0 引言
【研究意义】蜂王浆是由工蜂咽下腺、上颚腺为主等腺体共同分泌的淡黄色乳浆状物质[1],富含蛋白质和脂肪酸,是蜂王和幼虫的食物,其中所含的脂肪酸在蜂群营养[2]和幼虫发育[3]方面发挥重要作用。其中一种不饱和脂肪酸10-羟基-2癸烯酸(10-HDA)[4],因其在自然界中仅蜂王浆中存在,又被称为“王浆酸”。蜂王浆中10-HDA的含量是评价蜂王浆质量的重要标准和贸易标准之一[5]。蜂王浆同时还是一种对人类健康具有重要作用的天然功能性保健品,具有调节人体血压[6]、提高免疫力[7]、消炎抗菌[8]等功效。20世纪90年代中国从意大利蜜蜂(意蜂,Apis mellifera liguatica,ITBs)中培育出蜂王浆高产蜜蜂(浆蜂,RJBs),浆蜂和意蜂为同一亚种,是意蜂的一个品系[9-10],浆蜂的蜂王浆产量约为意蜂10倍以上,一群浆蜂可年产约10 kg蜂王浆[11]。中国蜂王浆年产量达4 000多吨[12],是世界第一生产大国,浆蜂的选育为中国蜂王浆产业发展发挥巨大贡献。然而在浆蜂蜂王浆高产的同时,10-HDA含量出现一定程度的下降,因此,明确究竟是浆蜂上颚腺生理机能未随着产浆量提高而增强还是其他环境因素所致,对于提高10-HDA含量具有重要意义。【前人研究进展】上颚腺是蜜蜂头部一对与上颚相连的袋状外分泌腺体[13]。蜂王上颚腺分泌物主要是各类信息素,用于蜂群的交流和维护蜂群稳定[14-15]。工蜂上颚腺主要分泌幼虫食物所需脂肪酸[16]和2-庚酮等作为报警信息素[17]。上颚腺基因组学研究表明,与蜂王相比,工蜂上颚腺中脂肪酸合酶(fatty acid synthase,FAS)及fas高表达,表明上颚腺中有关脂肪酸代谢在不同级型间存在差异[18]。经过蜂王浆高产的选育,浆蜂和意蜂的咽下腺[19]、神经系统[20]的蛋白质组等方面已经出现了明显分化,浆蜂咽下腺显著提高了蛋白质合成力,以保证蜂王浆高产的生理需求[21]。神经系统为了保证蜂王浆高产的需求,浆蜂大脑的神经肽加强了对幼虫信息素识别、花粉采集、水代谢平衡的调控能力[20]。这些神经生物学功能的加强在行为和生理上在保证浆蜂蜂王浆的蛋白质种类和含量与意蜂没有差异[21],且比中蜂蜂王浆蛋白的含量还显著提高[22]。上颚腺蛋白质组研究表明,刚出房幼蜂上颚腺表达的蛋白主要是用于腺体的初始发育;哺育蜂时期,上颚腺蛋白质组主要与分泌行为有关,且浆蜂脂肪酸合成能力显著高于意蜂[11];采集蜂时期,上颚腺表达的蛋白可以增强蜜蜂的采集效率和对蜂巢的守卫能力[11]。蛋白质磷酸化作用是一种重要的蛋白质翻译后修饰形式,通常发生在丝氨酸(Ser,S)、苏氨酸(Thr,T)、酪氨酸(Tyr,Y)残基上[23]。蛋白质磷酸化修饰可以调控蛋白功能[24],调控许多生命活动,如细胞周期[25]、生长、发育、凋亡[26]、信号转导[27]等。在蜜蜂中,磷酸化蛋白参与调控多项生命活动,如幼虫生长[28]、抗菌[29]抗病毒[30]以及咽下腺和大脑[31]的生长发育[23]等。研究表明,磷酸化蛋白质组对咽下腺腺体发育以及核糖体活性具有重要的调控作用,从而调节蜂王浆蛋白的合成、翻译过程[23,32]。最新研究表明,哺育蜂和采集蜂大脑蛋白质的磷酸化通过调节蛋白质的功能使其支持不同生理年龄工蜂的神经生理活动,进而调控蜜蜂的哺育和采集行为。蛋白质磷酸化通过调节糖代谢和糖原生成等代谢途径加强哺育蜂的哺育功能,而磷酸化蛋白通过调节光传导通路加强采集蜂的条件采集能力[31]。然而磷酸化蛋白质如何调控上颚腺发育机制及脂肪酸代谢机理尚未开展研究。【本研究切入点】针对蜂王浆产业的理论问题,在磷酸化蛋白质组水平研究上颚腺磷酸化蛋白质对其发育的调控和脂肪酸代谢机理进行研究。【拟解决的关键问题】阐明浆蜂上颚腺磷酸化蛋白质组调控其腺体发育和脂肪酸代谢机理,为蜂王浆优质高产提供理论依据和生产实践基础。1 材料与方法
试验于2016年9月至2017年5月在中国农业科学院蜜蜂研究所完成。1.1 化学试剂
IMAC磷酸化肽段富集材料购自中国科学院大连化学物理研究所;尿素购自Solarbio;Ti(SO4)2(硫酸钛)、硫脲、CHAPS、Tris碱、DTT(二硫苏糖醇)购自Amresco;蛋白酶抑制剂购自Roche,Basel;丙酮和TFA(三氟乙酸)购自J.T.Baker;Bradford工作液购自普利莱;IAA(碘乙酰胺)来自Merk公司;ACN(乙腈)购自Fisher;Trypsin酶购自Promega;甲酸购自MREDA Technology。1.2 蛋白样品制备
1.2.1 IMAC材料制备 称取GTP微球200 mg于烧杯中,加入300 mL超纯水,超声混匀;按照GTP微球:Ti(SO4)2=1:100 称量硫酸钛20 g,缓慢加入烧杯中并搅拌,待其完全溶解放入磁铁转子;将烧杯置于磁力搅拌器上,室温搅拌过夜;将搅拌好的材料分装到50 mL离心管中,用少量超纯水清洗烧瓶加入离心管中,配平,20 000 r/min,离心10 min,弃上清;超纯水洗涤离心后材料,20 000 r/min,离心10 min,洗涤6次,尽量将非特异性吸附钛离子洗净;用200 mmol·L-1 NaCl溶液洗涤上述材料,20 000 r/min,离心10 min,洗两次;再用超纯水洗涤上述材料,20 000 r/min,离心10 min,洗2—3次,保留沉淀;用30 mL超纯水悬浮材料,分装至30个1.5 mL离心管中,每管1 mL,12 500 r/min,离心15 min,弃上清,旋转蒸发仪干燥。后储存于4℃备用。1.2.2 样品准备 试验所用浆蜂和意蜂均饲养于中国农业科学院蜜蜂研究所试验蜂场,浆蜂来自中国浙江省,意蜂来自意大利。分别选取群势相同的5箱浆蜂和意蜂,选取即将羽化出房的子脾置于培养箱中,第2天取一部分新羽化蜂用作出房蜂样品,其余用油漆笔在其胸部背板进行标记,后放回原箱中。待第10天取出用作哺育蜂样品。在蜂箱门口抓取带有花粉团的归巢蜜蜂用作采集蜂样品。用体式显微镜解剖蜜蜂头部,取得上颚腺样品。样品取自5个蜂群,每群取300只,然后将5群样品合并,存于-80℃备用。
1.2.3 蛋白提取 蛋白提取按照HAN等[30]的方法,每30 mg上颚腺样品加入300 μL样品裂解液(8 mol·L-1尿素,2 mol·L-1硫脲,4% CHAPS,20 mmol·L-1 Tris-碱,30 mmol·L-1二硫苏糖醇(DTT),1 mg/10 μL)及蛋白酶抑制剂,冰浴充分研磨,超声,使样品完全溶解。4℃,15 000×g,离心20 min。避开脂肪层取上清液,加入3倍体积预冷的丙酮溶液,冰浴沉淀30 min。4℃,15 000×g,离心20 min。弃上清。开口2—3 min让丙酮挥发,后将沉淀重新溶解于100 μL的5 mol·L-1尿素中,再加入400 μL(4倍体积)40 mmol·L-1 NH4HCO3。使用Bradford方法测定最终的蛋白浓度。
