0 引言
【研究意义】种子的萌发率对于农业生产非常重要,提高种子萌发率,可以降低人力、物力和财力的消耗,提高粮食亩产。作物种子萌发率受种子休眠及萌发机制相互作用的制约,同时也受环境条件的影响。其中,水分是影响种子萌发的主要因素之一,种子萌发并开始生长时,需要吸收大量水分达到吸胀状态以保障发芽。提高干旱胁迫条件下作物的萌发率,有助于维持产量稳定,保障农业的可持续发展。目前,对于调控作物种子萌发率的分子机制研究较少。另外,谷子起源于中国,具有耐旱等突出特性,是中国重要的粮食作物,是单子叶作物抗逆功能研究的重要模式作物[1]。NAC转录因子家族是一个植物特有的转录因子超级家族[2],许多NAC转录因子响应非生物胁迫[3]。因此,研究谷子种子萌发期抗逆相关基因的调控途径,对提高作物的抗逆性具有重要意义。【前人研究进展】在种子休眠过程中起关键作用的内源信号分子是脱落酸(abscisic acid,ABA)和赤霉素(gibberellin acid,GA)[4],其中ABA促进种子脱水耐性和种子休眠的获得,抑制种胚萌发;而GA解除种子休眠,促进萌发,种子休眠或萌发取决于这两种激素的平衡调控。ABA是调控植物干旱胁迫响应的主要植物激素,当植物感受到土壤水分亏缺后,引起ABA合成大量增加[5],从而减弱和植物生长有关的新陈代谢活动,同时抑制种子萌发[6]。NAC转录因子家族的成员在N端具有一个高度保守的DNA结合区(NAC结构域),大约包含150个氨基酸残基,NAC结构域含有A、B、C、D、E 5个亚结构域,C端区域高度变异[7]。NAC转录因子家族可以特异地结合CGT(G/A)和CACG顺式作用元件,调节下游目标基因的表达[8]。NAC转录因子和多种生物过程相关,包括细胞分裂、顶端分生组织发育、次生壁形成、侧根形成、叶片衰老和响应多种生物胁迫和非生物胁迫[2,9]。NAC转录因子调控植物抗逆反应的研究较多[10]。NAC家族的一些成员在模式植物如拟南芥,作物如大豆、水稻、小麦和玉米中被发现,在干旱胁迫信号途径中发挥重要作用。过表达拟南芥NAC基因ANAC019、ANAC055和ANAC072提高了转基因植物的抗旱性[11]。NAC转录因子参与水稻非生物胁迫的响应,比如过表达OsNAC9、OsNAC5或者OsNAP提高了转基因水稻的抗寒性和耐盐性[12]。在一些转基因水稻中,ABA在NAC转录因子调节非生物胁迫的信号通路中发挥重要作用[12]。水稻中有许多ABA信号途径依赖型的NAC转录因子,例如OsNAC5、OsNAC6和OsNAC10通过调节特异的目标基因提高了转基因植株的抗旱性[2,9]。【本研究切入点】目前,关于ABA相关的抗逆调控信号途径研究较多,但是,关于干旱条件下调节作物种子萌发过程的机制研究较少。谷子具有抗旱、耐瘠薄的特点,但对谷子抗逆的分子机制仍然了解较少[13-14]。中国农业科学院作物科学研究所马有志研究员课题组通过对谷子干旱、低氮、低钾转录组测序,分析发现许多谷子抗逆基因响应谷子胁迫处理,其中SiNAC18响应干旱胁迫较强[15]。【拟解决的关键问题】从谷子转录组测序结果中挑选了干旱显著表达上调的NAC类转录因子基因SiNAC18(Si003116,XM_004967871),通过在拟南芥中过表达SiNAC18并分析其生物学功能,同时对其参与的信号途径进行分析,为揭示谷子干旱条件下影响种子萌发的调控途径创造条件。1 材料与方法
试验于2016年在中国农业科学院作物科学研究所完成。1.1 试验材料
所用谷子品种为龙谷25,由中国农业科学院作物科学研究所刁现民研究员课题组提供。拟南芥野生型种子(Columbia-0)、植物表达载体pBI121和绿色荧光蛋白标记载体16318hGFP由中国农业科学院作物科学研究所马有志研究员课题组保存。高保真DNA聚合酶PrimeSTAR HS DNA Polymerase、In-Fusion HD Cloning System试剂盒购于宝生物工程有限公司(TaKaRa)。利福平、氨苄青霉素、卡那霉素和完全培养基混合物(MS粉)等购自北京拜尔迪生物技术有限公司。琼脂糖凝胶回收试剂盒、植物总RNA提取试剂盒、FastQuant RT Super Mix反转录试剂盒和Real Master Mix(SYBR Green)试剂盒均购自天根生化科技(北京)有限公司。试验所用引物合成及测序工作由北京奥科鼎盛生物科技有限公司完成。1.2 谷子SiNAC18的生物信息学分析
