Effects of Successive Biochar Addition to Soil on Nitrogen Functional Microorganisms and Nitrous Oxide Emission
DONG Cheng, CHEN ZhiYong, XIE YingXin, ZHANG YangYang, GOU PeiXin, YANG JiaHeng, MA DongYun, WANG ChenYang, GUO TianCaiCollege of Agronomy, Henan Agricultural University/National Engineering Research Center for Wheat, Zhengzhou 450002通讯作者:
责任编辑: 李云霞
收稿日期:2019-12-17接受日期:2020-02-16网络出版日期:2020-10-01
| 基金资助: |
Received:2019-12-17Accepted:2020-02-16Online:2020-10-01

摘要
关键词:
Abstract
Keywords:
PDF (1927KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
董成, 陈智勇, 谢迎新, 张阳阳, 缑培欣, 杨家蘅, 马冬云, 王晨阳, 郭天财. 生物炭连续施用对农田土壤氮转化微生物及N2O排放的影响[J]. 中国农业科学, 2020, 53(19): 4024-4034 doi:10.3864/j.issn.0578-1752.2020.19.015
DONG Cheng, CHEN ZhiYong, XIE YingXin, ZHANG YangYang, GOU PeiXin, YANG JiaHeng, MA DongYun, WANG ChenYang, GUO TianCai.
0 引言
【研究意义】生物炭具有含碳量高、比表面积较大、孔隙结构丰富、养分含量高、碳稳定性强等独特的理化性质[1,2],使其近年来在固碳减排[3]、水土保持、减少养分流失[4]、提高作物产量[5]等方面具有良好的应用潜力,但生物炭施用对农田N2O排放和作物产量影响的具体微生物效应机制尚未明确。因此,开展多年连续添加生物炭对农田土壤氮转化微生物功能基因丰度、N2O排放及作物产量影响方面的研究,可为生物炭在农业方面的推广使用提供理论依据。【前人研究进展】施用生物炭对农田N2O排放影响的研究已成为近年来的研究热点,土壤硝化作用和反硝化作用作为农田N2O排放的主要途径[6],但其研究机制存在较大争议,且这种争议主要集中在生物炭制备原材料、生产工艺及土壤类型等方面[7]。VERHOEVEN等[8]通过在14种不同类型土壤中施用9种不同材料得到的生物炭研究认为,生物炭施用可提高土壤pH和减少土壤N2O排放。然而,CLOUGH等[9]研究发现,施用木质生物炭对草地土壤N2O排放并无显著影响。更有研究者发现在燥红壤中施用生物炭后,也可显著促进土壤硝化作用,进而导致土壤N2O在短时间内集中排放[10]。另外,土壤硝化作用过程中氨氧化古菌(ammonia- oxidizing archaea,AOA)和氨氧化细菌(ammonia- oxidizing bacteria,AOB)的氨单加氧酶(amoA)以及反硝化作用过程中对应的亚硝酸盐还原基因nirK、nirS和编码氧化亚氮还原酶的唯一基因nosZ基因[11,12,13]被广泛用作功能标志基因来研究土壤硝化及反硝化过程,成为近些年来的研究热点。王晓辉等[14] 发现,土壤中加入生物炭后,AOA、AOB和nirK基因型反硝化细菌的基因丰度明显增加,但对nirS和nosZ基因丰度没有显著影响。WARNOCK等[15] 研究表明,施用生物炭可减少土壤硝化细菌和反硝化细菌的有效可用氮源,进而起到减少N2O排放以及改变农田氮循环的效果。