0 引言
【研究意义】前期研究中,LI等[1]提出了以六倍体AnAnCnCnCoCo为桥梁,与白菜型油菜杂交,创建新型甘蓝型油菜,拓宽甘蓝型油菜遗传背景的策略。本研究将调查以甘蓝型油菜与菌核病抗性甘蓝杂交合成的六倍体为桥梁,与白菜型油菜杂交的可交配性,考察杂种的苗期表型和结实率,并以菌核病抗性为目标性状,评价该策略的应用潜力。这对利用甘蓝和白菜型油菜遗传资源,拓宽和改良甘蓝型油菜遗传背景具有重要意义。【前人研究进展】甘蓝型油菜(Brassica napus,AACC,2n=38)是世界上重要的油料作物之一,是由甘蓝和白菜杂交,染色体自然加倍而成[2-4]。与亲本种相比,甘蓝型油菜的遗传背景相对狭窄[5],利用亲本种遗传资源是拓宽现有甘蓝型油菜遗传背景的重要途径[6-10]。亲本种的利用途径主要有:(1)利用甘蓝与白菜型油菜杂交,人工合成甘蓝型油菜[11-12],该方法需结合胚挽救和染色体加倍等组织培养技术;(2)利用甘蓝型油菜与亲本种杂交,自交或回交,选育新型甘蓝型油菜[4,13-15],该方法从后代中筛选新型甘蓝型油菜所需的周期长,效率低。除此以外,钱伟等[16]发明了一种以甘蓝型油菜与甘蓝杂交创建的六倍体AnAnCnCnCoCo为桥梁,大量快速利用白菜型油菜遗传资源,创建新型甘蓝型油菜,拓宽甘蓝型油菜遗传背景的新策略。在此基础上,MEI等[17]利用六倍体作为桥梁,与甘蓝型油菜不断回交,获得了转移甘蓝菌核病抗性的甘蓝型油菜。【本研究切入点】本研究将采用改良菌核病抗性的六倍体为桥梁,与大量的白菜型油菜杂交,试图导入甘蓝型油菜亲本种优良遗传性状,并利用亲本种广泛的遗传资源,改良现有甘蓝型油菜遗传背景。【拟解决的关键问题】利用菌核病抗性甘蓝与甘蓝型油菜杂交获得的六倍体为桥梁,与白菜型油菜杂交,通过考察可交配性、杂种育性和菌核病抗性来分析该策略对拓宽和改良甘蓝型油菜背景的应用潜力。1 材料与方法
试验于2013—2015年在西南大学进行。1.1 试验材料
前期,利用甘蓝型油菜(中双9号)与菌核病抗性甘蓝(C01)杂交,合成了六倍体(AnAnCnCnCoCo)[17]。2013年将74份半冬性、20份春性和16份冬性白菜型油菜(电子附表1)与六倍体杂交,创建新型甘蓝型油菜。成熟期,统计各杂交组合的角果数和种子数,检测可交配性。2014和2015年,杂种F1种植于西南大学油菜工程技术研究中心试验地,用于杂种育性检测和菌核病抗性鉴定。菌核病抗性鉴定采用耐病品种中双9号作为对照。Table S1
附表1
附表1110份白菜型油菜类型
Table S1110 types of B. rapa
| 编号 Code | 类型 Type | 来源 Origin | 生态型 Ecotype |
|---|---|---|---|
| 1 | PI223501 | 阿富汗 Afghanistan | 半冬性 Semi-winter |
| 2 | PI432372 | 孟加拉 Bangladesh | 半冬性 Semi-winter |
| 3 | PI392024 | 加拿大 Canada | 半冬性 Semi-winter |
| 4 | PI226505 | 伊朗 Iran | 半冬性 Semi-winter |
| 5 | PI227296 | 伊朗 Iran | 半冬性 Semi-winter |
| 6 | PI138894 | 伊朗 Iran | 半冬性 Semi-winter |
| 7 | PI458618 | 新西兰 New Zealand | 半冬性 Semi-winter |
| 8 | PI443024 | 挪威 Norway | 半冬性 Semi-winter |
| 9 | PI269431 | 巴基斯坦 Pakistan | 半冬性 Semi-winter |
| 10 | PI209020 | 波多黎各 Puerto Rico | 半冬性 Semi-winter |
| 11 | PI263055 | 俄罗斯 Russian Federation | 半冬性 Semi-winter |
| 12 | PI360878 | 瑞典 Sweden | 半冬性 Semi-winter |
| 13 | 1075 | 中国西南大学 Southwest University, China | 半冬性 Semi-winter |
| 14 | 0002-1 | 中国西南大学 Southwest University, China | 半冬性 Semi-winter |
| 15 | 0005-1 | 中国西南大学 Southwest University, China | 半冬性 Semi-winter |
