Litter standing crop of shrubland ecosystems in southern China
GEJie-Lin1, XIONGGao-Ming1, LIJia-Xiang1, XUWen-Ting1, ZHAOChang-Ming1, LUZhi-Jun2, LIYue-Lin3, XIEZong-Qiang1,*, 1State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China2Key Laboratory of Aquatic Botany and Watershed Ecology, Wuhan Botanical Garden, Chinese Academy of Sciences, Wuhan 430074, China3South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China 通讯作者:* 通信作者Author for correspondence (E-mail: xie@ibcas.ac.cn) 责任编辑:GEJie-LinXIONGGao-MingLIJia-XiangXUWen-TingZHAOChang-MingLUZhi-JunLIYue-LinXIEZong-Qiang 收稿日期:2016-06-14 接受日期:2016-11-10 网络出版日期:2017-01-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:中国科学院战略性先导科技专项 (XDA05050302)
关键词:凋落物的累积;空间格局;碳转换系数;年平均气温;土壤属性;气候 Abstract Aims Litter is an important component of terrestrial ecosystems, which plays significant roles in carbon and nutrient cycles. Quantifying regional-scale pattern of litter standing crop would improve our understanding in the mechanism of the terrestrial ecosystem carbon cycle, also with help in predicting the responses of carbon cycle of terrestrial ecosystems to future climate change. Our objective was to examine variation in litter standing crop of shrublands along the environmental gradients in southern China. Methods During 2011-2014, we investigated the litter standing crop at 453 shrublands sites by the stratified random sampling, reflecting climatic and soil attributes across southern China. Important findings We found that the mean value of litter standing crop in these shrubland ecosystems across southern China was 0.32 kg·m-2. It was 68% of forest litter standing crop (0.47 kg·m-2) and was five times higher than that in grasslands (0.06 kg·m-2) in China. Litter standing crop increased with latitude. Our results showed that litter standing crop was negatively correlated with mean annual temperature, soil total P and soil pH, but not significantly correlated with other environmental variables, including mean annual precipitation, soil carbon, nitrogen and soil organic matter. The conversion coefficient of carbon in litter standing crop was 0.41, which is significantly lower than that of vegetation in shrublands (0.50), resulting in an overestimate in carbon storage of litter standing crop in shrubland up to 22% by applying wrong conversion coefficient. We concluded that litter standing crop of shrublands is an important component in terrestrial ecosystems. Mean annual temperature was the most important environmental variable, accounting for the variation in litter standing crop of shrublands in southern China. To our best of knowledge, this is the first study to quantify variation in litter standing crop of shrublands at the regional scale. Therefore, our study will have important implications for assessing the carbon budget of terrestrial ecosystems in China.
研究区位于我国南方江苏、浙江、安徽、湖北、江西、福建、广东、湖南、重庆、广西、海南等11个省市自治区。地理范围18.26°-34.63° N, 105.17- 121.26° E, 海拔0-1865 m。各省调查地点的气候和土壤状况详见表1。 Table 1 表1 表1所研究省市自治区的地理位置、气候和土壤养分概况 Table 1General information on geographic location, climate and soil for different provinces, municipalities and autonomous regions
中国南方灌丛生态系统凋落物现存量存在较大的标准偏差和变异系数, 呈现出正偏态分布(图1)。灌丛凋落物现存量的平均值为0.32 kg·m-2, 最小值为0.03 kg·m-2, 最大值为2.90 kg·m-2。单因素方差分析发现, 常绿和落叶灌丛的凋落物现存量差别不显著(p > 0.05)。该地区灌丛生态系统凋落物现存量的碳转换系数为0.41。 显示原图|下载原图ZIP|生成PPT 图1中国南方灌丛凋落物现存量的频度分布图。 -->Fig. 1Frequency distribution of litter standing crop of shrublands in southern China. -->
2.2 凋落物现存量的空间分布格局
中国南方灌丛生态系统凋落物现存量存在较弱的线性空间分布格局。因此, 为了更清楚地探讨它们的空间分布格局, 本研究将数据按照2°的经度和纬度间隔进行整合, 以揭示凋落物现存量随经纬度之间的变化趋势。结果发现: 中国南方灌丛凋落物的现存量在不同纬度和经度间表现出较大的变异特征; 灌丛凋落物现存量存在显著的纬度格局(F = 4.96, p < 0.001), 随着纬度的增加而增加(p < 0.001), 但经度格局不明显(p > 0.05)(图2)。 显示原图|下载原图ZIP|生成PPT 图2中国南方灌丛凋落物现存量随经纬度的变化(平均值±标准误差)。A, 纬度。B, 经度。 -->Fig. 2Variation in litter standing crop of shrublands across the latitudinal and longitudinal gradients in southern China (mean ± SE). A, Latitude. B, Longitude. -->
2.3 凋落物现存量与环境因子之间的关系
简单线性回归分析发现, 凋落物现存量与年平均气温、土壤磷含量、pH值和土壤容重呈现显著的负相关关系(p < 0.05), 与干燥度正相关(p < 0.05), 而与其他环境因子(年降水量、土壤碳、氮以及有机碳含量)的相关性不显著(p > 0.05)(图3)。进一步采用逐步向后回归分析表明, 凋落物现存量与年平均气温、土壤全磷含量和土壤pH值显著负相关(p < 0.05)。方差分解发现年平均气温对凋落物现存量的变异解释率最大, 占上述多元线性模型解释率的67.33%。 显示原图|下载原图ZIP|生成PPT 图3中国南方灌丛凋落物现存量与环境因子的关系。本文仅显示凋落物现存量与环境因子关系显著的图。A-E, 现存量与单个环境因子(年平均气温、土壤全磷含量、土壤容重和干燥度)的关系。F, 现存量与年平均气温(MAT)、土壤全磷含量和土壤pH值之间的多元线性回归关系。 -->Fig. 3Variation in litter standing crop of shrublands of southern China in relation to environmental factors in China. We only showed the figures that depicted the significant relationship between litter standing crop and environmental factors. A-E, The relationship between litter standing crop and single environmental variables including mean annual temperature (MAT), soil total phosphorus (P) content, soil pH value, soil bulk density and climatic dryness (Idw). F, Observed value vs. predicted value of litter standing crop from multiple regression using mean annual temperature (MAT), soil P content and soil pH value as predictors. -->
AertsR (1997). Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: A triangular relationship .Oikos, 79, 439-449. [本文引用: 1]
[2]
ChaveJ, NavarreteD, AlmeidaS, AlvarezE, AragãoLE, BonalD, ChâteletP, Silva-EspejoJ, GoretJY, HildebrandP (2010). Regional and seasonal patterns of litterfall in tropical South America .Biogeosciences, 7, 43-55. [本文引用: 1]
[3]
ChenSS, LiuHY, GuoDL (2010). Litter stocks and chemical quality of natural birch forest along temperature and precipitation gradients in eastern Inner Mongolia, China .Chinese Journal of Plant Ecology, 34, 1007-1015. (in Chinese with English abstract)[陈莎莎, 刘鸿雁, 郭大立 (2010). 内蒙古东部天然白桦林的凋落物性质和储量及其随温度和降水梯度的变化格局 . 植物生态学报, 34, 1007-1015.] [本文引用: 1]
[4]
DescheemaekerK, MuysB, NyssenJ, PoesenJ, RaesD, HaileM, DeckersJ (2006). Litter production and organic matter accumulation in exclosures of the Tigray highlands, Ethiopia . Forest Ecology and Management, 233, 21-35. [本文引用: 4]