在NCBI数据库(https://www.ncbi.nlm.nih.gov/ gene/)中用谷子SiNAC18的蛋白序列进行BLASTP比对,得到与其同源性较高的其他物种的基因序列后用ClustalX 2.0软件进行多序列比对;根据拟南芥NAC家族分组标准搜索与谷子和水稻同源性最高的基因,用MEGA 5.05软件以相邻法(Bootstrap设为1 000)构建系统进化树。1.3 SiNAC18的表达模式分析
取长势一致的谷子幼苗分别进行ABA(100 μmol·L-1)、NaCl(100 mmol·L-1)、干旱(6% PEG-6000)等处理。取未处理(0 h)和处理1、6、12、24和48 h的整个植株以及未处理的材料,用液氮速冻后放置-80℃冰箱保存。用植物总RNA提取试剂盒提取谷子的RNA,用FastQuant RT Super Mix反转录试剂盒合成cDNA。使用RealMaster Mix(SYBR Green)试剂盒和实时荧光定量PCR仪(ABI7500)进行real-time PCR反应。以SiActin(Si001873m.g)为内参基因(表1)。采用三步法反应程序,95℃ 15 min;95℃ 5 s,60℃ 20 s,72℃ 30 s后收集荧光信号,40个循环。采用2-ΔΔCt法根据各样品在特定荧光阈值下的Ct值计算基因在不同样品中的相对表达量。Table 1
表1
表1SiNAC18相关引物
Table 1SiNAC18 related primers
| 引物 Primer | 引物序列 Primer sequence (5′-3′) | 用途 Function | 退火温度 Tm (℃) |
|---|---|---|---|
| N18 | F1:ATGCAGCTGGATGACTGGG | 基因克隆 Gene cloning | 53 |
| R1:TCATATTCTGTCCGGCAGAT | |||
| N18 | F2:ATCTCTAGAGGATCCATGCAGCTGGATGAC | 构建GFP载体 Vector construction of GFP | 65 |
| R2:GCTCACCATGGATCCGCATATTCTGTCCGG | |||
| N18 | F3:GACTCTAGAGGATCCATGCAGCTGGATGAC | 构建pBI121载体 Vector construction of pBI121 | 62 |
| R3:ACCACCCGGGGATCCTCATATTCTGTCCGG | |||
| N18 | F4:TCGCACAAGAGGCGATTCAT | 实时定量 PCR Real-time PCR | 55 |
| R4:GGCTCAAACAGGGAGAAGGT | |||
| AtPIP2A | F:CGATGGCTACAGCACAGGGAC | 实时定量PCR Real-time PCR | 60 |
| R:TACCATGAACACGGCAAATCC | |||
| AtP5CR | F:CGCGGTGGGGAAGATATTGA | 实时定量 PCR Real-time PCR | 60 |
| R:GCTAACCATCGTTGCAGCTC | |||
| AtP5CS2 | F:TACAGTTCCAAGGCCTGCAC | 实时定量 PCR Real-time PCR | 55 |
| R:CAGCACCAAGTCCGAACCTA | |||
| AtPRODH | F:GATCTGCTGCGGTGGGAATA | 实时定量 PCR Real-time PCR | 55 |
| R:TGGAGGATTGTGTCTTCCGC | |||
| AtPRX34 | F:CCAAACTCTTCGTGGACTATGC | 实时定量 PCR Real-time PCR | 55 |
| R:AACTCTTGGTCGCTCTGGAT | |||
| AtRD29A | F:ACGGGATTTGACGGAGAACC | 实时定量 PCR Real-time PCR | 58 |
| R:CCGTCTTTGGGTCTCTTCCC | |||
| SiActin | F:GGCAAACAGGGAGAAGATGA | 谷子内参基因 Internal control gene in Setaria italica | 60 |
| R:GAGGTTGTCGGTAAGGTCACG | |||
| AtActin | F:TGTTCCCATCAGAACCGTGA | 拟南芥内参基因 Internal control gene in Arabidopsis thaliana | 60 |