1.2.4 液内酶切 加入50 μL(1/10总体积)100 mmol·L-1 DTT,4℃放置1 h,加入250 μL(5倍DTT体积)100 mmol·L-1 IAA,避光放置1 h。按照酶:蛋白=1:50(W/W)比例加入Trypsin胰蛋白酶,37℃下进行消化反应24 h,后加入1 μL甲酸终止酶切。4℃,14 000×g,离心15 min,取500 μL上清液转移至新管。
1.2.5 磷酸化肽段富集 用500 μL的binding buffer(80% ACN & 6% TFA水溶液)重悬一管IMAC材料;在1.2.4上清液中加入重悬的IMAC材料,30 μL TFA;振荡2 h,14 000×g离心10 min,弃上清;加入1 mL washing buffer 1(50% ACN & 6% TFA & 200 mmol·L-1 NaCl水溶液),振荡30 min,14 000×g,离心10 min,弃上清;加入1 mL washing buffer 2(30% ACN & 0.1% TFA水溶液),振荡30 min,14 000×g,离心10 min,弃上清;加入100 μL elution buffer(500 mmol·L-1 K2HPO4,pH 7),振荡30 min,14 000×g,离心10 min,取上清至新管,重复上述操作。
1.2.6 Zip-tip C18除盐 活化:在50% ACN,0.3% TFA水溶液中反复吹打10次,活化C18柱子;平衡:在equilibrium buffer(0.1% TFA水溶液)中反复吹打10次,平衡C18柱子;富集:用Zip-tip反复吹打样品10次(避免产生气泡);洗脱:取30 μL elution buffer(80% ACN,0.1% TFA水溶液)至新管,用Zip-tip反复吹打10次,将肽段洗脱;用水吹打3—5次,洗去elution buffer,重复富集、洗脱及用水吹打步骤5次左右;干燥:干燥上步中elution buffer。
1.3 质谱分析
用100 μL 0.1%甲酸溶解干燥样品,4℃,14 000×g离心15 min,取50 μL上清于上样管中,同时避免有气泡;将上样管放入色谱仪样品槽,每针上样体积8 μL,每个样品做3针重复。采用纳升级液相色谱系统EASY-nLC 1000(Thermo Fisher Scientific)通过纳升电喷雾源与质谱Q-Exactive(Thermo Fisher Scientific)串联,上样流速5 μL·min-1。流动相A(0.1%甲酸),流动相B(0.1%甲酸 & 80% ACN);通过纳升ESI源将洗脱的肽段注入质谱仪。以数据依赖模式收集离子信号,参数设置如下:母离子扫描分辨率为70 000,400 m/z,荷质比范围:300—1 800 m/z,对丰度最高前20个母离子碎片离子通过高能碰撞诱导解离模式,MS/MS扫描分辨率为17 500,碰撞能:27,动态剔除(带电荷为1或者>8的剔除;动态剔除:10 s)。通过Xcalibur软件(版本2.2,Thermo Fisher Scientific)收集MS/MS数据并保存为Raw文件。1.4 数据处理与分析
1.4.1 数据质量评估 利用MaxQuant 1.5.8.3对质谱数据进行蛋白质定量,Andromeda用于数据库搜索[33]:采用的是蜜蜂数据库(2017年2月从NCBI下载,22 473条目并包含13个常规污染库)。参数设置如下:“Group-specific parameters”选项卡中“Digestion”(酶特异性)选择“Tryspin/P”;“Modifications”(可变修饰)选择“Oxixidation(M)、Acetyl(Protein N-term)、Phospho(STY)”;“Label-free quantification”(非标记定量)选择“LFQ”;“Global parameters”选项卡中“adv. identification”勾选“Match between runs”;“Protein quantification”中“Label min. ratio count”选择“1”;“Label free quantification”勾选“Separate LFQ in parameter groups”。1.4.2 PEAKS 采用PEAKS 7.5软件进行数据库搜索,数据库与MaxQuant所用数据库相同。参数设置如下:先进行从头测序(De novo)计算,Enzyme选择Tryspin(TPCK修饰的胰蛋白酶);fixed modifications(固定修饰)选择Carbamidomethyl(氨甲酰甲基);variable modifications(可变修饰)选择Oxidation(M)和 Phosphorylation (STY);母离子质量数误差范围 (Precursor mass)为30.0 ppm;碎片离子误差范围(Fragment ion)为0.05 Da;每条肽段最多允许有两个漏切位点(Maximum missed cleavages per peptide:2);每个肽段最多允许有3种翻译后修饰(Maximum allowed variable PTM per peptide);搜库完成后,采用假阳性率(FDR)≤1.0%及鉴定到的蛋白中特有肽段(unique peptide)≥1这两个条件对搜库结果进行筛选。运用PEAKS Q板块对搜库结果进行定量,参数如下:保留时间(retention time shift tolerance):0.5 min;质量误差范围(mass error tolerance):30 ppm;特有肽段(unique peptide)≥1;电荷范围(charge between):2—8;蛋白和肽段都是差异倍数(fold change)≥1.5,P≤0.05(significant≥13)。
1.5 生物信息学分析
1.5.1 Perseus分析 将MaxQuant搜库结果导入Perseus软件进行数据质量评估,根据肽段丰度进行聚类定量分析、主成分分析。1.5.2 磷酸化肽段motif分析 将PEAKS搜库结果导入Scaffold PTM软件确定磷酸化位点,以蜜蜂数据库(2017年2月从NCBI下载,22 460条目)为背景,预测磷酸化肽段的基序类型。
2 结果
2.1 数据质量评估