[5]
DomkeGM, PerryCH, WaltersBF, WoodallCW, RussellMB, SmithJE (2016). Estimating litter carbon stocks on forest land in the United States .Science of Total Environment, 557, 469-478. [本文引用: 4]
[6]
DorrepaalE, CornelissenJHC, AertsR, WallénB, van LogtestijnRSP (2005). Are growth forms consistent predictors of leaf litter quality and decomposability across peatlands along a latitudinal gradient? Journal of Ecology, 93, 817-828. [本文引用: 1]
[7]
FangJY, HuangY, ZhuJL, SunWJ, HuHF (2015a). Carbon budget of forest ecosystems and its driving forces .China Basic Science, 3, 20-25. (in Chinese with English abstract)[方精云, 黄耀, 朱江玲, 孙文娟, 胡会峰 (2015a). 森林生态系统碳收支及其影响机制 . 中国基础科学, 3, 20-25.] [本文引用: 1]
[8]
FangJY, YuGR, RenXB, LiuGH, ZhaoXQ (2015b). Carbon sequestration in China’s terrestrial ecosystems under climate change—Progress on ecosystem carbon sequestration from the CAS Strategic Priority Research Program . Bulletin of Chinese Academy of Sciences, 30, 848-857. (in Chinese with English abstract)[方精云, 于贵瑞, 任小波, 刘国华, 赵新全 (2015b). 中国陆地生态系统固碳效应——中国科学院战略性先导科技专项“应对气候变化的碳收支认证及相关问题”之生态系统固碳任务群研究进展 . 中国科学院院刊, 30, 848-857. ] [本文引用: 1]
[9]
García-PalaciosP, McKieBG, HandaIT, FrainerA, HättenschwilerS (2016). The importance of litter traits and decomposers for litter decomposition: A comparison of aquatic and terrestrial ecosystems within and across biomes .Functional Ecology, 30, 819-829. [本文引用: 1]
[10]
HijmansRJ, CameronSE, ParraJL, JonesPG, JarvisA (2005). Very high resolution interpolated climate surfaces for global land areas .International Journal of Climatology, 25, 1965-1978. [本文引用: 1]
[11]
HilliS, StarkS, DeromeJ (2010). Litter decomposition rates in relation to litter stocks in boreal coniferous forests along climatic and soil fertility gradients .Applied Soil Ecology, 46, 200-208. [本文引用: 3]
[12]
HuHF, WangZH, LiuGH, FuBJ (2006). Vegetation carbon storage of major shrublands in China .Journal of Plant Ecology (Chinese Version), 30, 539-544. (in Chinese with English abstract)[胡会峰, 王志恒, 刘国华, 傅伯杰 (2006). 中国主要灌丛植被碳储量 . 植物生态学报, 30, 539-544.] [本文引用: 4]
[13]
LiQ, ZhouDW, ChenXY (2014). The accumulation, decomposition and ecological effects of above-ground litter in terrestrial ecosystem .Acta Ecologica Sinica, 34, 3807-3819. (in Chinese with English abstract)[李强, 周道玮, 陈笑莹 (2014). 地上枯落物的累积、分解及其在陆地生态系统中的作用 . 生态学报, 34, 3807-3819.] [本文引用: 1]
[14]
LiuC, WestmanCJ, BergB, KutschW, WangGZ, ManR, IlvesniemiH (2004). Variation in litterfall-climate relationships between coniferous and broadleaf forests in Eurasia .Global Ecology and Biogeography, 13, 105-114. [本文引用: 1]
[15]
LiuGS (1997). Soil Physical and Chemical Analysis and the Profile Description. China Standard Publishing House, Beijing. (in Chinese)[刘光崧 (1997). 土壤理化分析与剖面描述. 中国标准出版社, 北京.] [本文引用: 1]
[16]
LüGH, LiRP, WenRH, XieYB, WangXY (2014). Analysis of climatic effects on the components of forest litterfall production .Chinese Agricultural Science Bulletin, 30, 1-6. (in Chinese with English abstract)[吕国红, 李荣平, 温日红, 谢艳兵, 王笑影 (2014). 森林凋落物组分的气象影响分析 . 中国农学通报, 30, 1-6. ] [本文引用: 2]
[17]
MaYZ, ChengDL, ZhongQL, JinBJ, XuCB, HuB (2013). Allometric relationships among different components of forest litterfall in China .Chinese Journal of Plant Ecology, 37, 1071-1079. (in Chinese with English abstract)[马玉珠, 程栋梁, 钟全林, 靳冰洁, 徐朝斌, 胡波 (2013). 中国森林凋落物不同组分异速比例关系 . 植物生态学报, 37, 1071-1079. ] [本文引用: 1]
[18]
MartyC, HouleD, GagnonC (2015). Variation in stocks and distribution of organic C in soils across 21 eastern Canadian temperate and boreal forests .Forest Ecology and Management, 345, 29-38. [本文引用: 1]
[19]
MatthewsE (1997). Global litter production, pools, and turnover times: Estimates from measurement data and regression models . Journal of Geophysical Research: Atmospheres, 102, 18771-18800. [本文引用: 1]
ParsonsSA, CongdonRA, LawlerIR (2014a). Determinants of the pathways of litter chemical decomposition in a tropical region .New Phytologist, 203, 873-882. [本文引用: 1]
[22]
ParsonsSA, CongdonRA, ShooLP, Valdez-RamirezV, WilliamsSE (2014b). Spatial variability in litterfall, litter standing crop and litter quality in a tropical rain forest region .Biotropica, 46, 378-386. [本文引用: 2]
[23]
PengSL, LiuQ (2002) The dynamics of forest litter and its responses to global warming .Acta Ecologica Sinica, 22, 1534-1544. (in Chinese with English abstract)[彭少麟, 刘强 (2002). 森林凋落物动态及其对全球变暖的响应 . 生态学报, 22, 1534-1544. ] [本文引用: 1]
[24]
PiaoS, FangJ, CiaisP, PeylinP, HuangY, SitchS, WangT (2009). The carbon balance of terrestrial ecosystems in China .Nature, 458, 1009-1013. [本文引用: 1]
[25]
PitkänenA, SimolaH, TurunenJ (2012). Dynamics of organic matter accumulation and decomposition in the surface soil of forestry-drained peatland sites in Finland .Forest Ecology and Management, 284, 100-106. [本文引用: 1]
[26]
Portillo-EstradaM, PihlatieM, KorhonenJF, LevulaJ, FrumauAK, IbromA, LembrechtsJJ, MorillasL, HorváthL, JonesSK (2016). Climatic controls on leaf litter decomposition across European forests and grasslands revealed by reciprocal litter transplantation experiments .Biogeosciences, 13, 1621-1633. [本文引用: 1]
[27]
Ray-MukherjeeJ, NimonK, MukherjeeS, MorrisDW, SlotowR, HamerM (2014). Using commonality analysis in multiple regressions: A tool to decompose regression effects in the face of multicollinearity .Methods in Ecology and Evolution, 5, 320-328. [本文引用: 1]
[28]
SalinasN, MalhiY, MeirP, SilmanM, Roman CuestaR, HuamanJ, SalinasD, HuamanV, GibajaA, MamaniM (2011). The sensitivity of tropical leaf litter decomposition to temperature: Results from a large-scale leaf translocation experiment along an elevation gradient in Peruvian forests .New Phytologist, 189, 967-977. [本文引用: 1]
[29]
SchreegLA, MackMC, TurnerBL (2013). Nutrient-specific solubility patterns of leaf litter across 41 lowland tropical woody species . Ecology, 94, 94-105. [本文引用: 1]
[30]
