0 引言
【研究意义】大麦属于“三高”药食同源作物[1],其籽粒及其食品因富含蛋白质、总黄酮、γ-氨基丁酸(gamma-aminobutyric acid,GABA)及β-葡聚糖等营养功能成分,被美国农业部列为保健食品[2],对预防和治疗慢性疾病有特殊功效和巨大药用价值[3]。由于蛋白质、总黄酮和GABA含量遗传易受环境影响,传统育种方法培育其高含量大麦品种非常困难[4]。因此,对大麦籽粒蛋白质、总黄酮和GABA含量的QTL进行定位,对分子标记辅助选育符合大麦保健食品加工的功能大麦品种具有重大意义。【前人研究进展】大麦是遗传学研究的重要模式植物,绘制分子遗传图谱为QTL定位奠定了基础。已知大麦第一张连锁图谱是利用RFLP标记针对第6染色体绘制的[5]。随着分子标记的不断开发和深入研究,WENZL等[6-7]、KARAKOUSIS等[8]、READ等[9]、ROSTOKS等[10]、HEARNDEN等[11]、MARCEL等[12]利用多种分子标记绘制了多张大麦的遗传连锁图谱。由于构建的材料、群体、分子标记及种植环境等因素不同,图谱密度不能满足分子标记辅助育种的需求,仍需进行完善。已知加杨[13]、大豆[14]、苦荞[15]等作物的蛋白质和功能成分含量之间有一定相关性,这些成分的相关性揭示了分子标记间的关联,但对大麦籽粒蛋白质和功能成分含量之间的相关性研究未见报道。关于大麦蛋白质含量QTL定位研究已有一些报道。MATHER等[16]在二棱大麦构建的DH群体发现在第1、4、5和7染色体上存在4个蛋白质QTL;LI等[17]在大麦第2和7染色体上发现2个蛋白质QTL;NETSVETAEV[18]在大麦第4和7染色体上分别发现2个控制清蛋白的QTL;大麦7条染色体上49个DarT标记与4种蛋白质组分含量关联,且绝大多数蛋白质组分与总蛋白质含量为相同基因控制[19];因受小麦6B染色体Gpc-B1(与蛋白质含量有关)被图位克隆的影响,位于大麦6H染色体上hvm74标记附近的同源基因与提高蛋白质含量有关[20]。大麦总黄酮和GABA是复杂的次生代谢产物,主要受遗传控制,环境效应也具有一定的调控作用[21-23]。关于总黄酮含量的遗传研究,杨晓梦等[24]对大麦籽粒总黄酮含量进行了遗传分析,发现其遗传符合E-1模型,主基因和多基因效应为86.20%,剩下13.8%由环境决定;刘仙俊[25]和郭蕾蕾[4]在大麦第2、3和6染色体上发现控制总黄酮含量的微效QTL;JENDE[26]克隆了18个影响大麦黄酮合成的基因,其中,结构基因5个,转录调控基因1个。关于GABA含量的遗传研究,已知大麦籽粒GABA含量遗传符合E-0模型,主基因效率较大(56.49%)[24]。在大麦第1、3和4染色体上发现影响GABA含量的QTL[4,27]。【本研究切入点】目前,国内外对大麦籽粒蛋白质和功能成分含量之间的定位研究较少,对两者之间的相关性研究也未见报道。【拟解决的关键问题】本研究采用蛋白质、总黄酮和GABA含量差异较大的紫光芒裸二棱与Schooner构建的包含193个植株的重组自交系(RIL)群体绘制遗传连锁图谱,对籽粒蛋白质、总黄酮和GABA含量进行QTL定位分析,挖掘控制这些成分的基因位点,在基因水平上揭示其相关性,为进一步的精细定位和克隆奠定基础,为分子标记辅助选育高蛋白高功能成分含量大麦新品种提供理论参考。1 材料与方法
1.1 试验材料
供试材料为母本紫光芒裸二棱(P1,云南高蛋白地方青稞品种)、父本Schooner(P2,澳大利亚引进的优质啤酒大麦品种)及其构建的包含193个植株的RIL群体。1.2 田间种植
2007年,以紫光芒裸二棱为母本,Schooner为父本杂交获得F2代种子,2008—2011年加代繁殖至F7代RIL群体。2012年和2013年10月将两亲本和RIL群体193个单株种植于玉溪市研和镇(海拔1 638 m),随机区组排列条播,行长2 m,每行150粒种子,行距0.3 m,3次重复。该试验地前茬是水稻,粘壤土且肥力中上等,常规栽培管理。成熟期收获种子,自然风干后,各株系取10 g种子用打粉机打成粉末状低温贮存,用于蛋白质和功能成分含量的测定分析。1.3 蛋白质、总黄酮和GABA含量的测定