| R:CACCTGTCTTTGGGTCAACAA |
新窗口打开
1.4 SiNAC18蛋白亚细胞定位
扩增SiNAC18的编码序列,构建p16318hGFP- SiNAC18融合表达载体,以谷子叶片为材料参考YOO等[16]的方法进行原生质体的制备和转化,室温避光培养16 h以上,使用激光共聚焦显微镜(Zeiss LSM700)观察SiNAC18蛋白亚细胞定位情况。1.5 SiNAC18转基因植株的获得
通过In-Fusion® HD Cloning Kit克隆试剂盒(Clontech)将扩增得到的SiNAC18 cDNA序列参考CLOUGH等[17]的方法,插入到带有CaMV 35S启动子的pBI121表达载体上,采用农杆菌转化法转入GV3101农杆菌感受态,转化拟南芥。在含卡那霉素(50 mg·L-1)的MS 培养基上对收获的拟南芥种子进行筛选,经筛选和扩繁后,选择纯合的T3代种子用于表型鉴定试验。1.6 SiNAC18转基因拟南芥种子萌发期抗逆性鉴定
将拟南芥野生型(WT)及SiNAC18转基因株系OE1和OE2的种子首先用70%的酒精处理2 min,无菌水漂洗3次,每次2 min,然后用1%的次氯酸钠溶液处理12 min,期间颠倒离心管数次,无菌水漂洗3次后,将种子点播于MS培养基、PEG-6000浓度为10%和15%的MS培养基,ABA浓度为2和5 μmol·L-1 MS培养基上,低温春化3 d后,将培养皿置于温度22℃,相对湿度65%,16 h光照/8 h黑暗的条件下进行培养。每隔12 h统计萌发率。1.7 胁迫相关基因在SiNAC18转基因植株中的表达分析
用植物总RNA提取试剂盒提取SiNAC18转基因拟南芥的RNA,用FastQuant RT Super Mix反转录试剂盒合成cDNA。根据相关文献报道,选择胁迫相关基因AtPIP2A、AtP5CR、AtPRODH、AtPRX34和AtRD29A,设计引物,以AtActin(AT3G15260m.g)为内参基因(表1),使用RealMaster Mix(SYBR Green)试剂盒和实时荧光定量PCR仪(ABI7500)进行RT-PCR反应。采用三步法反应程序,95℃ 15 min;95℃ 5 s,60℃ 20 s,72℃ 30 s后收集荧光信号,40个循环。采用2-ΔΔCt法根据各样品在特定荧光阈值下的Ct值计算基因在不同样品中的相对表达量。2 结果
2.1 SiNAC18生物信息学分析
谷子基因SiNAC18全长1 074 bp,编码由357个氨基酸组成的亲水性蛋白,分子量约为38.8 kD。NAC家族蛋白具有称为NAC结构域(矮牵牛NAM结构域和拟南芥ATAF1、ATAF2和CUC2结构域)[18]的共有序列,其位于N-末端区域并被分为5个亚结构域(A—E)[19-20]。在Phytozome中寻找和SiNAC18同源性最高的其他物种包括水稻、拟南芥、大豆和玉米中NAC类转录因子,用ClustalX 2.0对SiNAC18及其同源序列进行比对分析,发现这些蛋白N端都具有A、B、C、D和E这5个保守结构域,蛋白C端具有高度多态性(图1),证明SiNAC18的N端序列与其结合下游基因启动子元件相关。根据OOKA等[21]的报道,NAC转录因子家族可分为Ⅰ和Ⅱ两个大组,其中Ⅰ组又分为14个亚组,Ⅱ组分为4个亚组。根据拟南芥和水稻中NAC转录因子家族的分类结果,选择每个亚组中同源性最高的蛋白,构建物种间系统进化树,结果显示,SiNAC18属于第Ⅰ组的NAP亚组,与拟南芥基因AtNAC29同源性最高。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1SiNAC18与其他物种同源NAC类蛋白的氨基酸序列比对
黑色下划线标记为NAC转录因子N端保守域A、B、C、D和E(A:LPPGFRFPTDEELYL;B:ADYKPWLP;C:EWYFFSPRDRKYPNGRPN RGSGYWKATGTDKI;D:GKKALVFYGPKGKTWIMHEYRL;E:LDWVLCRIYKK)。黑色阴影表示保守区,*标注保守氨基酸
-->Fig. 1Multiple alignment of amino acid sequence of SiNAC18 and homologous NAC like proteins from other plant species