SimmonsJA, FernandezIJ, BriggsRD, DelaneyMT (1996). Forest floor carbon pools and fluxes along a regional climate gradient in Maine, USA .Forest Ecology and Management, 84, 81-95. [本文引用: 1]
[31]
SunX, KangH, ChenHYH, BergB, BartelsSF, LiuC (2016). Biogeographic patterns of nutrient resorption from Quercus variabilis Blume leaves across China . Plant Biology, 18, 505-513. [本文引用: 1]
[32]
TangSS, YangWQ, YinR, XiongL, WangHP, WangB, ZhangY, PengYJ, ChenQS, XuZF (2014). Spatial characteristics in decomposition rate of foliar litter and controlling factors in Chinese forest ecosystems .Chinese Journal of Plant Ecology, 38, 529-539. (in Chinese with English abstract)[唐仕姗, 杨万勤, 殷睿, 熊莉, 王海鹏, 王滨, 张艳, 彭艳君, 陈青松, 徐振锋 (2014). 中国森林生态系统凋落叶分解速率的分布特征及其控制因子 . 植物生态学报, 38, 529-539.] [本文引用: 1]
[33]
Technical Manual Writing Group of Ecosystem Carbon Sequestration Project (2015). Observation and Investigation for Carbon Sequestration in Terrestrial Ecosystems. Science Press, Beijing. (in Chinese)[生态系统固碳项目技术规范编写组 (2015). 生态系统固碳观测与调查技术规范. 科学出版社, 北京. ] [本文引用: 1]
[34]
TwilleyRW, LugoAE, Patterson-ZuccaC (1986). Litter production and turnover in basin mangrove forests in southwest Florida .Ecology, 67, 670-683. [本文引用: 1]
[35]
VergutzL, ManzoniS, PorporatoA, NovaisRF, JacksonRB (2012). Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants .Ecological Monograph, 82, 205-220. [本文引用: 1]
[36]
WangSL, ChenCY (2010). The Ecology of Forest Litter. Science Press, Beijing.(in Chinese) [汪思龙, 陈楚莹 (2010). 森林残留物生态学. 科学出版社, 北京.] [本文引用: 1]
[37]
WenD, HeNP (2016). Spatial patterns of litter density and their controlling factors in forest and grasslands of China . Acta Ecologica Sinica, 36, 2876-2884. (in Chinese with English abstract)[温丁, 何念鹏 (2016). 中国森林和草地凋落物现存量的空间分布格局及其控制因素 . 生态学报, 36, 2876-2884.] [本文引用: 5]
[38]
WestcottV, EnrightN, MillerB, FontaineJ, LadeJ, LamontB (2014). Biomass and litter accumulation patterns in species-rich shrublands for fire hazard assessment .International Journal of Wildland Fire, 23, 860-871. [本文引用: 1]
[39]
WuZY (1980). Vegetation of China. Science Press, Beijing. (in Chinese)[吴征镒 (1980). 中国植被. 科学出版社, 北京.] [本文引用: 2]
[40]
YanER, WangXH, ZhouW (2008). Characteristics of litterfall in relation to soil nutrients in mature and degraded evergreen broad-leaved forests of Tiantong, East China .Journal of Plant Ecology (Chinese Version), 32, 1-12. (in Chinese with English abstract)[阎恩荣, 王希华, 周武 (2008). 天童常绿阔叶林不同退化群落的凋落物特征及与土壤养分动态的关系 . 植物生态学报, 32, 1-12.] [本文引用: 1]
[41]
ZhangD, HuiD, LuoY, ZhouG (2008). Rates of litter decomposition in terrestrial ecosystems: Global patterns and controlling factors .Journal of Plant Ecology, 1, 85-93. [本文引用: 1]
[42]
ZhangX, WangW (2015). Control of climate and litter quality on leaf litter decomposition in different climatic zones .Journal of Plant Research, 128, 791-802. [本文引用: 3]
[43]
ZhangXP, WangXP, ZhuB, ZongZJ, PengCH, FangJY (2008). Litter fall production in relation to enviromental factors in northeast China’s forests .Journal of Plant Ecology (Chinese Version), 32, 1031-1040. (in Chinese with English abstract)[张新平, 王襄平, 朱彪, 宗占江, 彭长辉, 方精云 (2008). 我国东北主要森林类型的凋落物产量及其影响因素 . 植物生态学报, 32, 1031-1040.] [本文引用: 2]
[44]
ZhaoHM, HuangG, MaJ, LiY, ZhouL (2012). Responses of surface litter decomposition to seasonal water addition in desert .Chinese Journal of Plant Ecology, 36, 471-482. (in Chinese with English abstract)[赵红梅, 黄刚, 马健, 李彦, 周丽 (2012). 荒漠区地表凋落物分解对季节性降水增加的响应 . 植物生态学报, 36, 471-482.] [本文引用: 1]
[45]
ZhengL, LuLH (2012). Standing crop and nutrient characteristics of forest floor litter in China .Journal of Northwest Forestry University, 27, 63-69. (in Chinese with English abstract)[郑路, 卢立华 (2012). 我国森林地表凋落物现存量及养分特征 . 西北林学院学报, 27, 63-69.] [本文引用: 1]
[46]
ZhouX, TalleyM, LuoY (2009). Biomass, litter, and soil respiration along a precipitation gradient in southern Great Plains, USA .Ecosystems, 12, 1369-1380. [本文引用: 1]
Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: A triangular relationship 1 1997
... 陆地生态系统凋落物的现存量受到多种环境因素共同控制, 大尺度上水分和热量通过控制植被类型影响凋落物的产量和分解, 进而影响凋落物的现存量(Zhou et al., 2009; Chave et al., 2010).本研究发现, 中国南方灌丛生态系统的凋落物现存量地理格局受到年平均气温、土壤pH值以及土壤全磷含量等因子的制约, 但最重要的环境因子是年平均气温.这与前人对凋落物产量和分解的研究结果(Desch- eemaeker et al., 2006; Pitkänen et al., 2012)一致.年平均气温是植物生长所需要的基本环境条件, 其年际间变化影响植物的物候变化, 从而制约着凋落物的产量.凋落物现存量受到凋落物生产和分解的制约(Hill et al., 2010).大量研究表明, 凋落物的产量与年平均气温显著正相关(张新平等, 2008; Salinas et al., 2011; 吕国红等, 2014), 不考虑凋落物的分解时, 凋落物的现存量应与年平均气温成正比, 但本研究发现凋落物现存量与年平均气温成反比.因此, 凋落物的分解可能是影响中国南方灌丛凋落物现存量空间分布的关键过程.上述推断与前人关于凋落物分解自然气候梯度的研究(Zhang et al., 2008; Hilli et al., 2010)一致.不同的研究发现, 随着纬度增加, 凋落物的分解速率下降, 温度升高可以显著加快不同生态系统的地表凋落物的分解(Aerts, 1997; Dorrepaal et al., 2005; Zhang & Wang, 2015).例如, Dorrepaal等(2005)研究欧洲西北部的灌木物种分解时发现, 年平均气温是与灌木树种凋落物分解速率最密切的控制因子.同样Hilli等(2010)研究森林凋落物分解对凋落物储量的影响时发现凋落物分解过程是凋落物储量空间变异的最为关键过程.本研究位于热带和亚热带地区, 温度是影响灌丛凋落物产量和分解的限制因子, 对凋落物分解的影响程度更突出, 因此年平均气温对凋落物生产和分解之间作用的差异性是导致中国南方灌丛生态系统凋落物现存量空间分异的主要原因. ...
Regional and seasonal patterns of litterfall in tropical South America 1 2010
... 陆地生态系统凋落物的现存量受到多种环境因素共同控制, 大尺度上水分和热量通过控制植被类型影响凋落物的产量和分解, 进而影响凋落物的现存量(Zhou et al., 2009; Chave et al., 2010).本研究发现, 中国南方灌丛生态系统的凋落物现存量地理格局受到年平均气温、土壤pH值以及土壤全磷含量等因子的制约, 但最重要的环境因子是年平均气温.这与前人对凋落物产量和分解的研究结果(Desch- eemaeker et al., 2006; Pitkänen et al., 2012)一致.年平均气温是植物生长所需要的基本环境条件, 其年际间变化影响植物的物候变化, 从而制约着凋落物的产量.凋落物现存量受到凋落物生产和分解的制约(Hill et al., 2010).大量研究表明, 凋落物的产量与年平均气温显著正相关(张新平等, 2008; Salinas et al., 2011; 吕国红等, 2014), 不考虑凋落物的分解时, 凋落物的现存量应与年平均气温成正比, 但本研究发现凋落物现存量与年平均气温成反比.因此, 凋落物的分解可能是影响中国南方灌丛凋落物现存量空间分布的关键过程.上述推断与前人关于凋落物分解自然气候梯度的研究(Zhang et al., 2008; Hilli et al., 2010)一致.不同的研究发现, 随着纬度增加, 凋落物的分解速率下降, 温度升高可以显著加快不同生态系统的地表凋落物的分解(Aerts, 1997; Dorrepaal et al., 2005; Zhang & Wang, 2015).例如, Dorrepaal等(2005)研究欧洲西北部的灌木物种分解时发现, 年平均气温是与灌木树种凋落物分解速率最密切的控制因子.同样Hilli等(2010)研究森林凋落物分解对凋落物储量的影响时发现凋落物分解过程是凋落物储量空间变异的最为关键过程.本研究位于热带和亚热带地区, 温度是影响灌丛凋落物产量和分解的限制因子, 对凋落物分解的影响程度更突出, 因此年平均气温对凋落物生产和分解之间作用的差异性是导致中国南方灌丛生态系统凋落物现存量空间分异的主要原因. ...