采用《食品中蛋白质的测定(GB5009.5-2010)》标准的凯氏定氮法,由农业部农产品产量监督检验测试中心(昆明)对籽粒蛋白质进行含量的测定。在云南省农业生物技术重点实验室参照赵春艳等[28]的硝酸铝络合比色法对籽粒总黄酮含量进行测定;参照赵大伟等[29]的比色法测定GABA含量,3次重复,取平均值。
1.4 SSR检测
所用的373对SSR引物基本覆盖大麦基因组,引物序列来自网站(www.gramene.org),由北京鼎国昌盛生物技术有限责任公司合成。取少量大麦苗期新鲜叶片,按照CTAB法进行DNA提取。
PCR反应体系为10 μL,包括上下游引物各0.5 μL、DNA 2 μL、PCR Buffer 1.5 μL、dNTPs 0.4 μL、ddH2O 4.9 μL和Taq polymerase 0.2 μL。PCR扩增程序为94℃ 5 min;94℃ 40 s,55℃ 40 s,72℃ 1 min,35个循环;72℃ 5 min,4℃保存。
PCR扩增产物用垂直板聚丙烯凝胶电泳分离,银染显色检测。
1.5 带型记录
按照QTL IciMapping软件的记录方法,带型和母本(紫光芒裸二棱)一致记为“0”,和父本(Schooner)一致记为“2”,杂合带记为“1”,缺失记为“-1”。1.6 QTL定位分析
采用QTL IciMappingV3.3构建大麦RIL群体遗传图谱,并用MapDrawV2.1绘制,借助完备区间作图法(ICIM)进行QTL分析,以LOD≥2.5作为QTL判断阈值,分析表型性状与标记带型之间的连锁关系,确定目标性状QTL位点和估算遗传效应[30]。QTL命名遵循MCCOUCH等[31]方法。2 结果
2.1 SSR标记分布情况
利用373对SSR引物对亲本紫光芒裸二棱和Schooner进行多态性筛选,筛选出136对多态性较好的引物(图1)。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1SSR引物在群体中的扩增情况
-->Fig. 1Segregation of SSR maker in the mapping population
-->
2.2 遗传图谱
筛选出的多态性标记有135对,分布在大麦7条染色体上,包括7个连锁群,除了Bmac0144未连接到连锁群上。该遗传图谱总长为2 224.29 cM,两标记间的平均间距为16.48 cM。在该遗传图谱中,多数标记位于的连锁群与前人发表的连锁染色体基本一致[32]。2.3 大麦籽粒数量性状定位
2.3.1 蛋白质、总黄酮和GABA含量表型分析 使用SPSS17.0软件对2年试验数据进行统计分析(表1)。亲本间及其RIL群体之间在蛋白质、总黄酮和GABA含量均存在极显著差异(P<0.01),母本紫光芒裸二棱在蛋白质、总黄酮和GABA含量上是高值亲本,父本Schooner是低值亲本。RIL群体间蛋白质、总黄酮和GABA含量具有较大的变异幅度。从性状分布的偏度和峰度值来看,其绝对值基本<1,从频率分布趋势(图2)可看出各性状表现为连续变异,具有一定超亲分离现象,表现出典型的数量性状特征,表明大麦营养功能成分遗传是受多基因影响,可进行QTL定位分析。Table1
表1
表1亲本及其RIL群体营养功能成分表型值(2012年和2013年)
Table1Phenotypic values of parents and its RIL population for functional components contents (2012 and 2013)
| 年份 Year | 性状 Traits | 亲本Parents | 重组自交系群体 RILs population | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 紫光芒裸二棱 Ziguangmang | Schooner | 最大值 Max | 最小值 Min | 均值±标准误 Mean±SE | 变异系数 CV(%) | 偏度 Skewness | 峰度 Kurtosis | ||