| 16 | 0007-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 17 | 0010-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 18 | 0011-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 19 | 0014-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 20 | 0015-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 21 | 0016-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 22 | 0020-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 23 | 0021-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 24 | 0023-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 25 | 0027-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 26 | 0028-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 27 | 0237-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 28 | 0243-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 29 | 0245-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 30 | 0249-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 31 | 0260-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 32 | 0267-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 33 | 0281-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 34 | 0340-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 35 | 0426-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 36 | 0427-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 37 | 0434-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 38 | 0465-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 39 | 0475-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 40 | 0477-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 编号 Code | 类型 Type | 来源 Origin | 生态型 Ecotype |
| 41 | 0489-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 42 | 0595-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 43 | 0768-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 44 | 0770-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 45 | 1773-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 46 | 毕节油菜 Bijieyoucai | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 47 | 5K1002 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 48 | 黄油1号 Huangyou 1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 49 | 6R095 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 50 | 6Y733 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 51 | 6Y812 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 52 | 7B12-1 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 53 | 7C-126 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 54 | 9B05-3 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 55 | 高棋种 Gaoqizhong | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 56 | 黄岩宁波种 Huangyanningbozhong | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 57 | R8021 | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 58 | 大冶矮脚白油菜 Daye aijiaobaiyoucai | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 59 | 云梦半直立油菜 Yunmeng banzhiliyoucai | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 60 | 沅江桂林子 Yuanjiang guilinzi | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 61 | 治城小花油菜 Zhicheng xiaohuayoucai | 中国 西南大学 Southwest University, China | 半冬性 Semi-winter |
| 62 | Ames 30081 | 美国 United States | 半冬性 Semi-winter |
| 63 | 巢湖油菜 Caohu youcai | 中国安徽 Anhui, China | 半冬性 Semi-winter |
| 64 | 苏州青 Suzhouqing | 中国江苏 Jiangsu, China | 半冬性 Semi-winter |
| 65 | 丰城孙渡油菜 Fengcheng sunduyoucai | 中国江西 Jiangxi, China | 半冬性 Semi-winter |
| 66 | 大悟洋白菜 Dawu yangbaicai | 中国湖北 Hubei, China | 半冬性 Semi-winter |
| 67 | 宜昌三区油菜 Yichang sanquyoucai | 中国湖北 Hubei, China | 半冬性 Semi-winter |
| 68 | 湘乡油菜 Xiangxiang youcai | 中国湖南 Hunan, China | 半冬性 Semi-winter |
| 69 | 武冈油菜白 Wugang youcaibai | 中国湖南 Hunan, China | 半冬性 Semi-winter |
| 70 | 长沙姑香 Changsha guxiang | 中国湖南 Hunan, China | 半冬性 Semi-winter |
| 71 | 永结油菜 Yongjie youcai | 中国甘肃 Gansu, China | 半冬性 Semi-winter |
| 72 | 秦油4号 Qinyou 4 | 中国陕西 Shanxi, China | 半冬性 Semi-winter |
| 73 | 宁强仓社矮油菜 Ningqiang cangsheaiyoucai | 中国陕西 Shanxi, China | 半冬性 Semi-winter |
| 74 | 青油13号 Qingyou 13 | 中国青海Qinghai, China | 半冬性 Semi-winter |
| 75 | PI254543 | 阿富汗Afghanistan | 春性 Spring |
| 76 | PI254542 | 阿富汗Afghanistan | 春性 Spring |
| 77 | PI268371 | 阿富汗 Afghanistan | 春性 Spring |
| 78 | PI162778 | 阿根廷 Argentina | 春性 Spring |
| 79 | PI365643 | 加拿大 Canada | 春性 Spring |
| 80 | CGN06842 | 德国科隆 Cologne, Germany | 春性 Spring |
| 81 | CGN06831 | 德国科隆 Cologne, Germany | 春性 Spring |
| 编号 Code | 类型 Type | 来源 Origin | 生态型 Ecotype |