内蒙古东部天然白桦林的凋落物性质和储量及其随温度和降水梯度的变化格局 1 2010
... 碳转换系数是研究陆地生态系统碳收支的重要参数.本研究发现, 中国南方灌丛凋落物现存量的碳转换系数为0.41, 低于前人研究灌丛生态系统所采用的植被活体碳转换系数(0.50)(胡会峰等, 2006).因此在研究灌丛生态系统碳储量时, 采用常用的植被活体碳转换系数可能会高估灌丛生态系统凋落物碳储量22%.凋落物现存量的碳转换系数与碳浓度相关, 受到环境因子、凋落物的类型(叶、小枝、花果等)、凋落物的分解程度制约(陈莎莎等, 2010).在不同的环境梯度下, 灌丛生态系统生产的凋落物的种类和组成差异明显, 进而造成凋落物在进入地表时碳浓度发生改变.一般而言, 在年平均气温较低时, 植物为响应低温环境会产生较高碳含量的有机物, 而碳元素在器官凋落时一般很少被重新吸收(Vergutz et al., 2012; Schreeg et al., 2013; Sun et al., 2016), 因此导致进入地表前凋落物中碳浓度保持稳定.而进入地表的凋落物随着分解进行, 碳浓度会出现下降.上述过程相互作用, 使得凋落物现存量中碳浓度低于植被活体碳浓度, 从而造成区域尺度上凋落物现存量的碳转换系数低于植被活体. ...
Litter production and organic matter accumulation in exclosures of the Tigray highlands, Ethiopia 4 2006
... 凋落物也称枯落物, 是指由植被地上部分产生并归还到地表的所有有机物质的总称(彭少麟和刘强, 2002; Parsons et al., 2014a; Domke et al., 2016).凋落物的生产、积累以及分解是凋落物研究的重要内容(Descheemaeker et al., 2006; Parsons et al., 2014b).凋落物现存量又称凋落物的累积量, 通常指一定面积上特定生态系统地表堆积的凋落物的质量, 反映植被凋落层在时间和空间上现存凋落物的多少, 是凋落物输入和输出(分解)后的净积累量, 可被看作是不同分解阶段和状态的凋落物的复合体(阎恩荣等, 2008; Hilli et al., 2010; Domke et al., 2016).当凋落物从植物体脱落进入地表时, 就开始了物理、化学和生物的分解过程, 通过这些过程释放出生态系统所需要的养分和CO2, 从而参与陆地生态系统的物质循环和能量流动过程.因此, 准确地评估凋落物现存量对揭示陆地生态系统碳循环具有重要的生态学意义(Descheemaeker et al., 2006; Marty et al., 2015; Domke et al., 2016). ...
... , 从而参与陆地生态系统的物质循环和能量流动过程.因此, 准确地评估凋落物现存量对揭示陆地生态系统碳循环具有重要的生态学意义(Descheemaeker et al., 2006; Marty et al., 2015; Domke et al., 2016). ...
... 目前, 与凋落物的产量和分解的研究(汪思龙和陈楚莹, 2010; 唐仕姗等, 2014; García-Palacios et al., 2016)相比, 对凋落物现存量的研究相对较少, 多局限于单个地点的研究(郑路和卢立华, 2012; Portillo-Estrada et al., 2016), 对大尺度凋落物现存量格局及其气候控制因子的认识仍不清楚.凋落物现存量受凋落物产量和分解量的动态关系决定(Twilley et al., 1986; 马玉珠等, 2013).因此, 影响凋落物产量和分解过程的因素都会对凋落物的现存量产生重要影响(张新平等, 2008; 李强等, 2014), 推测凋落物的现存量同样受到这些因子的制约.然而, 凋落物的产量和分解对不同环境因子的响应存在差异.有研究表明, 凋落物的产量主要受年平均气温的制约(Liu et al., 2004; 吕国红等, 2014), 而凋落物的分解通常与年平均气温、年降水量有关, 其控制因子一般因研究地区而异(赵红梅等, 2012; Zhang & Wang, 2015).因此, 凋落物的现存量是否存在一定的空间分布格局, 目前尚缺乏相应的实证研究(Simmons et al., 1996; Descheemaeker et al., 2006; Westcott et al., 2014). ...
... 陆地生态系统凋落物的现存量受到多种环境因素共同控制, 大尺度上水分和热量通过控制植被类型影响凋落物的产量和分解, 进而影响凋落物的现存量(Zhou et al., 2009; Chave et al., 2010).本研究发现, 中国南方灌丛生态系统的凋落物现存量地理格局受到年平均气温、土壤pH值以及土壤全磷含量等因子的制约, 但最重要的环境因子是年平均气温.这与前人对凋落物产量和分解的研究结果(Desch- eemaeker et al., 2006; Pitkänen et al., 2012)一致.年平均气温是植物生长所需要的基本环境条件, 其年际间变化影响植物的物候变化, 从而制约着凋落物的产量.凋落物现存量受到凋落物生产和分解的制约(Hill et al., 2010).大量研究表明, 凋落物的产量与年平均气温显著正相关(张新平等, 2008; Salinas et al., 2011; 吕国红等, 2014), 不考虑凋落物的分解时, 凋落物的现存量应与年平均气温成正比, 但本研究发现凋落物现存量与年平均气温成反比.因此, 凋落物的分解可能是影响中国南方灌丛凋落物现存量空间分布的关键过程.上述推断与前人关于凋落物分解自然气候梯度的研究(Zhang et al., 2008; Hilli et al., 2010)一致.不同的研究发现, 随着纬度增加, 凋落物的分解速率下降, 温度升高可以显著加快不同生态系统的地表凋落物的分解(Aerts, 1997; Dorrepaal et al., 2005; Zhang & Wang, 2015).例如, Dorrepaal等(2005)研究欧洲西北部的灌木物种分解时发现, 年平均气温是与灌木树种凋落物分解速率最密切的控制因子.同样Hilli等(2010)研究森林凋落物分解对凋落物储量的影响时发现凋落物分解过程是凋落物储量空间变异的最为关键过程.本研究位于热带和亚热带地区, 温度是影响灌丛凋落物产量和分解的限制因子, 对凋落物分解的影响程度更突出, 因此年平均气温对凋落物生产和分解之间作用的差异性是导致中国南方灌丛生态系统凋落物现存量空间分异的主要原因. ...
Estimating litter carbon stocks on forest land in the United States 4 2016
... 凋落物也称枯落物, 是指由植被地上部分产生并归还到地表的所有有机物质的总称(彭少麟和刘强, 2002; Parsons et al., 2014a; Domke et al., 2016).凋落物的生产、积累以及分解是凋落物研究的重要内容(Descheemaeker et al., 2006; Parsons et al., 2014b).凋落物现存量又称凋落物的累积量, 通常指一定面积上特定生态系统地表堆积的凋落物的质量, 反映植被凋落层在时间和空间上现存凋落物的多少, 是凋落物输入和输出(分解)后的净积累量, 可被看作是不同分解阶段和状态的凋落物的复合体(阎恩荣等, 2008; Hilli et al., 2010; Domke et al., 2016).当凋落物从植物体脱落进入地表时, 就开始了物理、化学和生物的分解过程, 通过这些过程释放出生态系统所需要的养分和CO2, 从而参与陆地生态系统的物质循环和能量流动过程.因此, 准确地评估凋落物现存量对揭示陆地生态系统碳循环具有重要的生态学意义(Descheemaeker et al., 2006; Marty et al., 2015; Domke et al., 2016). ...