| 2012 | 蛋白质Protein (%) | 19.3 | 12.7 | 19.4 | 12.4 | 15.52±0.09 | 8.18 | 0.13 | 0.02 |
| 总黄酮 Total flavonoids (mg·100g-1) | 338.08 | 284.05 | 429.39 | 201.71 | 299.53±3.15 | 14.65 | 0.18 | -0.37 | |
| γ-氨基丁酸 GABA (mg·100g-1) | 15.84 | 12.88 | 21.02 | 9.11 | 13.76±0.15 | 15.19 | 0.69 | 0.76 | |
| 2013 | 蛋白质Protein (%) | 22.85 | 10.78 | 20.46 | 9.51 | 14.32±0.17 | 16.01 | 0.06 | -0.54 |
| 总黄酮 Total flavonoids (mg·100g-1) | 260.54 | 210.38 | 401.25 | 157.35 | 273.86±3.68 | 18.65 | 0.32 | -0.59 | |
| γ-氨基丁酸 GABA (mg·100g-1) | 19.58 | 10.98 | 31.35 | 4.49 | 11.67±0.36 | 42.76 | 1.33 | 1.78 | |
新窗口打开
2.3.2 蛋白质、总黄酮和GABA含量之间的相关性分析 通过进行相关性分析(表2),发现2年试验的相关分析结果一致。蛋白质、总黄酮和GABA含量之间呈极显著正相关,且正相关性较大,即籽粒蛋白质含量越高,其总黄酮和GABA含量就越高。说明控制这些成分的基因可能存在紧密连锁,因此,应注意这些成分的关联,为功能大麦品质育种提供一些科学依据。
2.3.3 蛋白质、总黄酮和GABA含量的QTL定位分析
采用ICIM法,取LOD≥2.5为QTL存在的判断值,对2012年和2013年大麦籽粒蛋白质、总黄酮和GABA含量进行QTL定位和遗传效应分析。2012年,试验共检测到8个QTL(表3,图3),其中,3个蛋白质含量QTL、2个总黄酮含量QTL、3个GABA含量QTL。2013年试验共检测到12个QTL(表3,图3),其中,6个蛋白质含量QTL、5个总黄酮含量QTL和1个GABA含量QTL。
2012年,定位于4H、6H和7H 3条染色体上的籽粒蛋白质、总黄酮和GABA含量共8个QTL,其中,4H染色体检测到2个QTL位点,6H和7H各检测到3个QTL位点。3个蛋白质QTL分别为qPC-4H、qPC-6H和qPC-7H,位于4H、6H和7H,LOD值范围为2.89—4.46,表型变异率范围为6.45%—15.45%,最大变异率为15.45%的主效QTL位于7H,表明蛋白质含量是由少数主效及微效基因共同影响的数量性状。4H染色体上的加性效应是正值(0.379),表明提高蛋白质含量的等位基因源于高值母本紫光芒裸二棱,而6H和7H上为负值(-0.362和-0.506),表明增效基因源于父本Schooner;2个总黄酮QTL分别为qTFC-6H和qTFC-7H,分布在6H和7H上,其LOD值为3.14、3.26,表型贡献率为10.38%和15.27%,可视为主效QTL,说明总黄酮含量主要由少数主效基因影响。7H染色体上的加性效应为正值(17.124),表示提供总黄酮含量的增效基因来自高值母本紫光芒裸二棱,而6H染色体上的为负值(-14.107),表明增效基因来自父本Schooner;3个GABA的QTL分别为qGABA-4H、qGABA-6H和qGABA-7H,位于4H、6H和7H,LOD值范围为2.85—4.96,表型变异率范围为5.44%—14.87%,最大变异率为14.87%的主效QTL位于7H,说明GABA含量由少数主效及微效基因共同影响。7H染色体上的加性效应为正值(0.803),表明增加GABA含量的等位基因源于高值母本紫光芒裸二棱,4H、6H上的加性效应为负值(-0.532和-0.548),表明父本Schooner提供增效基因。此外,位于6H Ebmac0806—GBM1270区间的2个QTL同时影响蛋白质和GABA含量,且加性作用方向一致,显示该区域是QTL效应的集中区域。