通过Perseus软件对数据进行质量评估,从多聚散点图(图1)可以看出,各样品间重复性较好,数据分布较集中;同一蜂种同一样品间皮尔森相关系数在0.85—0.96,样品重复性较好;但不同蜂种不同样品间相关系数在0.57—0.88,重复性较差。这表明数据质量较好。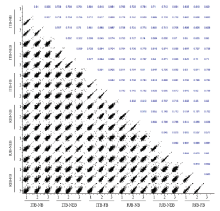 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1浆蜂和意蜂磷酸化蛋白质组数据相关性分布散点图数字代表皮尔森相关系数The numbers indicated the value of Pearson correlation coefficientsRJB-FB:浆蜂-采集蜂High royal jelly bee-forager bee;RJB-NEB:浆蜂-出房蜂High royal jelly bee-newly emerged bee;RJB-NB:浆蜂-哺育蜂High royal jelly bee-nurse bee;ITB-FB:意蜂-采集蜂Italian bee-forager bee;ITB-NEB:意蜂-出房蜂Italian bee-newly emerged bee;ITB-NB:意蜂-哺育蜂Italian bee-nurse bee。下同 The same as below
-->Fig. 1Multi scatter plot of phosphoproteome in high royal jelly bees (RJBs) and Italian bees (ITBs)
-->
2.2 浆蜂、意蜂上颚腺不同发育时期磷酸化蛋白质、肽段、位点鉴定
对浆蜂和意蜂出房蜂、哺育蜂和采集蜂上颚腺进行磷酸化蛋白质组分别进行分析。浆蜂各时期的磷酸化蛋白、肽段、位点数均多于意蜂,分别鉴定到2 225、1 922、2 159、1 740、1 592、1 682个磷酸化蛋白的4 799、3 882、4 808、3 593、2 719、3 679条肽段上658、646、517、700、750、639个化位点发生磷酸修饰(图2-B—D)。其中出房蜂时期浆蜂和意蜂共有磷酸化蛋白最多,为1 468个,分别占意蜂、浆蜂出房蜂蛋白总数84%、66%;哺育蜂时期,共有蛋白最少,为1 144个,分别占意蜂、浆蜂出房蜂蛋白总数71.8%、59.5%;采集蜂时期,共有蛋白1 289个,分别占意蜂、浆蜂出房蜂蛋白总数76.6%、59.7%(图2-A)。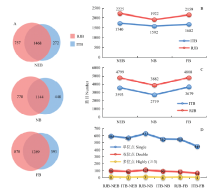 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2浆蜂和意蜂上颚腺磷酸化蛋白、肽段、位点数目A—D分别为浆蜂和意蜂不同时期共有蛋白、磷酸化蛋白、肽段、位点数目
-->Fig. 2The number of phosphoproteins, phosphopeptides and sites identified in mandibular glands of RJBs and ITBs A-D were the numbers of shared proteins, phosphoproteins, phosphopeptides and phosphosites of RJBs and ITBs at different stages, respectively
-->
2.3 浆蜂、意蜂不同时期上颚腺磷酸化蛋白主成分分析
对浆蜂和意蜂不同时期上颚腺蛋白质组做PCA分析,出房蜂蛋白质组彼此最相近,采集蜂次之,而浆蜂和意蜂哺育蜂距离最远,磷酸化蛋白质组差异最大,主成分1、2分别占变异性的20.7%、15.3%(图3)。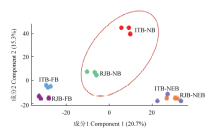 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3浆蜂和意蜂不同时期磷酸化蛋白质组主成分分析主成分分析基于成分1、2
-->Fig. 3The principal component analysis of phosphoproteome of mandibular glands at different stages of RJBs and ITBs The principal component analysis based on component 1 and component 2
-->
2.4 浆蜂、意蜂motif分析
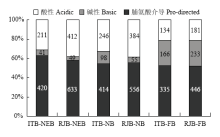
浆蜂、意蜂各时期磷酸化肽段均富集到3种motif(图4):酸性、碱性、脯氨酸介导的motif,但不同蜂种间各时期motif数量不同。浆蜂、意蜂的出房蜂、哺育蜂时期motif数量:脯氨酸介导>酸性>碱性;采集蜂时期脯氨酸介导>碱性>酸性。意蜂脯氨酸介导的motif数量下降,碱性motif上升,酸性motif先上升后显著下降;浆蜂酸性、脯氨酸介导的motif数量下降,碱性motif数量上升。对浆蜂、意蜂不同时期各类motif识别到的激酶家族进行分析(表1),脯氨酸介导的motif主要识别到4类激酶家族:WW domain binding、GSK-3, ERK1, ERK2, CDK5 substrate、GSK3, Erk1, Erk2 and CDK5 kinase和Casein kinase I substrate,其中Casein kinase I substrate激酶家族为浆蜂出房蜂时期特异识别到的。酸性的motif 主要识别到2类激酶家族:Pyruvate dehydrogenase kinase substrate、Casein kinase II substrate,其中Pyruvate dehydrogenase kinase substrate激酶家族为哺育蜂时期特异识别到的。碱性的motif主要识别到2类激酶家族:PKC epsilon kinase substrate、14-3-3 domain binding。其中14-3-3 domain binding激酶家族为采集蜂时期特异识别到的。哺育蜂时期,浆蜂较意蜂特异识别到GSK3, Erk1, Erk2 and CDK5 kinase家族,并且Casein kinase II substrate、Pyruvate dehydrogenase kinase substrate家族(酸性motif)数目要高于意蜂;意蜂PKC epsilon kinase substrate家族(碱性motif)数目高于浆蜂。Table 1
表1
表1浆蜂和意蜂各时期激酶家族
Table 1Kinase families of each stage of RJBs and ITBs
| 酸性Acidic | 碱性Basic | 脯氨酸介导Pro-directed | |
|---|---|---|---|