... ; Domke et al., 2016).当凋落物从植物体脱落进入地表时, 就开始了物理、化学和生物的分解过程, 通过这些过程释放出生态系统所需要的养分和CO2, 从而参与陆地生态系统的物质循环和能量流动过程.因此, 准确地评估凋落物现存量对揭示陆地生态系统碳循环具有重要的生态学意义(Descheemaeker et al., 2006; Marty et al., 2015; Domke et al., 2016). ...
The importance of litter traits and decomposers for litter decomposition: A comparison of aquatic and terrestrial ecosystems within and across biomes 1 2016
... 目前, 与凋落物的产量和分解的研究(汪思龙和陈楚莹, 2010; 唐仕姗等, 2014; García-Palacios et al., 2016)相比, 对凋落物现存量的研究相对较少, 多局限于单个地点的研究(郑路和卢立华, 2012; Portillo-Estrada et al., 2016), 对大尺度凋落物现存量格局及其气候控制因子的认识仍不清楚.凋落物现存量受凋落物产量和分解量的动态关系决定(Twilley et al., 1986; 马玉珠等, 2013).因此, 影响凋落物产量和分解过程的因素都会对凋落物的现存量产生重要影响(张新平等, 2008; 李强等, 2014), 推测凋落物的现存量同样受到这些因子的制约.然而, 凋落物的产量和分解对不同环境因子的响应存在差异.有研究表明, 凋落物的产量主要受年平均气温的制约(Liu et al., 2004; 吕国红等, 2014), 而凋落物的分解通常与年平均气温、年降水量有关, 其控制因子一般因研究地区而异(赵红梅等, 2012; Zhang & Wang, 2015).因此, 凋落物的现存量是否存在一定的空间分布格局, 目前尚缺乏相应的实证研究(Simmons et al., 1996; Descheemaeker et al., 2006; Westcott et al., 2014). ...
Very high resolution interpolated climate surfaces for global land areas 1 2005
Litter decomposition rates in relation to litter stocks in boreal coniferous forests along climatic and soil fertility gradients 3 2010
... 凋落物也称枯落物, 是指由植被地上部分产生并归还到地表的所有有机物质的总称(彭少麟和刘强, 2002; Parsons et al., 2014a; Domke et al., 2016).凋落物的生产、积累以及分解是凋落物研究的重要内容(Descheemaeker et al., 2006; Parsons et al., 2014b).凋落物现存量又称凋落物的累积量, 通常指一定面积上特定生态系统地表堆积的凋落物的质量, 反映植被凋落层在时间和空间上现存凋落物的多少, 是凋落物输入和输出(分解)后的净积累量, 可被看作是不同分解阶段和状态的凋落物的复合体(阎恩荣等, 2008; Hilli et al., 2010; Domke et al., 2016).当凋落物从植物体脱落进入地表时, 就开始了物理、化学和生物的分解过程, 通过这些过程释放出生态系统所需要的养分和CO2, 从而参与陆地生态系统的物质循环和能量流动过程.因此, 准确地评估凋落物现存量对揭示陆地生态系统碳循环具有重要的生态学意义(Descheemaeker et al., 2006; Marty et al., 2015; Domke et al., 2016). ...
... 陆地生态系统凋落物的现存量受到多种环境因素共同控制, 大尺度上水分和热量通过控制植被类型影响凋落物的产量和分解, 进而影响凋落物的现存量(Zhou et al., 2009; Chave et al., 2010).本研究发现, 中国南方灌丛生态系统的凋落物现存量地理格局受到年平均气温、土壤pH值以及土壤全磷含量等因子的制约, 但最重要的环境因子是年平均气温.这与前人对凋落物产量和分解的研究结果(Desch- eemaeker et al., 2006; Pitkänen et al., 2012)一致.年平均气温是植物生长所需要的基本环境条件, 其年际间变化影响植物的物候变化, 从而制约着凋落物的产量.凋落物现存量受到凋落物生产和分解的制约(Hill et al., 2010).大量研究表明, 凋落物的产量与年平均气温显著正相关(张新平等, 2008; Salinas et al., 2011; 吕国红等, 2014), 不考虑凋落物的分解时, 凋落物的现存量应与年平均气温成正比, 但本研究发现凋落物现存量与年平均气温成反比.因此, 凋落物的分解可能是影响中国南方灌丛凋落物现存量空间分布的关键过程.上述推断与前人关于凋落物分解自然气候梯度的研究(Zhang et al., 2008; Hilli et al., 2010)一致.不同的研究发现, 随着纬度增加, 凋落物的分解速率下降, 温度升高可以显著加快不同生态系统的地表凋落物的分解(Aerts, 1997; Dorrepaal et al., 2005; Zhang & Wang, 2015).例如, Dorrepaal等(2005)研究欧洲西北部的灌木物种分解时发现, 年平均气温是与灌木树种凋落物分解速率最密切的控制因子.同样Hilli等(2010)研究森林凋落物分解对凋落物储量的影响时发现凋落物分解过程是凋落物储量空间变异的最为关键过程.本研究位于热带和亚热带地区, 温度是影响灌丛凋落物产量和分解的限制因子, 对凋落物分解的影响程度更突出, 因此年平均气温对凋落物生产和分解之间作用的差异性是导致中国南方灌丛生态系统凋落物现存量空间分异的主要原因. ...
... ; Hilli et al., 2010)一致.不同的研究发现, 随着纬度增加, 凋落物的分解速率下降, 温度升高可以显著加快不同生态系统的地表凋落物的分解(Aerts, 1997; Dorrepaal et al., 2005; Zhang & Wang, 2015).例如, Dorrepaal等(2005)研究欧洲西北部的灌木物种分解时发现, 年平均气温是与灌木树种凋落物分解速率最密切的控制因子.同样Hilli等(2010)研究森林凋落物分解对凋落物储量的影响时发现凋落物分解过程是凋落物储量空间变异的最为关键过程.本研究位于热带和亚热带地区, 温度是影响灌丛凋落物产量和分解的限制因子, 对凋落物分解的影响程度更突出, 因此年平均气温对凋落物生产和分解之间作用的差异性是导致中国南方灌丛生态系统凋落物现存量空间分异的主要原因. ...
... 目前, 与凋落物的产量和分解的研究(汪思龙和陈楚莹, 2010; 唐仕姗等, 2014; García-Palacios et al., 2016)相比, 对凋落物现存量的研究相对较少, 多局限于单个地点的研究(郑路和卢立华, 2012; Portillo-Estrada et al., 2016), 对大尺度凋落物现存量格局及其气候控制因子的认识仍不清楚.凋落物现存量受凋落物产量和分解量的动态关系决定(Twilley et al., 1986; 马玉珠等, 2013).因此, 影响凋落物产量和分解过程的因素都会对凋落物的现存量产生重要影响(张新平等, 2008; 李强等, 2014), 推测凋落物的现存量同样受到这些因子的制约.然而, 凋落物的产量和分解对不同环境因子的响应存在差异.有研究表明, 凋落物的产量主要受年平均气温的制约(Liu et al., 2004; 吕国红等, 2014), 而凋落物的分解通常与年平均气温、年降水量有关, 其控制因子一般因研究地区而异(赵红梅等, 2012; Zhang & Wang, 2015).因此, 凋落物的现存量是否存在一定的空间分布格局, 目前尚缺乏相应的实证研究(Simmons et al., 1996; Descheemaeker et al., 2006; Westcott et al., 2014). ...