DUAN等[16]研究发现,新鲜生物炭可提高酸性土壤中nirK、nosZII和碱性土壤中nirK、nosZI,降低碱性土壤AOB基因丰度,从而有效减少N2O排放量,但老化生物炭则会对两种类型土壤硝化和反硝化作用均有显著的促进作用,且显著增加N2O排放。【本研究切入点】当前,施用生物炭对土壤氮转化微生物影响的研究也主要集中在与土壤碳、氮循环相关的几种功能微生物,且其对土壤N2O排放及作物产量影响的报道还存在争议,主要因为以往的研究多为短期的田间试验[17-19] 或室内培养试验[20,21,22,23]。中性或偏碱性的潮土作为中国分布面积最广的农田土壤对全国粮食产量的贡献超过1/5,具有强烈的硝化作用[24]。然而,连续多年在潮土农田施用生物炭后是否改变土壤氮循环相关功能基因丰度及土壤速效氮性质,对土壤硝化作用和反硝化作用又会产生何种影响,且最终是否能对农田生态系统产生增产减排效果,这些问题仍没有明确答案。【拟解决的关键问题】本试验通过设立连续多年生物炭添加农田定位试验,以黄淮典型小麦-玉米轮作潮土农田为研究对象,分析连续添加生物炭处理6年后不同生物炭施用量下夏玉米产量、土壤N2O排放的具体差异,探究连续多年施用生物炭的增产减排潜力及其影响作物产量和N2O排放的微生物学机制,从而揭示长期连续施用生物炭对土壤氮循环微生物、N2O排放及作物生产的具体影响,为生物炭农用和土壤科学管理提供依据。1 材料与方法
1.1 试验地点和供试物料
试验地位于河南省封丘县潘店乡中国科学院封丘农业生态实验站(35°00′N,114°24′E),该地处黄淮海平原中部,毗邻黄河,属暖温带大陆性季风气候,年平均气温14℃,无霜期214 d左右,年平均降水量625 mm,年平均蒸发量1 860 mm。土壤类型为潮土,2011年试验开始时初始土壤基本理化性质为有机质(OM)11.2 g·kg-1、全氮(TN)0.79 g·kg-1、速效磷(P)8.6 mg·kg-1、有效钾(K)92.1 mg·kg-1、pH为8.3(土水比1﹕2.5)。试验用生物炭为水稻秸秆在500℃条件下,以沼气为能源的炭化炉下热解自行制备[25],其基本理化性质为全氮(TN)10.3 g·kg-1、全碳(TC)514.8 g·kg-1、有效磷(P)1.71 g·kg-1、速效钾(K)49.2 mg·kg-1、pH为10.5(炭水比1﹕10)。
1.2 试验设计与土壤样品采集
试验起始于2011年10月,至2018年9月已完成7个小麦/玉米轮作种植周期 (10月至次年6月为小麦季,6—9月为玉米季)。由于当前生物炭在农田的施用量并无统一标准,通常情况下,我国小麦/玉米/水稻秸秆每公顷每季产量约为7.5 t,如将每季生产的秸秆在下季作物开始前全部炭化还田,按照秸秆炭化率30%计算得到的每公顷秸秆生物炭(500℃,4 h)量约为2.25 t [26]。因此,本研究从秸秆炭化还田的良性循环出发,围绕每季秸秆全部炭化后还田的量(2.25 t·hm-2)设置不施秸秆生物炭0(BC0,对照)、2.25 t·hm-2(秸秆全量炭化还田,BCL,低量)、6.75 t·hm-2(3倍秸秆全量炭化还田,BCM,中量)和11.25 t·hm-2(5倍秸秆全量炭化还田,BCH,高量)4个处理,小区面积4 m×4 m,重复3次,随机排列。各小区氮(N)、磷(P)、钾(K)肥施用情况分别为N 200 kg·hm-2 (常规尿素,含量46%)、P2O5 150 kg·hm-2 (重过磷酸钙,44%)和K2O 90 kg·hm-2(硫酸钾,50%)。其中,磷、钾肥作为基肥在播种前一次性施用,氮肥分基肥和拔节肥两次施用,基肥和追肥比例为6﹕4。生物炭与基肥在每次种植作物前均匀撒施于小区内,使其与0—20 cm耕层土壤均匀混合。小麦播种期通常为每年10月10—15日,收获期为每年6月1—5日;玉米播种期通常为每年6月6—10日,收获期为每年9月13—25日。本研究主要为连续生物炭添加6年后的夏玉米季试验结果,夏玉米于2018年6月8日播种,2018年9月13日收获。其他田间栽培管理措施同当地一般农田。土壤样品采用对角线5点取样法分别于2018年6—9月玉米全生育期每小区进行0—20 cm土层采集。同一小区处理新鲜土样均匀混合,过2 mm筛后,一部分于-80℃保存用于微生物DNA提取及测定,另一部分于4℃冰箱中保存用于土壤硝态氮和铵态氮含量的测定。