| ITB-NEB | Casein kinase II substrate | PKC epsilon kinase substrate | WW domain binding; GSK-3, ERK1, ERK2, CDK5 substrate; GSK3, Erk1, Erk2 and CDK5 kinase |
| RJB-NEB | Casein kinase II substrate | PKC epsilon kinase substrate | WW domain binding; GSK-3, ERK1, ERK2, CDK5 substrate; GSK3, Erk1, Erk2 and CDK5 kinase; Casein kinase I substrate |
| ITB-NB | Casein kinase II substrate; Pyruvate dehydrogenase kinase substrate | PKC epsilon kinase substrate | WW domain binding; GSK-3, ERK1, ERK2, CDK5 substrate |
| RJB-NB | Casein kinase II substrate; Pyruvate dehydrogenase kinase substrate | PKC epsilon kinase substrate | WW domain binding; GSK-3, ERK1, ERK2, CDK5 substrate; GSK3, Erk1, Erk2 and CDK5 kinase |
| ITB-FB | Casein kinase II substrate | PKC epsilon kinase substrate; 14-3-3 domain binding | WW domain binding; GSK-3, ERK1, ERK2, CDK5 substrate |
| RJB-FB | Casein kinase II substrate | PKC epsilon kinase substrate; 14-3-3 domain binding | WW domain binding; GSK-3, ERK1, ERK2, CDK5 substrate; GSK3, Erk1, Erk2 and CDK5 kinase |
新窗口打开
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4浆蜂和意蜂不同发育时期上颚腺磷酸化肽段富集motif分布图
-->Fig. 4Motif distribution extracted from phosphopeptides in mandibular glands at different stages of RJBs and ITBs
-->
2.5 差异蛋白定量分析
对上颚腺全蛋白质组与磷酸化蛋白质组进行聚类比较分析(图5-A、5-B),可以看出浆蜂和意蜂各时期蛋白均各自聚类到同一组,相同的表达谱说明蛋白质组和磷酸化蛋白质组在2个不同层面上以类似的表达模式调控腺体的发育和功能。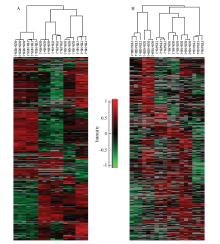 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5浆蜂和意蜂不同发育时期上颚腺差异表达全蛋白和磷酸化蛋白聚类分析A:上颚腺差异表达全蛋白;B:上颚腺差异表达磷酸化蛋白。每个样品有3个重复。纵向代表浆蜂和意蜂不同时期差异表达蛋白。横向代表不同蛋白。红色和绿色分别代表上调表达和下调表达蛋白质
-->Fig. 5Hierarchical clustering analysis of differentially expressed whole proteins & phosphoproteins at different stages of RJBs and ITBs A: Differentially expressed whole proteins; B: Differentially expressed phosphoproteins. Each sample had 3 replications. The columns represented differential proteins at different stages in RJBs and ITBs. The rows represented the individual protein. The up- or down-regulated proteins were labelled in red or green, respectively
-->
出房蜂时期,浆蜂特有蛋白主要富集到胞内信号转导、磷酸酶调节活性、蛋白激酶活性、RNA结合进程(图6-A),意蜂特有蛋白主要富集到蛋白质磷酸化作用进程(图6-B)。哺育蜂时期共鉴定到128个差异表达的蛋白,其中浆蜂高表达87个蛋白,意蜂有41个高表达蛋白(表2),其中多为能量代谢相关蛋白。意蜂高表达蛋白还参与蛋白质合成,浆蜂还包括部分与脂肪酸代谢相关蛋白:烯脂酰-辅酶A水合酶(probable enoyl-CoA hydratase,gi号328778689)、酰基辅酶A结合蛋白(acyl-CoA-binding domain- containing protein 5 isoform X2,gi号571571420)、乙酰辅酶A合成酶(acyl-CoA synthetase family member 2 mitochondrial precursor,gi号332801003)、过氧化物酶(peroxiredoxin 1,gi号328777120)、细胞色素P450(probable cytochrome P450 6a14,gi号571568474)。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图6浆蜂和意蜂出房蜂特有蛋白功能比较A、B分别代表浆蜂、意蜂出房蜂特有蛋白富集的功能类
-->Fig. 6Comparison of enriched functional categories of newly emerged bees unique proteins of RJBs and ITBs A and B were functional categories enriched between unique proteins in RJB & ITBs’ newly emerged bees, respectively
-->
Table 2
表2