... 陆地生态系统凋落物的现存量受到多种环境因素共同控制, 大尺度上水分和热量通过控制植被类型影响凋落物的产量和分解, 进而影响凋落物的现存量(Zhou et al., 2009; Chave et al., 2010).本研究发现, 中国南方灌丛生态系统的凋落物现存量地理格局受到年平均气温、土壤pH值以及土壤全磷含量等因子的制约, 但最重要的环境因子是年平均气温.这与前人对凋落物产量和分解的研究结果(Desch- eemaeker et al., 2006; Pitkänen et al., 2012)一致.年平均气温是植物生长所需要的基本环境条件, 其年际间变化影响植物的物候变化, 从而制约着凋落物的产量.凋落物现存量受到凋落物生产和分解的制约(Hill et al., 2010).大量研究表明, 凋落物的产量与年平均气温显著正相关(张新平等, 2008; Salinas et al., 2011; 吕国红等, 2014), 不考虑凋落物的分解时, 凋落物的现存量应与年平均气温成正比, 但本研究发现凋落物现存量与年平均气温成反比.因此, 凋落物的分解可能是影响中国南方灌丛凋落物现存量空间分布的关键过程.上述推断与前人关于凋落物分解自然气候梯度的研究(Zhang et al., 2008; Hilli et al., 2010)一致.不同的研究发现, 随着纬度增加, 凋落物的分解速率下降, 温度升高可以显著加快不同生态系统的地表凋落物的分解(Aerts, 1997; Dorrepaal et al., 2005; Zhang & Wang, 2015).例如, Dorrepaal等(2005)研究欧洲西北部的灌木物种分解时发现, 年平均气温是与灌木树种凋落物分解速率最密切的控制因子.同样Hilli等(2010)研究森林凋落物分解对凋落物储量的影响时发现凋落物分解过程是凋落物储量空间变异的最为关键过程.本研究位于热带和亚热带地区, 温度是影响灌丛凋落物产量和分解的限制因子, 对凋落物分解的影响程度更突出, 因此年平均气温对凋落物生产和分解之间作用的差异性是导致中国南方灌丛生态系统凋落物现存量空间分异的主要原因. ...
中国森林凋落物不同组分异速比例关系 1 2013
... 目前, 与凋落物的产量和分解的研究(汪思龙和陈楚莹, 2010; 唐仕姗等, 2014; García-Palacios et al., 2016)相比, 对凋落物现存量的研究相对较少, 多局限于单个地点的研究(郑路和卢立华, 2012; Portillo-Estrada et al., 2016), 对大尺度凋落物现存量格局及其气候控制因子的认识仍不清楚.凋落物现存量受凋落物产量和分解量的动态关系决定(Twilley et al., 1986; 马玉珠等, 2013).因此, 影响凋落物产量和分解过程的因素都会对凋落物的现存量产生重要影响(张新平等, 2008; 李强等, 2014), 推测凋落物的现存量同样受到这些因子的制约.然而, 凋落物的产量和分解对不同环境因子的响应存在差异.有研究表明, 凋落物的产量主要受年平均气温的制约(Liu et al., 2004; 吕国红等, 2014), 而凋落物的分解通常与年平均气温、年降水量有关, 其控制因子一般因研究地区而异(赵红梅等, 2012; Zhang & Wang, 2015).因此, 凋落物的现存量是否存在一定的空间分布格局, 目前尚缺乏相应的实证研究(Simmons et al., 1996; Descheemaeker et al., 2006; Westcott et al., 2014). ...
Variation in stocks and distribution of organic C in soils across 21 eastern Canadian temperate and boreal forests 1 2015
... 凋落物也称枯落物, 是指由植被地上部分产生并归还到地表的所有有机物质的总称(彭少麟和刘强, 2002; Parsons et al., 2014a; Domke et al., 2016).凋落物的生产、积累以及分解是凋落物研究的重要内容(Descheemaeker et al., 2006; Parsons et al., 2014b).凋落物现存量又称凋落物的累积量, 通常指一定面积上特定生态系统地表堆积的凋落物的质量, 反映植被凋落层在时间和空间上现存凋落物的多少, 是凋落物输入和输出(分解)后的净积累量, 可被看作是不同分解阶段和状态的凋落物的复合体(阎恩荣等, 2008; Hilli et al., 2010; Domke et al., 2016).当凋落物从植物体脱落进入地表时, 就开始了物理、化学和生物的分解过程, 通过这些过程释放出生态系统所需要的养分和CO2, 从而参与陆地生态系统的物质循环和能量流动过程.因此, 准确地评估凋落物现存量对揭示陆地生态系统碳循环具有重要的生态学意义(Descheemaeker et al., 2006; Marty et al., 2015; Domke et al., 2016). ...
Global litter production, pools, and turnover times: Estimates from measurement data and regression models 1 1997
Determinants of the pathways of litter chemical decomposition in a tropical region 1 2014
... 凋落物也称枯落物, 是指由植被地上部分产生并归还到地表的所有有机物质的总称(彭少麟和刘强, 2002; Parsons et al., 2014a; Domke et al., 2016).凋落物的生产、积累以及分解是凋落物研究的重要内容(Descheemaeker et al., 2006; Parsons et al., 2014b).凋落物现存量又称凋落物的累积量, 通常指一定面积上特定生态系统地表堆积的凋落物的质量, 反映植被凋落层在时间和空间上现存凋落物的多少, 是凋落物输入和输出(分解)后的净积累量, 可被看作是不同分解阶段和状态的凋落物的复合体(阎恩荣等, 2008; Hilli et al., 2010; Domke et al., 2016).当凋落物从植物体脱落进入地表时, 就开始了物理、化学和生物的分解过程, 通过这些过程释放出生态系统所需要的养分和CO2, 从而参与陆地生态系统的物质循环和能量流动过程.因此, 准确地评估凋落物现存量对揭示陆地生态系统碳循环具有重要的生态学意义(Descheemaeker et al., 2006; Marty et al., 2015; Domke et al., 2016). ...
Spatial variability in litterfall, litter standing crop and litter quality in a tropical rain forest region 2 2014
... 凋落物也称枯落物, 是指由植被地上部分产生并归还到地表的所有有机物质的总称(彭少麟和刘强, 2002; Parsons et al., 2014a; Domke et al., 2016).凋落物的生产、积累以及分解是凋落物研究的重要内容(Descheemaeker et al., 2006; Parsons et al., 2014b).凋落物现存量又称凋落物的累积量, 通常指一定面积上特定生态系统地表堆积的凋落物的质量, 反映植被凋落层在时间和空间上现存凋落物的多少, 是凋落物输入和输出(分解)后的净积累量, 可被看作是不同分解阶段和状态的凋落物的复合体(阎恩荣等, 2008; Hilli et al., 2010; Domke et al., 2016).当凋落物从植物体脱落进入地表时, 就开始了物理、化学和生物的分解过程, 通过这些过程释放出生态系统所需要的养分和CO2, 从而参与陆地生态系统的物质循环和能量流动过程.因此, 准确地评估凋落物现存量对揭示陆地生态系统碳循环具有重要的生态学意义(Descheemaeker et al., 2006; Marty et al., 2015; Domke et al., 2016). ...
... 凋落物也称枯落物, 是指由植被地上部分产生并归还到地表的所有有机物质的总称(彭少麟和刘强, 2002; Parsons et al., 2014a; Domke et al., 2016).凋落物的生产、积累以及分解是凋落物研究的重要内容(Descheemaeker et al., 2006; Parsons et al., 2014b).凋落物现存量又称凋落物的累积量, 通常指一定面积上特定生态系统地表堆积的凋落物的质量, 反映植被凋落层在时间和空间上现存凋落物的多少, 是凋落物输入和输出(分解)后的净积累量, 可被看作是不同分解阶段和状态的凋落物的复合体(阎恩荣等, 2008; Hilli et al., 2010; Domke et al., 2016).当凋落物从植物体脱落进入地表时, 就开始了物理、化学和生物的分解过程, 通过这些过程释放出生态系统所需要的养分和CO2, 从而参与陆地生态系统的物质循环和能量流动过程.因此, 准确地评估凋落物现存量对揭示陆地生态系统碳循环具有重要的生态学意义(Descheemaeker et al., 2006; Marty et al., 2015; Domke et al., 2016). ...