1.3 测试项目与方法
1.3.1 土壤理化性质测定 新鲜土壤样品采用2 mol·L-1 KCl溶液提取,土、溶液质量体积比为1﹕5,在摇床上振荡1 h,过滤后的提取液采用紫外分光光度计在双波长(220 nm,275 nm)下直接测定硝态氮浓度,采用靛酚蓝比色法在单波长(625 nm)下测定铵态氮浓度。鉴于土壤无机氮含量(土壤Nmin=硝态氮+铵态氮)可以有效表征土壤的供氮能力, 可作为推荐施氮的指标[27],因此,本试验也采用土壤Nmin作为表征多年连续施用生物炭后土壤肥力的一项重要指标。土壤含水量为采用重量烘干法测定,并根据已经测定的土壤容重指标将土壤重量含水量换算为土壤孔隙含水量[28,29](water-filled pore space,WFPS)。
1.3.2 N2O气体收集 N2O气体通过密闭箱法(图1)进行采集,在每季玉米播种前整地时,将直径为30 cm的圆形密闭气体采集箱底座放置于小区固定位置两行玉米之间。采样时间为上午8:00—10:00,在放上密闭箱的第0、15、30和45小时用注射器抽取密闭箱内气体,并利用提前备好的真空瓶自动采集气体后带回实验室进行N2O气体测定。采集气体的同时记录气温、土温及箱内气体温度。N2O气体用气相色谱仪 (Agilent7890A,Agilent Technologies,USA)测定。N2O排放通量计算如下[30]:
F = ρ × H × ΔC/Δt × 273 / (273 + T)
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1田间土壤N2O取样装置
Fig. 1The N2O sampling device in farmland
式中,F为温室气体排放通量(μg·m-2·h-1);ρ为标准状态下N2O气体的密度(1.25 kg·m-3);H为密闭箱高(m);ΔC/Δt为单位时间内气体浓度变化量(μg·h-1);T为密闭箱内平均温度(℃)。N2O季节累积排放量按照排放通量,由时间加权法计算得到。
1.3.3 土壤微生物总DNA提取和荧光定量PCR分析 采用MP Biomedicals公司FastDNA ?SPIN Kit for Soil试剂盒提取土壤总DNA。每个样品称取0.5 g 的土壤到Lysing Matrix E Tube。按照FastDNA ?SPIN Kit for Soil试剂盒方法和步骤进行土壤总DNA提取。用核酸定量仪 (NanoDrop2000)检测DNA的纯度和浓度,用浓度为1% 的琼脂糖凝胶电泳检测提取DNA的完整性。各功能基因的PCR循环条件如表1所示。.
Table 1
表1
表1荧光定量PCR引物及扩增条件
Table 1
| 扩增基因 Target | 引物 Primer | 序列 (5′— 3′) Sequence (5′- 3′) | 荧光定量PCR程序 Real-time PCR profiles | 参考文献 References |
|---|---|---|---|---|
| AOA | Arch-amoA F | STAATGGTCTGGCTTAGACG | 95℃ 5 min;95℃ 30 s,58℃ | [31] |
| Arch-amoA R | GCGGCCATCCATCTGTATGT | 30 s,72℃ 1 min,35 cycles | ||
| AOB | amoA-1F | GGGGTTTCTACTGGTGGT | 95℃ 5 min;95℃ 30 s,58℃ | [32] |
| amoA-2R | CCCCTCKGSAAAGCCTTCTTC | 30 s,72℃ 1 min,35 cycles | ||
| nirK | nirK1F | GGMATGGTKCCSTGGCA | 95℃ 5 min;95℃ 30 s,58℃ | [33] |