表2浆蜂和意蜂哺育蜂时期差异表达磷酸化蛋白
Table 2Differentially expressed phosphoproteins at nurse bee stage of RJBs and ITBs
| 蜂种 Breed | 蛋白名称 Protein name | 登录号 Accession number | 表达量变化倍数 Fold change |
|---|---|---|---|
| ITB | Major royal jelly protein 3-like partial | gi|1031997018 | 4.08 |
| ITB | Troponin T skeletal muscle isoform X2 | gi|571532274 | 3.92 |
| ITB | Cytochrome P450 9e2-like isoform X4 | gi|571570218 | 2.94 |
| ITB | High mobility group protein I | gi|1032011809 | 2.69 |
| ITB | Uncharacterized protein LOC550684 | gi|936975506 | 2.54 |
| ITB | ATP-dependent RNA helicase DHX8 isoform X2 | gi|571577563 | 2.46 |
| ITB | Calcium-transporting ATPase sarcoplasmic/ endoplasmic reticulum type isoform X3 | gi|1032007412 | 2.38 |
| ITB | Centrosome-associated zinc finger protein CP190 | gi|1032006431 | 2.33 |
| ITB | Histone H1-like | gi|328791749 | 2.33 |
| ITB | Mitoguardin | gi|1032018083 | 2.30 |
| ITB | Mitochondrial import receptor subunit TOM70 | gi|328782218 | 2.23 |
| ITB | Hrp65 protein-like isoform X1 | gi|1032001022 | 2.19 |
| ITB | UTP--glucose-1-phosphate uridylyltransferase isoform X1 | gi|571546151 | 2.13 |
| ITB | Basigin isoform X2 | gi|1031999962 | 2.09 |
| ITB | U4/U6.U5 tri-snRNP-associated protein 1 | gi|66513420 | 2.09 |
| ITB | SAFB-like transcription modulator | gi|571525142 | 2.08 |
| ITB | Major royal jelly protein 2 precursor | gi|58585108 | 2.05 |
| ITB | RNA-binding protein 28 | gi|571533499 | 1.97 |
| ITB | Transitional endoplasmic reticulum ATPase TER94 | gi|571575247 | 1.89 |
| ITB | Collagen alpha-1 (IV) chain | gi|328782977 | 1.89 |
| ITB | Zinc finger protein on ecdysone puffs isoform X2 | gi|571499485 | 1.88 |
| ITB | Endophilin-A | gi|571500338 | 1.87 |
| ITB | Golgi integral membrane protein 4-like isoform X1 | gi|1032027113 | 1.87 |
| ITB | LOW QUALITY PROTEIN: microtubule- associated protein tau-like | gi|1032007664 | 1.86 |
| ITB | NADP-dependent malic enzyme isoform X2 | gi|571563710 | 1.81 |
| ITB | Protein LSM14 homolog A isoform X1 | gi|328779321 | 1.74 |
| ITB | Chascon-like | gi|522838229 | 1.73 |
| ITB | Uncharacterized protein DDB_G0287625-like | gi|328778208 | 1.72 |
| ITB | La protein homolog | gi|110759433 | 1.70 |
| ITB | Protein disulfide-isomerase A6 | gi|66551889 | 1.68 |
| ITB | Neurofilament heavy polypeptide | gi|110755329 | 1.67 |
| ITB | Uncharacterized protein LOC100577504 | gi|571556413 | 1.66 |
| ITB | Soluble scavenger receptor cysteine-rich domain containing protein SSC5D | gi|571576399 | 1.65 |
| ITB | RNA-binding protein 26 | gi|571563867 | 1.62 |
| ITB | Probable protein phosphatase CG10417 | gi|571521091 | 1.61 |
| ITB | Pinin | gi|571553836 | 1.60 |
| ITB | Dentin matrix acidic phosphoprotein 1-like | gi|328789423 | 1.57 |
| ITB | Serine/threonine-protein kinase VRK1 | gi|1032021963 | 1.56 |
| ITB | SPARC | gi|571538912 | 1.54 |
| ITB | Inner nuclear membrane protein Man1 | gi|1032001166 | 1.53 |