The carbon balance of terrestrial ecosystems in China 1 2009
The sensitivity of tropical leaf litter decomposition to temperature: Results from a large-scale leaf translocation experiment along an elevation gradient in Peruvian forests 1 2011
... 陆地生态系统凋落物的现存量受到多种环境因素共同控制, 大尺度上水分和热量通过控制植被类型影响凋落物的产量和分解, 进而影响凋落物的现存量(Zhou et al., 2009; Chave et al., 2010).本研究发现, 中国南方灌丛生态系统的凋落物现存量地理格局受到年平均气温、土壤pH值以及土壤全磷含量等因子的制约, 但最重要的环境因子是年平均气温.这与前人对凋落物产量和分解的研究结果(Desch- eemaeker et al., 2006; Pitkänen et al., 2012)一致.年平均气温是植物生长所需要的基本环境条件, 其年际间变化影响植物的物候变化, 从而制约着凋落物的产量.凋落物现存量受到凋落物生产和分解的制约(Hill et al., 2010).大量研究表明, 凋落物的产量与年平均气温显著正相关(张新平等, 2008; Salinas et al., 2011; 吕国红等, 2014), 不考虑凋落物的分解时, 凋落物的现存量应与年平均气温成正比, 但本研究发现凋落物现存量与年平均气温成反比.因此, 凋落物的分解可能是影响中国南方灌丛凋落物现存量空间分布的关键过程.上述推断与前人关于凋落物分解自然气候梯度的研究(Zhang et al., 2008; Hilli et al., 2010)一致.不同的研究发现, 随着纬度增加, 凋落物的分解速率下降, 温度升高可以显著加快不同生态系统的地表凋落物的分解(Aerts, 1997; Dorrepaal et al., 2005; Zhang & Wang, 2015).例如, Dorrepaal等(2005)研究欧洲西北部的灌木物种分解时发现, 年平均气温是与灌木树种凋落物分解速率最密切的控制因子.同样Hilli等(2010)研究森林凋落物分解对凋落物储量的影响时发现凋落物分解过程是凋落物储量空间变异的最为关键过程.本研究位于热带和亚热带地区, 温度是影响灌丛凋落物产量和分解的限制因子, 对凋落物分解的影响程度更突出, 因此年平均气温对凋落物生产和分解之间作用的差异性是导致中国南方灌丛生态系统凋落物现存量空间分异的主要原因. ...
Nutrient-specific solubility patterns of leaf litter across 41 lowland tropical woody species 1 2013
... 碳转换系数是研究陆地生态系统碳收支的重要参数.本研究发现, 中国南方灌丛凋落物现存量的碳转换系数为0.41, 低于前人研究灌丛生态系统所采用的植被活体碳转换系数(0.50)(胡会峰等, 2006).因此在研究灌丛生态系统碳储量时, 采用常用的植被活体碳转换系数可能会高估灌丛生态系统凋落物碳储量22%.凋落物现存量的碳转换系数与碳浓度相关, 受到环境因子、凋落物的类型(叶、小枝、花果等)、凋落物的分解程度制约(陈莎莎等, 2010).在不同的环境梯度下, 灌丛生态系统生产的凋落物的种类和组成差异明显, 进而造成凋落物在进入地表时碳浓度发生改变.一般而言, 在年平均气温较低时, 植物为响应低温环境会产生较高碳含量的有机物, 而碳元素在器官凋落时一般很少被重新吸收(Vergutz et al., 2012; Schreeg et al., 2013; Sun et al., 2016), 因此导致进入地表前凋落物中碳浓度保持稳定.而进入地表的凋落物随着分解进行, 碳浓度会出现下降.上述过程相互作用, 使得凋落物现存量中碳浓度低于植被活体碳浓度, 从而造成区域尺度上凋落物现存量的碳转换系数低于植被活体. ...
Forest floor carbon pools and fluxes along a regional climate gradient in Maine, USA 1 1996
... 目前, 与凋落物的产量和分解的研究(汪思龙和陈楚莹, 2010; 唐仕姗等, 2014; García-Palacios et al., 2016)相比, 对凋落物现存量的研究相对较少, 多局限于单个地点的研究(郑路和卢立华, 2012; Portillo-Estrada et al., 2016), 对大尺度凋落物现存量格局及其气候控制因子的认识仍不清楚.凋落物现存量受凋落物产量和分解量的动态关系决定(Twilley et al., 1986; 马玉珠等, 2013).因此, 影响凋落物产量和分解过程的因素都会对凋落物的现存量产生重要影响(张新平等, 2008; 李强等, 2014), 推测凋落物的现存量同样受到这些因子的制约.然而, 凋落物的产量和分解对不同环境因子的响应存在差异.有研究表明, 凋落物的产量主要受年平均气温的制约(Liu et al., 2004; 吕国红等, 2014), 而凋落物的分解通常与年平均气温、年降水量有关, 其控制因子一般因研究地区而异(赵红梅等, 2012; Zhang & Wang, 2015).因此, 凋落物的现存量是否存在一定的空间分布格局, 目前尚缺乏相应的实证研究(Simmons et al., 1996; Descheemaeker et al., 2006; Westcott et al., 2014). ...
Biogeographic patterns of nutrient resorption from Quercus variabilis Blume leaves across China 1 2016
... 碳转换系数是研究陆地生态系统碳收支的重要参数.本研究发现, 中国南方灌丛凋落物现存量的碳转换系数为0.41, 低于前人研究灌丛生态系统所采用的植被活体碳转换系数(0.50)(胡会峰等, 2006).因此在研究灌丛生态系统碳储量时, 采用常用的植被活体碳转换系数可能会高估灌丛生态系统凋落物碳储量22%.凋落物现存量的碳转换系数与碳浓度相关, 受到环境因子、凋落物的类型(叶、小枝、花果等)、凋落物的分解程度制约(陈莎莎等, 2010).在不同的环境梯度下, 灌丛生态系统生产的凋落物的种类和组成差异明显, 进而造成凋落物在进入地表时碳浓度发生改变.一般而言, 在年平均气温较低时, 植物为响应低温环境会产生较高碳含量的有机物, 而碳元素在器官凋落时一般很少被重新吸收(Vergutz et al., 2012; Schreeg et al., 2013; Sun et al., 2016), 因此导致进入地表前凋落物中碳浓度保持稳定.而进入地表的凋落物随着分解进行, 碳浓度会出现下降.上述过程相互作用, 使得凋落物现存量中碳浓度低于植被活体碳浓度, 从而造成区域尺度上凋落物现存量的碳转换系数低于植被活体. ...
中国森林生态系统凋落叶分解速率的分布特征及其控制因子 1 2014
... 目前, 与凋落物的产量和分解的研究(汪思龙和陈楚莹, 2010; 唐仕姗等, 2014; García-Palacios et al., 2016)相比, 对凋落物现存量的研究相对较少, 多局限于单个地点的研究(郑路和卢立华, 2012; Portillo-Estrada et al., 2016), 对大尺度凋落物现存量格局及其气候控制因子的认识仍不清楚.凋落物现存量受凋落物产量和分解量的动态关系决定(Twilley et al., 1986; 马玉珠等, 2013).因此, 影响凋落物产量和分解过程的因素都会对凋落物的现存量产生重要影响(张新平等, 2008; 李强等, 2014), 推测凋落物的现存量同样受到这些因子的制约.然而, 凋落物的产量和分解对不同环境因子的响应存在差异.有研究表明, 凋落物的产量主要受年平均气温的制约(Liu et al., 2004; 吕国红等, 2014), 而凋落物的分解通常与年平均气温、年降水量有关, 其控制因子一般因研究地区而异(赵红梅等, 2012; Zhang & Wang, 2015).因此, 凋落物的现存量是否存在一定的空间分布格局, 目前尚缺乏相应的实证研究(Simmons et al., 1996; Descheemaeker et al., 2006; Westcott et al., 2014). ...
... 凋落物也称枯落物, 是指由植被地上部分产生并归还到地表的所有有机物质的总称(彭少麟和刘强, 2002; Parsons et al., 2014a; Domke et al., 2016).凋落物的生产、积累以及分解是凋落物研究的重要内容(Descheemaeker et al., 2006; Parsons et al., 2014b).凋落物现存量又称凋落物的累积量, 通常指一定面积上特定生态系统地表堆积的凋落物的质量, 反映植被凋落层在时间和空间上现存凋落物的多少, 是凋落物输入和输出(分解)后的净积累量, 可被看作是不同分解阶段和状态的凋落物的复合体(阎恩荣等, 2008; Hilli et al., 2010; Domke et al., 2016).当凋落物从植物体脱落进入地表时, 就开始了物理、化学和生物的分解过程, 通过这些过程释放出生态系统所需要的养分和CO2, 从而参与陆地生态系统的物质循环和能量流动过程.因此, 准确地评估凋落物现存量对揭示陆地生态系统碳循环具有重要的生态学意义(Descheemaeker et al., 2006; Marty et al., 2015; Domke et al., 2016). ...