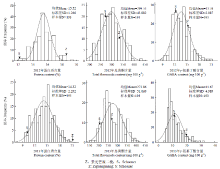 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2大麦籽粒蛋白质、总黄酮和GABA含量在RIL群体中的频率分布(2012年和2013年)
-->Fig. 2Frequency distribution of protein, total flavonoids and GABA content in RIL (2012 and 2013)
-->
Table 2
表2
表2大麦籽粒蛋白质、总黄酮和GABA含量之间的相关性
Table 2The correlation among protein, total flavonoids and GABA in barley grains
| 年份 Year | 性状 Trait | 蛋白质 Protein | 总黄酮 Total flavonoids | γ-氨基丁酸 GABA |
|---|---|---|---|---|
| 2012 | 蛋白质 Protein | 1.000 | ||
| 总黄酮 Total flavonoids | 0.993** | 1.000 | ||
| γ-氨基丁酸 GABA | 0.985** | 0.986** | 1.000 | |
| 2013 | 蛋白质 Protein | 1.000 | ||
| 总黄酮 Total flavonoids | 0.570** | 1.000 | ||
| γ-氨基丁酸 GABA | 0.352** | 0.413** | 1.000 |
新窗口打开
2013年,定位于1H、2H、4H、5H、6H和7H等6条染色体上的籽粒蛋白质、总黄酮和GABA含量共12个QTL,其中,1H和4H染色体各检测到1个QTL位点,2H、5H和6H各检测到2个QTL位点,7H染色体检测到4个QTL位点。6个蛋白质QTL分别为qPC-1H、qPC-2H、qPC-4H、qPC-6H、qPC-7Ha和qPC-7Hb,位于1H、2H、4H、6H和7H,LOD值范围为2.76—9.69,表型变异率范围为4.11%—18.86%,变异率为13.30%和18.86%的2个主效QTL分别位于6H和7H,表明蛋白质含量受少数主效及微效基因共同影响。1H、2H、6H和7H染色体上的加性效应均为负值(-0.466、-0.647、-0.946、-0.509和-0.996),表明增效基因源于父本Schooner,而4H上为正值(0.700),表明提高蛋白质含量的等位基因源于高值母本紫光芒裸二棱。4H和6H染色体检测到的2个QTL位点(qPC-4H和qPC-6H)与2012年定位的一致,7H染色体的1个QTL位点(qPC-7Ha)与2012年的相邻,且加性作用方向一致,说明这三个蛋白质QTL位点比较可靠;5个总黄酮QTL分别为qTFC-2H、qTFC-5H、qTFC-6H、qTFC-7Ha和qTFC-7Hb,分布在2H、5H、6H和7H上,其LOD值范围为3.35—9.89,表型贡献率范围为6.06%—29.01%,变异率为 17.55%、24.17%和29.01%的3个主效QTL分别位于2H和7H,说明总黄酮含量也受到少数微效基因影响。7H染色体上的加性效应均为正值(25.313和27.640),表示提供总黄酮含量的增效基因来自高值母本紫光芒裸二棱,而2H、5H和6H染色体上均为负值(-21.395、-16.154和-14.347),表明增效基因来自父本Schooner。