氧化能力的主要贡献者是绿原酸、没食子酸、咖啡酸和儿茶素, 而紫肉马铃薯的主要贡献者是花青素(Lachman et al., 2009; Meng et al., 2012).苯丙烯酸类酚酸的抗氧化活性高于羟基苯甲酸类, 可能是由于苯丙烯酸类中存在CH═CH—COOH, 而羟基苯甲酸类存在COOH, 前者表现出较强的供氢能力和清除自由基的能力.因此, 咖啡酸、阿魏酸和对香豆酸的抗氧化活性高于原儿茶酸、香草酸和对羟基苯甲酸(Meng et al., 2012).本研究显示, 机械损伤后冷蒿枝叶和根系中羟基苯甲酸类和苯丙烯酸类酚酸都显著增加, 表明冷蒿体内抗氧化活性很强. ...
Varietal differences among the phenolic profiles and antioxidant properties of four cultivars of spine grape (Vitis davidii Foex) in Chongyi County (China). 2 2012
... 诸多研究表明, 游离态酚酸都具有很强的抗氧化性能, 能有效地清除自由基, 抑制膜脂过氧化和清除重金属, 植物抗氧化活性与体内酚酸含量有显著的相关关系(Matkowski, 2006; Brandolini et al., 2013; 刘冉等, 2014).植物产生防御反应的特征之一, 是酚酸含量显著升高, 植物受到机械损伤、昆虫取食等均能诱导体内酚酸含量迅速增加, 以增加植物的抵抗能力(Nicholson & Hammerschmidt, 1992; Campos & Saltveit, 2002; Kováčik et al., 2010).进一步研究发现, 黄肉马铃薯(Solanum tuberosum)抗氧化能力的主要贡献者是绿原酸、没食子酸、咖啡酸和儿茶素, 而紫肉马铃薯的主要贡献者是花青素(Lachman et al., 2009; Meng et al., 2012).苯丙烯酸类酚酸的抗氧化活性高于羟基苯甲酸类, 可能是由于苯丙烯酸类中存在CH═CH—COOH, 而羟基苯甲酸类存在COOH, 前者表现出较强的供氢能力和清除自由基的能力.因此, 咖啡酸、阿魏酸和对香豆酸的抗氧化活性高于原儿茶酸、香草酸和对羟基苯甲酸(Meng et al., 2012).本研究显示, 机械损伤后冷蒿枝叶和根系中羟基苯甲酸类和苯丙烯酸类酚酸都显著增加, 表明冷蒿体内抗氧化活性很强. ...

 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT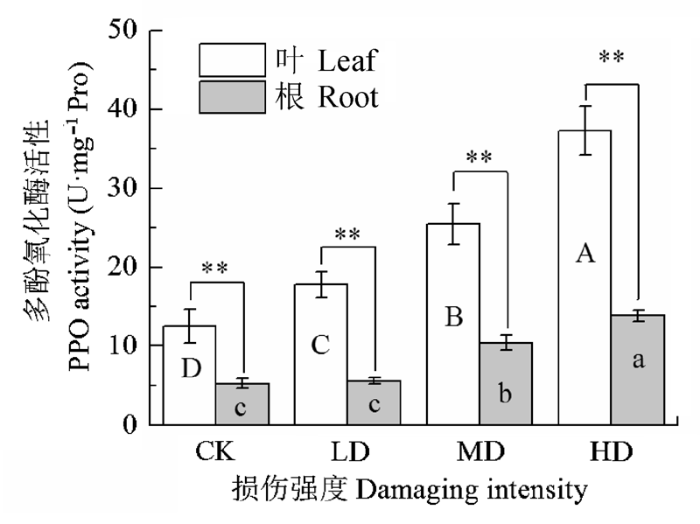 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT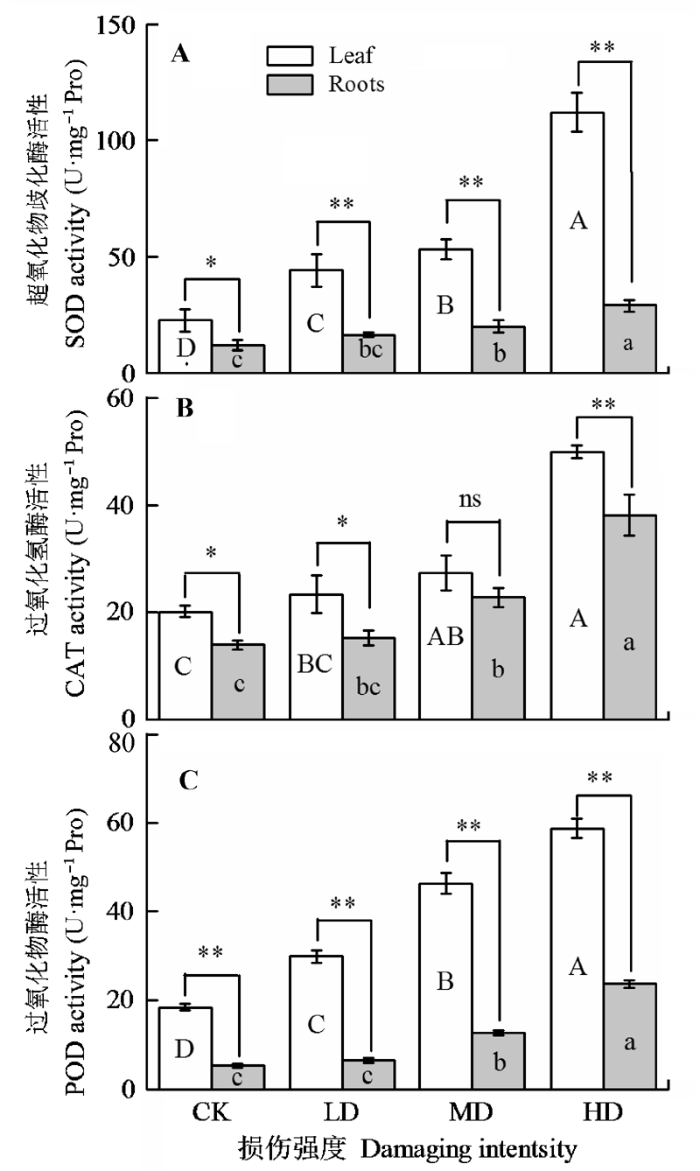 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}