| ITB | Multidrug resistance-associated protein 4-like | gi|1032005922 | 1.50 |
| RJB | Septin-1 isoform X1 | gi|328787592 | 1.56 |
| RJB | Mitochondrial fission regulator 2-like | gi|571510158 | 1.59 |
| 蜂种 Breed | 蛋白名称 Protein name | 登录号 Accession number | 表达量变化倍数 Fold change |
| RJB | Hyccin isoform X1 | gi|571505174 | 1.64 |
| RJB | Trichohyalin isoform X1 | gi|571569800 | 1.67 |
| RJB | Enhancer of mRNA-decapping protein 4 | gi|66500463 | 1.69 |
| RJB | LOW QUALITY PROTEIN: nucleosome assembly protein 1-like 1 | gi|1032024601 | 1.72 |
| RJB | Glucosylceramidase-like isoform X2 | gi|571535945 | 1.72 |
| RJB | Acyl-CoA-binding domain-containing protein 5 isoform X2 | gi|571571420 | 1.75 |
| RJB | LOW QUALITY PROTEIN: twitchin | gi|1032015155 | 1.79 |
| RJB | Heterogeneous nuclear ribonucleoprotein R isoform X12 | gi|1032019189 | 1.82 |
| RJB | Lysozyme isoform X2 | gi|571576788 | 1.89 |
| RJB | Cytoplasmic dynein 1 light intermediate chain 2 isoform X1 | gi|328780822 | 2.00 |
| RJB | NSFL1 cofactor p47-like | gi|571527109 | 2.00 |
| RJB | Trithorax group protein osa isoform X6 | gi|328787106 | 2.00 |
| RJB | Protein lethal(2)essential for life-like | gi|110750764 | 2.08 |
| RJB | Mitogen-activated protein kinase-binding protein 1 isoform X8 | gi|571575649 | 2.08 |
| RJB | Nascent polypeptide-associated complex subunit alpha isoform X2 | gi|571507498 | 2.08 |
| RJB | Protein sel-1 homolog 1-like | gi|328793806 | 2.08 |
| RJB | Aldehyde dehydrogenase mitochondrial | gi|66530423 | 2.08 |
| RJB | WASH complex subunit FAM21-like | gi|1032006927 | 2.13 |
| RJB | LOW QUALITY PROTEIN: serine/arginine repetitive matrix protein 1 | gi|1032002043 | 2.17 |
| RJB | Uncharacterized protein LOC725017 | gi|1032022797 | 2.17 |
| RJB | Cytosolic non-specific dipeptidase | gi|66535742 | 2.22 |
| RJB | U3 small nucleolar ribonucleoprotein protein MPP10 | gi|571537696 | 2.17 |
| RJB | Uncharacterized protein LOC413556 | gi|1032026621 | 2.27 |
| RJB | Uncharacterized protein LOC100578680 | gi|1031994790 | 2.27 |
| RJB | Acyl-CoA synthetase family member 2 mitochondrial precursor | gi|332801003 | 2.33 |
| RJB | Forkhead box protein O isoform X2 | gi|1032025913 | 2.33 |
| RJB | Uncharacterized protein LOC408677 isoform X6 | gi|571515004 | 2.38 |
| RJB | Prosaposin isoform X2 | gi|571523747 | 2.38 |
| RJB | Uncharacterized protein LOC724121 isoform X3 | gi|328779496 | 2.38 |
| RJB | Glyceraldehyde-3-phosphate dehydrogenase 2 | gi|48142692 | 2.38 |
| RJB | Cyclin-Y | gi|328789216 | 2.44 |
| RJB | Peroxiredoxin 1 | gi|328777120 | 2.50 |
| RJB | Uncharacterized protein LOC410793 | gi|571514351 | 2.50 |
| RJB | ADP/ATP translocase | gi|58531215 | 2.56 |
| RJB | Uncharacterized protein LOC726616 | gi|110763674 | 2.56 |
| RJB | Probable enoyl-CoA hydratase mitochondrial | gi|328778689 | 2.56 |
| RJB | Protein Gawky | gi|1032011880 | 2.70 |