The NAC conserved sequence A, B, C, D, E is underlined. (A: LPPGFRFPTDEELYL; B: ADYKPWLP; C: EWYFFSPRDRKYPNGRPNRG SGYWKATGTDKI; D: GKKALVFYGPKGKTWIMHEYRL; E: LDWVLCRIYKK). Black shading indicates conserved domain, and * labelled for conserved amino acid
-->
2.2 SiNAC18亚细胞定位
将p16318hGFP-SiNAC18融合表达载体和GFP空载体质粒转化到谷子原生质体中,在激光共聚焦显微镜下观察SiNAC18-GFP融合蛋白的亚细胞定位情况。结果显示,GFP空对照蛋白在细胞核、细胞质和细胞膜中均有表达(图2-A),而SiNAC18-GFP融合蛋白只在细胞核中显示绿色荧光,表明SiNAC18 蛋白定位在胞核中(图2-B)。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2SiNAC18蛋白亚细胞定位
A:对照GFP;B:融合蛋白SiNAC18-GFP
-->Fig. 2Subcellular localization of SiNAC18
A: Control GFP; B: Fusion protein SiNAC18-GFP
-->
2.3 SiNAC18表达模式分析
用ABA(100 μmol·L-1)、NaCl(100 mmol·L-1)、H2O2(100 mmol·L-1)、干旱(6% PEG-6000)等不同胁迫条件处理谷子幼苗,液体MS培养基处理作为对照,通过real-time PCR检测谷子幼苗中SiNAC18的表达情况(图3),结果显示,SiNAC18在ABA处理条件下表达量明显提高,在处理后24 h表达量达到处理前的9倍;SiNAC18在NaCl处理条件下表达也上调,在处理后24 h表达量达到处理前的5倍;在过氧化氢(H2O2)处理下,处理48 h后表达量是处理前的9倍;在PEG-6000处理情况下,SiNAC18表达量上升,在24 h时表达量最高,达到处理前的22倍。结果表明,SiNAC18在谷子受ABA、高盐、过氧化氢、干旱等非生物胁迫处理下诱导表达。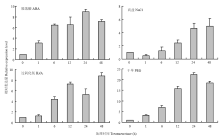 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3不同处理条件下SiNAC18表达谱
-->Fig. 3Expression profile of SiNAC18 under various stress treatment conditions
-->
2.4 转SiNAC18拟南芥种子萌发期抗旱性
将SiNAC18转基因拟南芥种子在ABA、NaCl、H2O2和PEG胁迫处理下进行抗性分析,结果显示,在ABA和PEG胁迫处理下,SiNAC18转基因拟南芥与野生型种子的萌发率存在明显差异;在高盐和H2O2胁迫下,转基因植株与野生型植株相比差异不显著。在MS培养基上,野生型拟南芥WT和转SiNAC18拟南芥种子的萌发速率基本一致(图4-A、4-D);但是在PEG-6000浓度为10%的MS培养基上,转基因株系OE1和OE2种子在36 h时萌发率为60%,是WT(20%)的3倍;到第3天时,转基因株系OE1和OE2(85%)的萌发率比WT(75%)高10%(图4-B、4-E);在PEG-6000浓度为15%的MS培养基上,转基因株系OE1和OE2的萌发率仍然显著高于WT(图4-C、4-F),结果表明,转SiNAC18基因拟南芥在种子萌发期相对于WT有较强的抗旱性,SiNAC18正向调控植物在干旱条件下的种子萌发过程。另一方面,ABA处理结果显示在MS培养基上,WT种子和转基因SiNAC18株系OE1和OE2的种子萌发率基本一致(图5-A、5-D);在ABA浓度为2 μmol·L-1的MS培养基上,种子萌发被抑制,在6 d后WT(100%)种子萌发率显著高于转基因株系OE1和OE2(60%)(图5-B、5-E);在ABA浓度为5 μmol·L-1的MS培养基上,在5 d后转基因株系(15%)的种子萌发率比WT(40%)更低(图5-C、5-F),结果表明转SiNAC18基因拟南芥相对于WT增强了对ABA的敏感性,SiNAC18正向调控植物ABA信号途径。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4SiNAC18转基因拟南芥种子在不同浓度PEG-6000处理下的萌发率