| 82 | CGN06832 | 德国科隆 Cologne, Germany | 春性 Spring |
| 83 | CGN06838 | 德国科隆 Cologne, Germany | 春性 Spring |
| 84 | CGN06839 | 德国科隆 Cologne, Germany | 春性 Spring |
| 85 | CGN06840 | 德国科隆 Cologne, Germany | 春性 Spring |
| 86 | CGN06841 | 德国科隆 Cologne, Germany | 春性 Spring |
| 87 | PI347605 | 印度 India | 春性 Spring |
| 88 | PI370740 | 印度 India | 春性 Spring |
| 89 | PI267726 | 日本 Japan | 春性 Spring |
| 90 | PI343183 | 荷兰 Netherlands | 春性 Spring |
| 91 | PI458977 | 美国 United States | 春性 Spring |
| 92 | 藏油10号 Zangyou 10 | 中国西藏 Xizang, China | 春性 Spring |
| 93 | 隆孜县当日镇俗坡村小油菜 Supocun xiaoyoucai, Dangri, Longzi | 中国西藏 Xizang, China | 春性 Spring |
| 94 | 隆孜县日当镇日当村小油菜 Ridangcun xiaoyoucai, Ridang, Longzi | 中国西藏Xizang, China | 春性 Spring |
| 95 | PI254541 | 阿富汗 Afghanistan | 冬性 Winter |
| 96 | PI268369 | 阿富汗 Afghanistan | 冬性 Winter |
| 97 | PI432376 | 孟加拉 Bangladesh | 冬性 Winter |
| 98 | CGN06819 | 德国科隆 Cologne, Germany | 冬性 Winter |
| 99 | CGN15216 | 德国科隆 Cologne, Germany | 冬性 Winter |
| 100 | ECD04 | 德国 Germany | 冬性 Winter |
| 101 | Neib03-2 | 德国 Germany | 冬性 Winter |
| 102 | PI458616 | 新西兰 New Zealand | 冬性 Winter |
| 103 | PI458931 | 瑞典 Sweden | 冬性 Winter |
| 104 | PI458932 | 瑞典 Sweden | 冬性 Winter |
| 105 | PI177286 | 土耳其 Turkey | 冬性 Winter |
| 106 | PI205283 | 土耳其 Turkey | 冬性 Winter |
| 107 | PI176885 | 土耳其 Turkey | 冬性 Winter |
| 108 | Ames 30084 | 美国 United States | 冬性 Winter |
| 109 | PI633153 | 法国 France | 冬性 Winter |
| 110 | PI603024 | 巴基斯坦 Pakistan | 冬性 Winter |
新窗口打开
1.2 试验方法
1.2.1 染色体行为分析 现蕾期,取幼嫩花蕾,于卡诺固定液(酒精:醋酸=3:1)固定24 h以上,取幼嫩子房用于体细胞染色体数目的观察;取幼嫩花药用于减数分裂时期染色体行为观察。具体方法参照LI等[18]。1.2.2 菌核病抗性鉴定 本研究采用的菌核病接种和抗性鉴定方法,参照MEI等[19]的离体茎秆接种方法。在90 mm的无菌PDA培养基中,接种菌核病菌丝,于22℃,湿度85%的条件下培养菌丝2 d,待菌丝布满整个培养基时,用直径6 mm的打孔器沿培养皿边缘一圈打孔,获得的菌丝块用于茎秆接种。青荚期,截取植株的茎秆,每个株系取3—5个单株,每个单株取30 cm长的茎秆,两端包裹保鲜膜,每个茎秆接种2个菌丝块于22℃、湿度85%的密闭环境中培养。接种3 d后,统计菌斑长度,并以中双9号作为对照,计算材料的相对感病度S=V/V对照,其中,V为待测材料的菌斑长度,V对照为‘中双9号’的病斑长度。S值越小,表示抗性越强;反之,抗性越弱。
1.3 数据分析
可交配性和育性均采用每角果粒数来衡量,六倍体与白菜型油菜杂交的可交配性=饱满种子数/角果数;新型甘蓝型油菜的育性为自交结实率,结实率=饱满种子总数/取样角果数。数据采用SAS version 8.0(SAS Institute)软件进行方差分析(ANOVA)和相关性分析。2 结果
2.1 六倍体的特点