| RJB | Uncharacterized protein LOC725662 | gi|1032013011 | 2.78 |
| RJB | Active breakpoint cluster region-related protein isoform X2 | gi|571519248 | 2.78 |
| RJB | Protein purity of essence isoform X4 | gi|571578252 | 2.94 |
| RJB | Actin clone 205-like | gi|1032021252 | 3.03 |
| RJB | Kinesin heavy chain | gi|66520179 | 3.23 |
| RJB | Transmembrane protein C9orf91 homolog isoform X1 | gi|571552777 | 3.33 |
| RJB | Superoxide dismutase 1 | gi|295849268 | 3.33 |
| 蜂种 Breed | 蛋白名称 Protein name | 登录号 Accession number | 表达量变化倍数 Fold change |
| RJB | JNK-interacting protein 3 isoform X5 | gi|571534840 | 3.33 |
| RJB | MATH and LRR domain-containing protein PFE0570w -like isoform X3 | gi|571499564 | 3.45 |
| RJB | Prostaglandin E synthase 3 | gi|571575529 | 3.85 |
| RJB | Uncharacterized protein LOC409039 isoform X2 | gi|571511889 | 3.85 |
| RJB | Myosin heavy chain 95F | gi|1032007914 | 4.00 |
| RJB | Zinc finger matrin-type protein CG9776-like isoform X2 | gi|571562328 | 4.00 |
| RJB | D-galactonate transporter | gi|66559223 | 4.00 |
| RJB | Uncharacterized protein DDB_G0283697 isoform X2 | gi|1032023143 | 4.00 |
| RJB | Actin related protein 1 | gi|297591985 | 4.35 |
| RJB | Coiled-coil domain-containing protein 6 | gi|66516850 | 4.35 |
| RJB | Cuticular protein 22 precursor | gi|399154150 | 4.55 |
| RJB | Nuclear export mediator factor NEMF homolog isoform X1 | gi|328781799 | 4.55 |
| RJB | PERQ amino acid-rich with GYF domain- containing protein CG11148 isoform X2 | gi|571545516 | 4.55 |
| RJB | Hsp90 co-chaperone Cdc37 | gi|335892823 | 4.76 |
| RJB | 60S ribosomal protein L24 isoform X1 | gi|110761627 | 4.76 |
| RJB | Double-stranded RNA-binding protein Staufen homolog 2 isoform X3 | gi|571553410 | 5.00 |
| RJB | Neuroglian isoform X1 | gi|328788305 | 5.00 |
| RJB | Uncharacterized protein LOC409784 isoform X3 | gi|571576965 | 5.26 |
| RJB | Transaldolase | gi|571501685 | 5.26 |
| RJB | 4-coumarate--CoA ligase 1-like isoform X1 | gi|1032027666 | 5.88 |
| RJB | Probable cytochrome P450 6a14 | gi|571568474 | 5.88 |
| RJB | Trehalase isoform X5 | gi|1032014256 | 6.25 |
| RJB | 40S ribosomal protein S12 | gi|571532294 | 6.67 |
| RJB | Myosin light chain alkali isoform X2 | gi|328777788 | 6.67 |
| RJB | Ras GTPase-activating protein-binding protein 2 | gi|328782334 | 7.14 |
| RJB | Guanine nucleotide-binding protein subunit beta-like protein | gi|48104663 | 7.14 |
| RJB | Serine/threonine-protein kinase PRP4 homolog | gi|1032025937 | 7.69 |
| RJB | Uncharacterized protein LOC100577669 isoform X2 | gi|1032023661 | 10.00 |
| RJB | Protein CREG1 | gi|66521538 | 11.11 |
| RJB | Forkhead box protein K1 isoform X3 | gi|571578374 | 11.11 |
| RJB | Lamin Dm0 isoform X2 | gi|328785544 | 20.00 |
| RJB | Uncharacterized protein LOC409057 isoform X4 | gi|571552460 | 20.00 |
| RJB | Apolipophorin-III-like protein precursor | gi|166795901 | 100.00 |