7H染色体检测到的1个QTL位点(qTFC-7Hb)与2012年定位的一致,且加性作用方向一致,说明该总黄酮QTL位点可靠性较大;1个GABA QTL为qGABA-5H,位于5H染色体,LOD值为3.76,表型变异率为8.78%,是微效QTL。加性效应为-1.479,表明由父本Schooner提供增效基因。GABA QTL仅在单一年份被检测到,说明GABA含量相关QTL受环境影响较大。此外,位于2HBmag0125—GBM1309和6H Ebmac0806—GBM1270区间的4个QTL同时影响蛋白质和总黄酮含量,且加性作用方向一致,显示这两个区域是QTL效应的集中区域。
Table 3
表3
表3大麦籽粒蛋白质、总黄酮和GABA含量的QTL分析(2012年和2013年)
Table 3QTL analysis for protein, total flavonoids and GABA content of barley grains (2012 and 2013)
| 年份 Year | 性状 Trait | QTL | 连锁群 Chr.location | 标记区间 Marker interval | 区间距离 Interval distance (cM) | LOD值 LOD value | 贡献率 Contribution (%) | 加性效应 Additive effects |
|---|---|---|---|---|---|---|---|---|
| 2012 | 蛋白质 Protein | qPC-4H | 4H | BMAG0740—BMAG0808 | 16.19 | 4.46 | 9.01 | 0.379 |
| qPC-6H | 6H | Ebmac0806—GBM1270 | 6.81 | 2.95 | 6.45 | -0.362 | ||
| qPC-7H | 7H | EBmac755—Bmag0507 | 49.17 | 2.89 | 15.45 | -0.506 | ||
| 总黄酮 Total Flavonoids | qTFC-6H | 6H | Bmac0316—BMAG0807 | 41.3 | 3.26 | 10.38 | -14.107 | |
| qTFC-7H | 7H | EBmatc0016—Bmag0206 | 42.68 | 3.14 | 15.27 | 17.124 | ||
| γ-氨基丁酸 GABA | qGABA-4H | 4H | GBM1422—GBM1236 | 1.89 | 2.85 | 5.44 | -0.532 | |
| qGABA-6H | 6H | Ebmac0806—GBM1270 | 6.81 | 2.90 | 5.57 | -0.548 | ||
| qGABA-7H | 7H | EBmac0764—Bmac0273 | 26.45 | 4.96 | 14.87 | 0.803 | ||
| 2013 | 蛋白质 Protein | qPC-1H | 1H | scssr08238—scssr02748 | 24.31 | 2.76 | 4.11 | -0.466 |
| qPC-2H | 2H | Bmag0125—GBM1309 | 36.4 | 2.80 | 7.95 | -0.647 | ||
| qPC-4H | 4H | BMAG0740—BMAG0808 | 16.19 | 7.02 | 9.33 | 0.700 | ||
| qPC-6H | 6H | Ebmac0806—GBM1270 | 6.81 | 8.59 | 13.30 | -0.946 | ||
| qPC-7Ha | 7H | Bmag0507—Bmag0321 | 3.32 | 3.34 | 4.57 | -0.509 | ||
| qPC-7Hb | 7H | Scssr07970—EBmac603 | 22.97 | 9.69 | 18.86 | -0.996 | ||