本研究采用‘中双9号’与菌核病抗性甘蓝‘C01’(B. incana)杂交,合成了六倍体AnAnCnCnCoCo(图1-A)。该六倍体的花粉育性是90.6%—92.7%,自交结实率为3—7粒/角果;菌核病抗性高于甘蓝型油菜‘中双9号’(六倍体:接种3 d后的病斑长度为4.6 cm,‘中双9号’接种3 d后的病斑长度是9.6 cm)[17]。六倍体的体细胞染色体数目为56条(图1-B),68.80%(86/125)处于减数分裂后期I的花粉母细胞是28:28分离(图1-C)。2.2 六倍体与白菜型油菜的可交配性
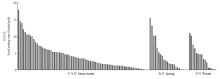
利用六倍体为桥梁,与110份来自3种不同生态型的白菜型油菜杂交,合成杂种。六倍体与白菜型油菜授粉后15 d的胚珠发育正常(图1-D),平均可交配性为(4.25±3.91)粒/角果(最高结实率:17.83粒/角果;最低结实率:0.06粒/角果)。尽管不同基因型的白菜型油菜与六倍体杂交,可交配性存在明显差异,但是来自半冬性、春性和冬性3种生态型的白菜型油菜与六倍体杂交,可交配性没有显著性差异(半冬性:(4.35±3.77)粒/角果;春性:(4.34±4.51)粒/角果;冬性:(4.01±3.43)粒/角果,P=0.44,图2)。六倍体与白菜型油菜杂交正、反交之间的可交配性没有显著性差异(六倍体为母本:平均结实率为4.27粒/角果;六倍体为父本:平均结实率为3.95粒/角果,P=0.69)。本研究说明了六倍体与白菜型油菜正反交都能结籽。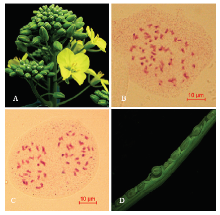 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1六倍体表型、细胞学及与白菜型油菜杂交胚珠发育A:六倍体花期形态;B:六倍体体细胞染色体数目(2n=56条);C:六倍体减数分裂后期I染色体分离比(28:28);D:六倍体与白菜型油菜授粉15 d后的胚珠发育
-->Fig. 1Morphology, cytogenetics of hexaploid and hybrid embryo growth between hexaploid and B. rapaA: Morphology of hexaploid at flowering stage; B: Chromosome number of hexaploid in ovary cell (2n=56); C: Chromosome segregation of hexaploid at anaphase I in PMC (28:28); D: Hybrid embryo growth between hexaploid and B. rapa at 15 days after pollination
-->
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2六倍体与110份白菜型油菜杂交可交配性
-->Fig. 2Crossability between hexaploid and 110 B. rapa
-->
2.3 六倍体与白菜型油菜杂种的形态学和育性
选取种子量多的67份(来自46份半冬性、12份春性和9份冬性白菜型油菜)杂种种植于田间,观察杂种的苗期表型和育性。杂种的苗期形态不同于白菜型油菜,与现有的甘蓝型油菜类似,合成的杂种形态学表型变异丰富(图3)。显示原图|下载原图ZIP|生成PPT图3六倍体与白菜型油菜杂种的苗期表型A:甘蓝型油菜(中双9号),B:六倍体,C—F:六倍体与白菜型油菜杂种,G—I:分别为杂种D—F对应的白菜型油菜亲本
-->Fig. 3Morphology of hybrids between hexaploid and B. rapa at seedling stageA: B. napus (Zhongshuang 9); B: Hexaploid; C-F: Hybrid between hexaploid and B. rapa; G-I: B. rapa parents of D-F, respectively
-->
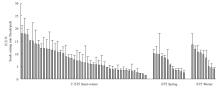
收获期,统计杂种的自交结实率,结果发现所有杂种都能自交结籽,平均结实率为(7.72±4.45)粒/角果。尽管不同杂种之间育性存在显著性差异,但是根据白菜型油菜的生态型,不同生态型之间没有显著性差异(P=0.95,冬性白菜型油菜合成的杂种平均自交结实率:(8.07±3.43)粒/角果;半冬性:(7.88±4.64)粒/角果;春性:(6.41±3.00)粒/角果)(图4)。这说明了不同生态型的白菜型油菜与该六倍体杂交,合成的杂种都能结籽,并且白菜型油菜亲本生态型对合成的杂种结实率没有显著性影响。六倍体与白菜型油菜杂种丰富的表型变异和育性说明了杂种具有拓宽甘蓝型油菜遗传背景的潜力。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图467份白菜型油菜与六倍体杂种的结实率