| RJB | Heat shock protein beta-1 isoform X2 | gi|328791482 | 100.00 |
| RJB | Casein kinase II subunit alpha isoform X2 | gi|571562757 | 3.85 |
| RJB | TPPP family protein CG45057 | gi|571574169 | Unique |
| RJB | Ubiquilin-1 | gi|571533610 | Unique |
| RJB | Serine/threonine-protein kinase MARK2 isoform X12 | gi|571505865 | Unique |
| RJB | POU domain protein CF1A | gi|66517328 | Unique |
| RJB | Serine-rich adhesin for platelets-like isoform X1 | gi|1032023709 | Unique |
| RJB | Large proline-rich protein BAG6 isoform X5 | gi|571534961 | Unique |
新窗口打开
3 讨论
蛋白质磷酸化修饰是一种重要的翻译后修饰方式,通过调控细胞讯号传导[27]进而调控蛋白质功能[24]。虽经蜂王浆高产选育,浆蜂和意蜂上颚腺在不同发育阶段表达的磷酸化蛋白都用于支持腺体的发育和分泌功能,即幼蜂上颚腺磷酸化蛋白质组主要支持腺体发育,哺育和采集蜂的磷酸化蛋白质组主要帮助腺体实现分泌功能。尽管不同发育时期磷酸化蛋白质组表达谱基本一致,但无论是磷酸化蛋白的数目还是肽段、位点的数目,浆蜂均多于意蜂,这表明经长期选育浆蜂上颚腺的磷酸化过程总体上较意蜂更为复杂。主成分分析(principal component analysis,PCA)通过降维的方式将高维数据投影到低维向量空间中,从而既保留数据的主要信息,又使其更易处理[34]。浆蜂和意蜂哺育蜂时期PCA的距离较远,共有蛋白数目较少,进一步说明磷酸化蛋白质组的差异主要体现在哺育蜂时期,因为该时期是蜂王浆分泌的高峰,随着浆蜂蜂王浆产量的提高,其中的10-HDA含量也要相应提高[11]以保证蜂王浆的营养功能,此时浆蜂上颚腺复杂的磷酸化过程可能是与浆蜂提高10-HDA的合成代谢相关。蛋白基序(motif)又称为超二级结构,指相邻的蛋白质二级结构单元相互接近,形成有规律的二级结构聚集体。即两个或两个以上二级结构单元由连接多肽连接起来,组合成有特殊几何排列的局部空间结构。Motif有5种类型,脯氨酸介导的(pro-directed)、酸性的(acidic)、碱性的(basic)、其他(other)和酪氨酸介导的(tyrosine)[35]。蛋白质磷酸化修饰通过激酶实现,激酶识别蛋白质中相对应的motif氨基酸序列,蛋白在蛋白激酶和磷酸酶的作用下发生磷酸化和去磷酸化,激活或关闭某些信号通路[36]。在上颚腺发育的不同时期,浆蜂、意蜂motif数量发生变化;同一时期浆蜂motif总量高于意蜂,表明浆蜂较意蜂具有更复杂的磷酸化机制。浆蜂出房蜂时期特异识别到的Casein kinase 1激酶家族,常作为信号转导途径的调节因子,参与DNA修复和转录[37],在Wnt信号通路中发生磷酸化等[38]。Wnt信号在动物胚胎的早期发育、器官形成过程中作用显著[39],在出房蜂时期支撑上颚腺的基础发育。通路富集的结果表明浆蜂在出房蜂时期的蛋白质表达更为丰富,经过王浆高产的选育,浆蜂从出房蜂开始就与意蜂的上颚腺磷酸化蛋白质谱表现出差异。浆蜂高表达蛋白较多的集中在“Hippo”信号通路。此通路参与调节细胞增殖和凋亡[40]、细胞周期和生长发育等过程[41],在浆蜂出房蜂时期高表达,说明浆蜂从出房蜂开始形态发育较意蜂更为发达,而上颚腺形态发育是其充分发挥功能的基础,这可能与浆蜂哺育蜂时期分泌10-HDA能力增强有关。浆蜂、意蜂哺育蜂时期motif均特异识别到的Pyruvate dehydrogenase kinase(PDK)激酶家族,其底物是丙酮酸脱氢酶,可以催化丙酮酸转化为乙酰辅酶A,从而参与TCA循环氧化供能,PDK进而调节细胞内能量代谢的过程[42]。乙酰辅酶A还是脂肪酸从头合成的起始底物[43]。PDK在哺育蜂时期高表达,表明其可能参与调节哺育蜂时期能量供应和基础物质的代谢。浆蜂哺育蜂较意蜂还特异识别到GSK3, Erk1, Erk2 and CDK5 kinase家族,它们都可调节细胞增殖分化相关过程[44-46],GSK3激酶还可以调节能量代谢相关过程[47]。在浆蜂中特异识别到上述激酶家族,推测其可能支持浆蜂上颚腺的形态发育,从而为其充分发挥分泌功能提供基础。脂肪酸合成的起始物质是乙酰辅酶A,通过合成硬脂酸(18C)[43],进行脱氢、水化、再脱氢、硫解等一些列氧化循环步骤[48],逐步生成低碳数脂肪酸。10-HDA的合成过程中发生脂肪酸的ω-氧化[43],需要细胞色素P450酶的参与[49],生成的脂酰辅酶A通过酰基辅酶A结合蛋白转运进入过氧化物酶体,完成脂肪酸的β-氧化过程[50]。其中水化反应需要烯脂酰辅酶A水合酶的参与[48]。浆蜂哺育蜂时期,乙酰辅酶A合成酶、细胞色素P450、酰基辅酶A结合蛋白、过氧化物酶、烯脂酰-辅酶A水合酶蛋白的高表达,为浆蜂哺育蜂分泌能力提高提供了物质基础,与其大量合成10-HDA的现象相符,其证据是在非流蜜期,浆蜂和意蜂产浆量相差约10倍,但二者蜂王浆的10-HDA含量基本持平[11]。浆蜂、意蜂采集蜂时期motif均特异识别到的14-3-3蛋白家族,是一类保守的调节分子家族,14-3-3蛋白能够结合多种信号蛋白[51],包括Bcl-2相关死亡启动子(BAD)蛋白[52]以及与细胞凋亡相关的信号通路蛋白等。这表明在采集蜂时期,上颚腺腺体功能发生转变,调节相关的信号通路发生变化。其余非特异性识别到的激酶家族,例如GSK3糖原合成酶激酶3家族,也参与细胞增殖、凋亡相关的信号通路[53]和Wnt信号通路等[54]。
4 结论
通过对浆蜂和意蜂出房蜂、哺育蜂、采集蜂3个发育时期上颚腺磷酸化蛋白质组进行研究,发现在出房蜂时期,磷酸化蛋白主要促进腺体的基础发育及形态发育,为哺育蜂时期大量产浆提供基础;浆蜂和意蜂在3个发育时期采用不同的磷酸化蛋白质组来支撑上颚腺功能的发挥;其中哺育蜂时期,两者间图谱差异最大,磷酸化蛋白主要参与调控上颚腺能量代谢和脂肪酸代谢等过程,表明经过蜂王浆高产的选育,浆蜂已具有不同于意蜂的磷酸化蛋白质组,以维持其蜂王浆高产时10-HDA含量的提高,保证蜂王浆的营养功能。(责任编辑 岳梅)
The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}