Rates of litter decomposition in terrestrial ecosystems: Global patterns and controlling factors 1 2008
... 陆地生态系统凋落物的现存量受到多种环境因素共同控制, 大尺度上水分和热量通过控制植被类型影响凋落物的产量和分解, 进而影响凋落物的现存量(Zhou et al., 2009; Chave et al., 2010).本研究发现, 中国南方灌丛生态系统的凋落物现存量地理格局受到年平均气温、土壤pH值以及土壤全磷含量等因子的制约, 但最重要的环境因子是年平均气温.这与前人对凋落物产量和分解的研究结果(Desch- eemaeker et al., 2006; Pitkänen et al., 2012)一致.年平均气温是植物生长所需要的基本环境条件, 其年际间变化影响植物的物候变化, 从而制约着凋落物的产量.凋落物现存量受到凋落物生产和分解的制约(Hill et al., 2010).大量研究表明, 凋落物的产量与年平均气温显著正相关(张新平等, 2008; Salinas et al., 2011; 吕国红等, 2014), 不考虑凋落物的分解时, 凋落物的现存量应与年平均气温成正比, 但本研究发现凋落物现存量与年平均气温成反比.因此, 凋落物的分解可能是影响中国南方灌丛凋落物现存量空间分布的关键过程.上述推断与前人关于凋落物分解自然气候梯度的研究(Zhang et al., 2008; Hilli et al., 2010)一致.不同的研究发现, 随着纬度增加, 凋落物的分解速率下降, 温度升高可以显著加快不同生态系统的地表凋落物的分解(Aerts, 1997; Dorrepaal et al., 2005; Zhang & Wang, 2015).例如, Dorrepaal等(2005)研究欧洲西北部的灌木物种分解时发现, 年平均气温是与灌木树种凋落物分解速率最密切的控制因子.同样Hilli等(2010)研究森林凋落物分解对凋落物储量的影响时发现凋落物分解过程是凋落物储量空间变异的最为关键过程.本研究位于热带和亚热带地区, 温度是影响灌丛凋落物产量和分解的限制因子, 对凋落物分解的影响程度更突出, 因此年平均气温对凋落物生产和分解之间作用的差异性是导致中国南方灌丛生态系统凋落物现存量空间分异的主要原因. ...
Control of climate and litter quality on leaf litter decomposition in different climatic zones 3 2015
... 目前, 与凋落物的产量和分解的研究(汪思龙和陈楚莹, 2010; 唐仕姗等, 2014; García-Palacios et al., 2016)相比, 对凋落物现存量的研究相对较少, 多局限于单个地点的研究(郑路和卢立华, 2012; Portillo-Estrada et al., 2016), 对大尺度凋落物现存量格局及其气候控制因子的认识仍不清楚.凋落物现存量受凋落物产量和分解量的动态关系决定(Twilley et al., 1986; 马玉珠等, 2013).因此, 影响凋落物产量和分解过程的因素都会对凋落物的现存量产生重要影响(张新平等, 2008; 李强等, 2014), 推测凋落物的现存量同样受到这些因子的制约.然而, 凋落物的产量和分解对不同环境因子的响应存在差异.有研究表明, 凋落物的产量主要受年平均气温的制约(Liu et al., 2004; 吕国红等, 2014), 而凋落物的分解通常与年平均气温、年降水量有关, 其控制因子一般因研究地区而异(赵红梅等, 2012; Zhang & Wang, 2015).因此, 凋落物的现存量是否存在一定的空间分布格局, 目前尚缺乏相应的实证研究(Simmons et al., 1996; Descheemaeker et al., 2006; Westcott et al., 2014). ...

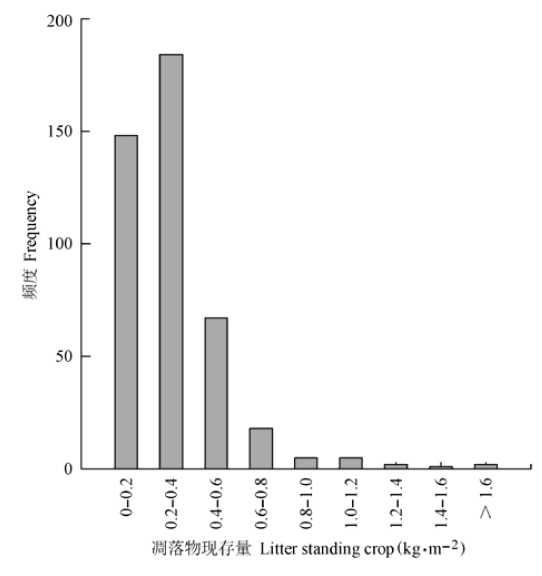 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT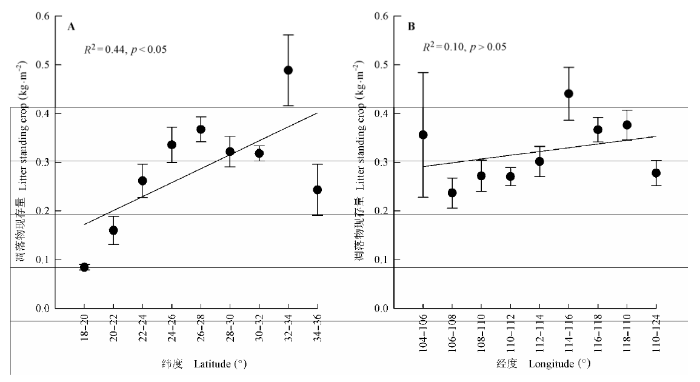 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT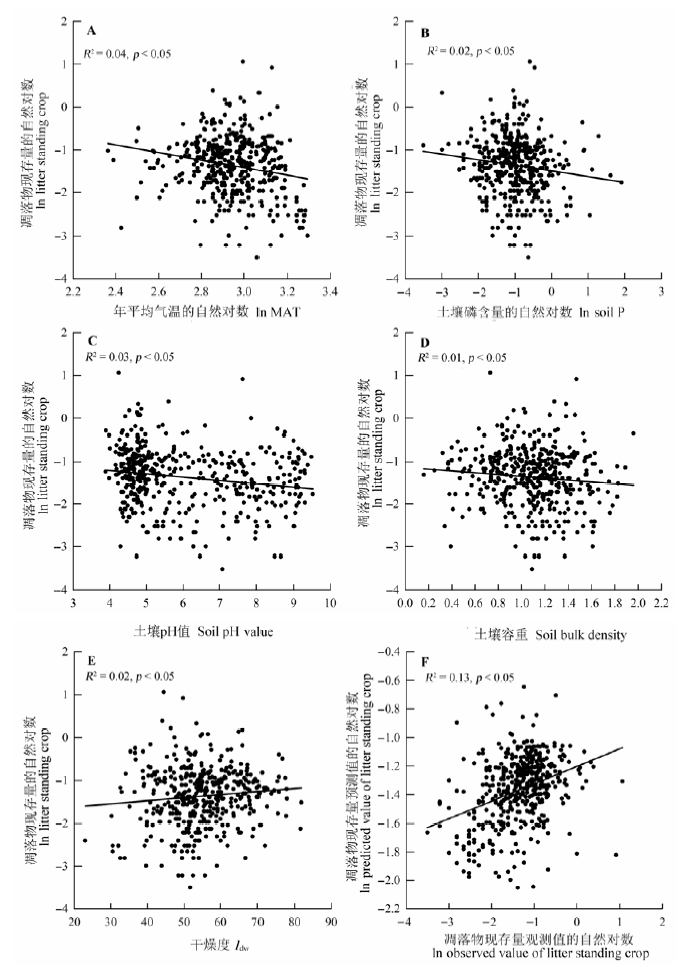 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}