A、D:种子在MS对照条件下的萌发率;B、E:种子在MS+10%PEG处理条件下的萌发率;C、F:种子在MS+15%PEG处理条件下的萌发率
-->Fig. 4Seed germination rates of SiNAC18 transgenic lines under different concentrations of PEG-6000 treatment conditions
A, D: Seed germination rates under condition of MS; B, E: Seed germination rates under condition with MS+10%PEG; C, F: Seed germination rates under condition with MS+15%PEG
-->
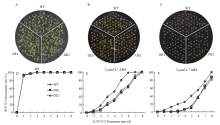 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5SiNAC18转基因拟南芥种子在不同浓度ABA处理下的萌发率
A、D:种子在MS对照条件下的萌发率;B、E:种子在MS+2 μmol·L-1 ABA处理条件下的萌发率;C、F:种子在MS+5 μmol·L-1 ABA处理条件下的萌发率
-->Fig. 5Seed germination rates of SiNAC18 transgenic lines under different concentrations of ABA treatment conditions
A, D: Seed germination rates under condition of MS; B, E: Seed germination rates under condition with MS+2 μmol·L-1 ABA; C, F: Seed germination rates under condition with MS+5 μmol·L-1 ABA
-->
2.5 ABA、胁迫响应相关基因在转SiNAC18植株中的表达
为研究SiNAC18在种子萌发期提高转基因株系抗旱能力的机制,选取了部分ABA及胁迫响应相关基因,检测这些基因在转基因株系和野生型WT中的表达差异(图6)。结果显示,ABA信号途径相关基因AtRD29A在转基因株系中的表达量比WT要高,表明SiNAC18通过激活ABA相关信号通路基因影响种子萌发期的干旱胁迫抗性;在转基因株系中,脯氨酸合成相关基因AtP5CR和AtPRODH的表达量明显高于在WT中的表达量,表明转SiNAC18拟南芥通过激活AtP5CR和AtPRODH的表达来合成脯氨酸抵抗干旱胁迫;过氧化物酶基因AtPOD在转基因株系中的表达量也高于WT中的表达量,证明氧化胁迫相关基因也受SiNAC18调控;水通道蛋白基因AtPIP2A在转基因株系中的表达量比WT要高,表明SiNAC18通过调控水分运输抵抗种子萌发期的干旱胁迫;脯氨酸合成相关基因AtP5CS2在转基因植株和野生型中的表达量没有规律变化。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图6在PEG处理条件下WT和SiNAC18转基因植株中下游基因表达分析
-->Fig. 6Expression analyses of downstream genes in WT and SiNAC18 transgenic plants under PEG stress conditions
-->
3 讨论
干旱是限制作物产量的一个主要的非生物胁迫因素,抗旱相关转录因子能够参与下游基因的表达调控和信号传导,激活许多下游抗逆功能基因表达,从而综合提高植物的抗逆性。迄今发现的与抗旱相关的转录因子主要有DREB、MYC/MYB、bZIP、WRKY和NAC类等基因家族。NAC转录因子家族是一个植物特有的转录因子超级家族,调节下游一系列基因的表达来参与植物各种生理生化活动,在调节植物的生长,响应逆境胁迫中发挥重要作用[2,9]。本研究从谷子中克隆了一个NAC类转录因子基因SiNAC18,并对其表达模式进行分析,结果表明,SiNAC18在谷子中受ABA、高盐、过氧化氢、干旱等胁迫处理的诱导表达(图3)。大量研究表明过表达抗旱相关转录因子可以提高植物生长期的抗旱性,如转SNAC1水稻在营养生长期表现出明显的耐旱和抗盐性,同时转SNAC1水稻对脱落酸(ABA)更敏感,气孔关闭更快使得水分丧失减少,而光合速率并没有显著差异[22]。