| 总黄酮 Total Flavonoids | qTFC-2H | 2H | Bmag0125—GBM1309 | 36.40 | 6.28 | 17.55 | -21.395 | |
| qTFC-5H | 5H | Bmag0222—GBM1176 | 47.39 | 3.35 | 9.37 | -16.154 | ||
| qTFC-6H | 6H | Ebmac0806—GBM1270 | 6.81 | 4.00 | 6.06 | -14.347 | ||
| qTFC-7Ha | 7H | GBM1516—EBmatc0016 | 24.30 | 9.89 | 24.17 | 25.313 | ||
| qTFC-7Hb | 7H | EBmatc0016—Bmag0206 | 42.68 | 9.86 | 29.01 | 27.640 | ||
| γ-氨基丁酸 GABA | qGABA-5H | 5H | GMS2—Scssr10148 | 37.54 | 3.76 | 8.78 | -1.479 |
新窗口打开
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3大麦籽粒蛋白质、总黄酮和γ-氨基丁酸含量QTL在染色体上的位置(2012年和2013年)
-->Fig. 3Positions of QTLs for protein, total flavonoids and GABA content of barley (2012 and 2013)
-->
3 讨论
3.1 大麦遗传图谱中标记距离的差异分析
在本文构建的遗传图谱中,大多标记所在的连锁染色体与前人结果较一致[32],但在染色体上的顺序和个别标记所属染色体有所变化,这可能是构建的群体类型及数量不同、标记出现偏分离和多等位位点造成;另外部分标记之间的遗传距离也有差异,这可能是因为遗传距离是根据标记间的重组率估计的一个相对值,标记在图谱中的位点也是相对的,根据遗传标记和群体大小的不同产生一定差异[33]。3.2 大麦籽粒蛋白质、总黄酮和GABA含量QTL一致性分析
目前,研究已发现大麦染色体上影响蛋白质、总黄酮和GABA含量的QTL,且结果不尽一致。本研究通过2年数据研究,共检测到9个大麦籽粒蛋白质含量的QTL,其中在4H和6H连锁群定位一致,在7H连锁群定位相邻。HAYES等[34]在六棱大麦构建的DH群体发现了分布在第2、3、4、6和7染色体上5个影响蛋白质含量的QTL位点;任喜峰[35]在2H、5H、7H上检测到3个蛋白质QTL。李解等[36]在2H和6H上发现2个影响蛋白质含量的单一QTL位点,和本研究共同的是2H、4H、6H和7H连锁群,且本研究在2H上发现的QTL位点与任喜峰所报道的位点较相近,说明2H、4H、6H和7H连锁群定位的位点可靠性较大。本研究2年共检测到7个有关总黄酮含量的QTL,其中,7H连锁群上有一个QTL位点定位一致,另一位点相邻。刘仙俊[25]在大麦2H、3H和6H检测到3个总黄酮微效QTL;郭蕾蕾[4]在2H、3H、5H和6H发现6个总黄酮微效QTL,和本研究共同的是2H、5H和6H连锁群,且本研究在6H上发现的QTL位点与郭蕾蕾所报道的位点相邻,说明2H、5H和6H连锁群定位的位点可能是准确的。
ZENG等[27]在大麦3H和4H染色体上发现4个影响GABA含量的QTL。郭蕾蕾[4]在大麦1H染色体检测到1个影响GABA含量的微效QTL。罗小娇等[37]在大麦3HL染色体上定位到1个微效QTL。本研究2年共检测到4个有关GABA含量的QTL,和ZENG报道的有一个共同的4H连锁染色体,且位点较相近,说明在4H连锁群定位的位点可靠性较大。