-->Fig. 4Seed setting rate of hybrids between hexaploid and 67 B. rapa
-->
2.4 六倍体与白菜型油菜杂种的菌核病抗性鉴定
随机选取23份杂种于青荚期截取茎秆进行菌核病抗性鉴定,连续重复两年。结果发现,2014年,这些杂种的平均菌斑长度为(5.77±2.39)cm,相对感病度为0.95±0.42,不同基因型之间存在显著差异(P<0.0001)。7个杂种的菌核病抗性显著高于‘中双9号’(平均菌斑长度为:(6.01±1.41)cm),5份杂种的病斑显著大于‘中双9号’。2015年,该23份杂种的平均菌斑长度为:(4.33±1.69)cm,相对感病度为0.73±0.29,不同基因型之间存在显著性差异(P<0.0012),11份杂种的病斑显著小于‘中双9号’。其中,6份杂种连续两年的菌核病相对感病度显著低于‘中双9号’(图5,表1)。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5六倍体与白菜型油菜茎秆菌核病抗性鉴定
-->Fig. 5Stem responses to S. sclerotiorum in hybrids between hexaploid and B. rapa
-->
Table 1
表1
表123份六倍体与白菜型油菜杂种的茎秆菌核病抗性鉴定
Table 1Stem responses to S. sclerotiorum in 23 hybrids between hexaploid and B. rapa
| 编号 Code | 白菜亲本 B. rapa parents | 病斑长度 Lesion length (cm) | 相对感病度 Relative susceptibility | |||
|---|---|---|---|---|---|---|
| 2014 | 2015 | 2014 | 2015 | |||
| 1 | 0770-1 | 6.30±1.27 | 3.10±0.96* | 1.05±0.21 | 0.52±0.16* | |
| 2 | PI176885 | 4.07±0.15* | 3.30±0.57* | 0.68±0.03* | 0.56±0.10* | |
| 3 | PI458616 | 3.40±0.73* | 2.00±1.13* | 0.57±0.12* | 0.34±0.2* | |
| 4 | CGN06831 | 5.35±0.35 | 3.45±1.06* | 0.89±0.06 | 0.58±0.18* | |
| 5 | CGN06832 | 5.80±0.99 | 4.95±1.06 | 0.97±0.16 | 0.83±0.18 | |
| 6 | PI267726 | 3.20±0.42* | 3.90±0.71 | 0.53±0.07* | 0.66±0.12 | |
| 7 | PI227296 | 5.85±1.03 | 3.50±1.56* | 0.97±0.17 | 0.59±0.26* | |
| 8 | PI432372 | 5.05±1.20 | 4.53±1.53 | 0.84±0.20 | 0.76±0.26 | |
| 9 | CGN15216 | 5.25±1.48 | 3.25±1.34* | 0.87±0.25 | 0.55±0.23* | |
| 10 | 0002-1 | 4.67±1.45 | 4.10±0.99 | 0.78±0.24 | 0.69±0.17 | |
| 11 | 0475-1 | 4.08±0.75* | 3.70±0.41* | 0.68±0.12* | 0.62±0.07* | |
| 12 | 0477-1 | 9.78±1.01* | 6.98±2.34 | 1.63±0.17* | 1.17±0.39 | |
| 13 | 0245-1 | 2.15±0.21* | 3.30±1.13* | 0.36±0.04* | 0.56±0.19* | |
| 14 | 0249-1 | 8.40±0.60* | 4.30±1.56 | 1.40±0.10* | 0.72±0.26 | |
| 15 | 0434-1 | 2.43±0.21* | 3.60±0.57* | 0.40±0.03* | 0.61±0.10* | |
| 16 | PI633153 | 4.67±0.40 | 3.75±0.79* | 0.78±0.07 | 0.63±0.13* | |
| 17 | 0007-1 | 5.93±0.55 | 4.68±1.38 | 0.99±0.09 | 0.79±0.23 | |
| 18 | 0010-1 | 4.07±1.58* | 3.95±0.64* | 0.68±0.26* | 0.66±0.11* | |
| 19 | 0028-1 | 4.53±1.32* | 4.20±1.56 | 0.75±0.22* | 0.71±0.26 | |