目前,关于植物抗旱性的研究大多集中在营养生长及生殖生长时期,对种子萌发期抗旱性的分子机制研究较少。ABA是种子萌发与休眠调控过程中起关键作用的内源信号分子,在种子吸水阶段ABA的含量是种子能否完成萌发的决定因素[23],拟南芥中CYP707A2蛋白的合成可以促进ABA降解,促进种子萌发[24]。也有研究证明,在ABA存在的条件下,拟南芥干旱诱导蛋白AtDi19-3的突变体与野生型相比种子萌发和子叶绿化速率更高,Atdi19-3突变体显示更大的干旱耐受性,表明AtDi19-3可以以ABA依赖性方式参与植物对干旱胁迫的反应[25]。本研究中,根据表达模式分析的结果,对SiNAC18转基因拟南芥进行抗性分析,结果显示,在ABA和PEG胁迫处理下,SiNAC18转基因拟南芥的萌发率与野生型拟南芥相比存在差异,在NaCl和H2O2胁迫下,转基因植株无明显表型。在干旱处理条件下,SiNAC18转基因株系比野生型WT的萌发率高(图4),表明转SiNAC18拟南芥在种子萌发期相对于WT有较强的抗旱性。干旱条件下提高作物的萌发率,可以保证作物的出苗率,从而维持产量的稳定。另一方面,ABA处理结果显示,WT种子萌发率高于转基因株系OE1和OE2(图5),表明转SiNAC18拟南芥相对于WT增强了对ABA的敏感性,SiNAC18正向调控植物ABA信号途径,SiNAC18通过调控植物ABA信号途径影响植物在种子萌发期的抗旱性,进一步分析SiNAC18调控的下游网络对于了解植物在萌发期的抗旱机制具有重要意义。
本研究中的表达模式分析结果显示SiNAC18受高盐、干旱等胁迫的诱导表达,然而转SiNAC18拟南芥只对干旱胁迫表现耐性,而对高盐及H2O2胁迫没有表现耐性。由干旱和高盐等造成的水分胁迫可以诱导许多基因的表达[26-27],然而不是所有基因都能对胁迫抗性产生影响,有些基因只是植物遭受胁迫时所作出的受害反应。因此,即使这些基因受胁迫诱导表达它们对植物的抗旱反应也不能产生影响。另外,盐胁迫主要表现为离子毒害和渗透胁迫[28]两个方面,干旱胁迫主要表现为渗透胁迫[29]。因此,高盐及干旱胁迫在作用机制方面有所不同,转SiNAC18拟南芥只表现抗旱性没有表现耐盐性说明SiNAC18在植物抗旱及耐盐响应中发挥不同的作用,或者在耐盐响应SiNAC18需要通过其他基因共同发挥作用,因此,转SiNAC18拟南芥对干旱及高盐胁迫抗性的存在差异。
当植物受到高盐或干旱等非生物胁迫时,会激活ABA依赖途径和非依赖途径两种信号途径来抵御不良环境的影响[18]。有研究报道拟南芥G蛋白β亚基突变体agb1-2相对于野生型拟南芥可以增强种子萌发期对ABA的敏感性,同时提高植物耐旱性,进一步研究证明AGB1通过下调AtMPK6、AtVIP1和AtMYB44级联参与调控拟南芥中的ABA信号传导途径和耐旱性[30]。为了研究SiNAC18在种子萌发期提高转基因株系抗旱能力的机制,选取了响应ABA及胁迫的相关基因,检测各个基因在转基因株系和野生型WT中的表达情况(图6)。结果显示,ABA信号途径相关基因AtRD29A、脯氨酸合成相关基因AtP5CR和AtPRODH、过氧化物酶基因AtPRX34在转基因株系中的表达量明显高于在WT中的表达量。研究发现ABA可以促进AtP5CR的表达[31],AtRD29A和AtP5CR的启动子区存在ABRE结合元件。同时,过氧化物酶基因AtPRX34通过H2O2和生长素之间的相互作用调节植物的生长与发育[32]。以上结果表明SiNAC18可以通过调控下游基因包括ABA相关信号通路基因、氧化胁迫相关基因的表达提高干旱条件下作物的萌发率。
本研究初步证明谷子SiNAC18在干旱胁迫响应过程中发挥重要作用,可能通过正向调控植物ABA信号途径、氧化胁迫调控等途径影响植物种子萌发期的抗旱性,从而保障种子的正常生长,有助于维持作物在干旱条件下产量的稳定。
4 结论
从谷子中克隆得到了一个NAC类转录因子基因SiNAC18。该基因受多种胁迫处理如干旱和ABA等的诱导表达,其蛋白定位于细胞核。在拟南芥中过表达该基因提高了种子萌发期对干旱胁迫的抗性及对ABA的敏感性。响应ABA及胁迫的相关基因在转SiNAC18基因株系中的表达量高于在WT中的表达量,表明SiNAC18可能通过ABA信号途径、氧化胁迫调控等途径来提高转基因拟南芥在干旱条件下的种子萌发率。The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}