本研究对3种营养功能成分定位的结果与别人的研究结果不太一致,这可能是因为蛋白质,总黄酮和GABA含量属于复杂的数量性状,易受环境的影响。QTL对不同环境下产生的基因表达水平不同,造成同一性状在不同环境下QTL的差异,形成定位结果不一致。此外还可能跟构建的材料背景,群体的类型和大小不同有很大关系。所以应该严格控制环境因素的影响,进行多年多点或同一地点,不同年份(不同时期)的成分含量测定和重复QTL检测;扩大定位群体和缩小标记区间,以发现较稳定表达的QTL位点,进一步探索大麦籽粒营养功能成分积累的分子机制。
3.3 大麦籽粒蛋白质、总黄酮和GABA含量及其QTL的相关性分析
蛋白质、总黄酮和GABA是重要的代谢产物,植物体内的初级代谢产物苯丙氨酸是合成蛋白质和黄酮类化合物骨架的前体物质[13],而GABA是由谷氨酸转化得到,三者代谢途径之间可能存在一定的内在关联。已报道一些作物的蛋白质和功能成分含量间具有一定相关性,王彩霞[38]发现有色稻米的功能营养成分和蛋白质呈显著或极显著正相关;梁慧珍等[14]报道了大豆的蛋白质和异黄酮含量的负相关性达极显著,并在G染色体上找到了影响异黄酮和蛋白质的部分QTL基因位点;刘三才等[15]发现荞麦蛋白质与总黄酮含量的正相关性极显著(0.588)。但对大麦籽粒蛋白质和功能成分含量之间的相关分析未见报道。本研究发现大麦籽粒蛋白质、总黄酮和GABA含量三者间呈极显著正相关,说明控制这些成分的基因可能存在多效性或紧密连锁。2012年试验检测到控制蛋白质、总黄酮和GABA含量3种成分的QTL位点主要分布于6H和7H染色体,其中,表型变异率大于10%的部分主效QTL分布于7H;6H Ebmac0806- GBM1270区间的QTL位点同时影响蛋白质和GABA含量。2013年试验检测到控制蛋白质和总黄酮含量QTL位点主要分布于6H和7H染色体,总黄酮和GABA含量QTL位点主要分布于5H染色体。贡献率大于10%的部分主效QTL也集中分布于7H、2H Bmag0125—GBM1309和6H Ebmac0806—GBM1270区间同时影响蛋白质和总黄酮含量。这些区域是QTL效应的集中区域,特别是两年都检测到的6H Ebmac0806—GBM1270区间,都有影响蛋白质、总黄酮和GABA含量。导致这种现象可能是因为这些区域QTL存在“一因多效”或基因紧密连锁,是成分关联的遗传基础。其中,蛋白质与GABA、蛋白质与总黄酮含量的QTL位点关联性最紧密,总黄酮与GABA含量之间的位点关联性次之。
综上所述,成分的相关性能反映出分子标记的关联,3种重要营养功能成分的部分定位结果与其相关分析是一致的,说明蛋白质和功能成分之间的遗传关系较紧密,这将有助于营养功能成分的聚合,筛选和培育出高蛋白高功能成分含量的大麦新品种。而这几种成分是由同一基因还是多个紧密连锁的基因控制及具体的遗传关联机制仍将有待于利用更高密度的连锁图谱和进一步精细定位深入探究。
4 结论
大麦籽粒营养功能成分之间的相关性分析表明,蛋白质、总黄酮与GABA含量之间呈极显著正相关。其QTL定位和遗传效应分析表明,控制蛋白质含量的9个QTL分别定位于1H、2H、4H、6H和7H这5条染色体上;控制总黄酮含量的7个QTL分别定位于2H、5H、6H和7H这4条染色体上;控制GABA含量的4个QTL分别定位于4H、5H、6H和7H这4条染色体上。其中控制蛋白质含量的2个QTL、总黄酮含量的1个QTL至少在两年试验均可检测到,GABA含量QTL仅在单一年份被检测到,说明GABA含量相关QTL受环境影响较大。控制蛋白质含量基因与总黄酮含量基因同位于2H、6H和7H染色体,控制蛋白质含量基因与GABA含量基因重合在4H、6H和7H染色体,控制总黄酮含量基因与GABA含量基因同位于5H、6H和7H染色体,尤其是6H Ebmac0806—GBM1270区间都影响蛋白质、总黄酮和GABA含量,且加性作用方向一致,有极显著相关性。亲本紫光芒裸二棱和Schooner均有提供增效的等位基因。因此,大麦籽粒蛋白质、总黄酮和GABA含量的相关性分析与其部分QTL定位结果是一致的,揭示了蛋白质和功能成分含量之间紧密的遗传关系。The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}