| 20 | 宁强仓社矮油菜 Ningqiangcangshe | 4.95±0.35 | 4.45±0.35 | 0.82±0.06 | 0.75±0.06 | |
| 21 | PI370740 | 7.17±0.73* | 6.00±0.57 | 1.19±0.12* | 1.01±0.10 | |
| 22 | PI603024 | 12.23±1.76* | 6.95±3.15 | 2.04±0.29* | 1.17±0.53 | |
| 23 | Ames 30084 | 8.30±0.95* | 4.30±0.85 | 1.38±0.16* | 0.72±0.14 | |
| 中双9号 Zhongshuang 9 | 6.01±1.41 | 5.94±1.06 | 1 | 1 | ||
新窗口打开
将两年的菌核病抗性进行比较,结果发现,尽管不同年度间,杂种的菌核病平均菌斑长度和平均相对感病度存在显著性差异(P<0.0001,P= 0.0004),但是两年的菌核病发病程度极显著正相关(r=0.74,P<0.00001)。说明六倍体与白菜型油菜杂种可转移甘蓝的菌核病抗性,表明该途径具有利用甘蓝亲本的遗传变异改良甘蓝型油菜遗传背景的潜力。
3 讨论
3.1 甘蓝型油菜亲本种的利用途径
甘蓝型油菜是由甘蓝和白菜型油菜杂交,染色体自然加倍而成[2]。甘蓝型油菜起源于7 500年前,但作为栽培种其栽培历史短,经过长期的驯化和进化,甘蓝型油菜的遗传背景比亲本种相对狭窄[3,5,7]。白菜型油菜是一个古老的栽培作物,其农艺特征和DNA结构上具有丰富的遗传变异[20-21]。白菜型油菜与甘蓝型油菜杂交和回交容易,利用该策略创建新型甘蓝型油菜已经在甘蓝型油菜育种中广泛应用[4,6,22]。甘蓝具有丰富的栽培类型和野生资源,以及优良的抗病、抗虫资源[23]。甘蓝型油菜与甘蓝杂交,是利用甘蓝遗传资源的重要手段。将甘蓝型油菜与甘蓝杂交,不断自交,后代也能利用甘蓝遗传资源,合成甘蓝型油菜[14-15]。但是该策略的使用需结合胚挽救获得杂种,杂种F1育性低,需大量的无性系隔离授粉方能获得后代。目前,同时利用白菜型油菜和甘蓝资源的主要途径是人工合成甘蓝型油菜。然而,该途径仍需借助胚挽救等组织培养技术获得杂种,并且后代染色体分离异常,基因组稳定性差[7,24]。钱伟等[16]发明了一种以甘蓝型油菜与甘蓝杂交,染色体加倍获得的六倍体为桥梁,与白菜型油菜杂交,创建新型甘蓝型油菜的方法。该方法不仅利用了甘蓝资源,还导入大量白菜型油菜遗传成分,创建的新型甘蓝型油菜能拓宽现有甘蓝型油菜遗传背景。该策略的提出为本研究的顺利开展提供了思路。本研究提出了以菌核病抗性甘蓝为亲本,与甘蓝型油菜杂交,创建了菌核病抗性改良的六倍体;以该六倍体为桥梁,与大量的白菜型油菜杂交的策略,该策略不仅能利用白菜型油菜丰富资源,还能将甘蓝菌核病抗性导入到油菜中。3.2 菌核病抗性的改良
菌核病是甘蓝型油菜的主要病害之一,严重危害其生产。尽管育种家们已培育出一些相对耐病的油菜品种(‘中油821’和‘中双’系列品种[25];‘先锋45S51’和‘先锋45S52’[26]),然而,甘蓝型油菜中缺乏菌核病抗性资源。笔者在甘蓝资源中发现了一个野生甘蓝(B. incana),对菌核病有突出抗性,其茎秆抗性是甘蓝型油菜耐病品种‘中油821’的20倍[19]。目前,MEI[16]、刘瑶[27]利用该野生甘蓝与甘蓝型油菜杂交,合成了具有菌核病抗性的六倍体,该六倍体与甘蓝型油菜不断回交,选育出的甘蓝型油菜具有改良的菌核病抗性。DING等[28]通过人工合成甘蓝型油菜的途径,将甘蓝亲本的菌核病抗性导入到油菜中。本研究将菌核病抗性甘蓝‘C01’与甘蓝型油菜‘中双9号’杂交,合成的具有菌核病抗性的六倍体为桥梁,与大量白菜型油菜杂交,合成杂种的菌核病抗性存在显著性差异。相比对照品种‘中双9号’,鉴定的23份杂种中,只有6份杂种菌核病抗性高于对照。产生这种现象的主要原因可能是:一方面,白菜型油菜中广泛缺乏菌核病抗性,造成六倍体与白菜型油菜杂种的菌核病抗性降低;另一方面,甘蓝型油菜与甘蓝杂交合成的六倍体(AnAnCnCnCoCo),由于A、C基因组的高度同源性[29-31],六倍体的染色体减数分裂中产生大量的染色体重排,可能导致配子中菌核病抗性位点的丢失。综上所述,本研究实现了一种利用甘蓝型油菜与菌核病抗性甘蓝杂交,创建抗菌核病的六倍体为桥梁,与白菜型油菜杂交,利用亲本种拓宽和改良甘蓝型油菜遗传背景的新途径。
4 结论
本研究将菌核病抗性甘蓝与甘蓝型油菜杂交合成的六倍体,与110份春性、冬性和半冬性的白菜型油菜正反交均能结籽。杂种苗期形态似甘蓝型油菜,但是表型变异丰富;杂种能自交结籽。经过两年的离体茎秆菌核病抗性鉴定,6份杂种的菌核病发病程度两年均显著低于中双9号。本研究提出的利用菌核病抗性甘蓝与甘蓝型油菜杂交合成的六倍体为桥梁,与大量的白菜型油菜杂交合成的杂种策略,具有利用亲本种遗传资源,拓宽和改良甘蓝型油菜遗传背景的潜力。The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}