| nirK5R | GCCTCGATCAGRTTRTGG | 30 s,72℃ 1 min,35 cycles | ||
| nirS | Cd3aF | GTSAACGTSAAGGARACSGG | 95℃ 5 min;95℃ 30 s,58℃ | [34] |
| R3cdR | GASTTCGGRTGSGTCTTGA | 30 s,72℃ 1 min,35 cycles | ||
| nosZ | nosZ-F | AGAACGACCAGCTGATCGACA | 95℃ 5 min;95℃ 30 s,60℃ | [35] |
| nosZ-R | TCCATGGTGACGCCGTGGTTG | 30 s,72℃ 1 min,35 cycles |
新窗口打开|下载CSV
1.4 数据分析
采用SPSS 23.0进行数据分析,不同处理间作物产量、土壤理化性质以及氨氧化古菌、细菌amoA基因、亚硝酸盐还原菌(nirK、nirS)、氧化亚氮还原菌nosZ基因拷贝数采用 Duncan 新复极差法进行多重比较(P<0.05),并对不同生物炭处理下土壤N2O、NH4+、NO3-、WFPS与微生物基因丰度进行相关性分析(Pearson,Two-tailed)。采用Excel 2010和origin 2017对土壤微生物基因丰度进行作图。2 结果
2.1 施用生物炭对玉米产量的影响
由图2可知,施用生物炭可显著提高夏玉米籽粒产量,其中,2018年度夏玉米季籽粒产量BCL、BCM和BCH生物炭处理籽粒产量分别较BC0增6.4%、17.9%、15.4%,其中,BCM处理达到最高值10 811 kg·hm-2,但BCM处理与BCH处理间无显著差异。此外,通过对各处理连续7年度(2011—2018)夏玉米季平均籽粒产量的影响比较发现,各生物炭处理也均显著高于对照BC0处理,但各生物炭处理间差异不显著。图2
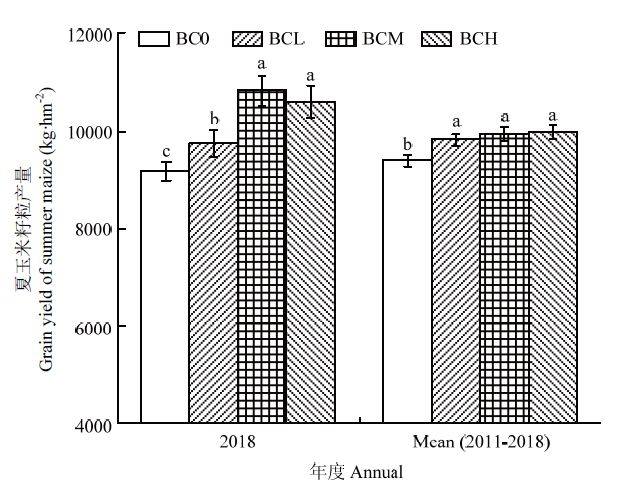 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2生物炭施用对玉米籽粒产量的影响
BC0、BCL、BCM、BCH分别代表对照、低量生物炭、中量生物炭和高量生物炭4种处理。相同年度不同处理图柱上不同字母表示处理间存在显著差异(P<0.05);柱形图顶部“±”代表标准误(n=3)。下同
Fig. 2Effects of biochar addition on grain yield of summer maize
BC0, BCL, BCM and BCH represent control, low, medium and high rate of biochar treatments, respectively. Different letters in the same annual indicate significant differences between treatments (P<0.05). Error bars in column top indicate standard errors (n=3). The same as below
2.2 施用生物炭对土壤无机氮含量的影响
由图3看出,各处理土壤无机氮(Nmin)含量整体变化趋势基本相同,均表现为在施肥后或者灌水(或降雨)后含量有所上升,且添加生物炭处理Nmin含量明显较未添加生物炭对照处理高。自2018年6月9日玉米播种至8月1日各处理Nmin含量波动均较大,7月25日(玉米大喇叭口期)施肥后土壤Nmin持续上升,施肥4 d后(7月29日)达到峰值,之后逐渐下降,8月20日(玉米灌浆期)降到最低值,之后趋于平缓。其中,在夏玉米成熟期(9月13日),BCL、BCM和BCH处理的土壤Nmin含量显著高于BC0处理,具体表现为BCH处理>BCM处理>BCL处理>BC0处理。该结果表明,连续多年添加生物炭具有提高农田无机氮含量的效果。图3
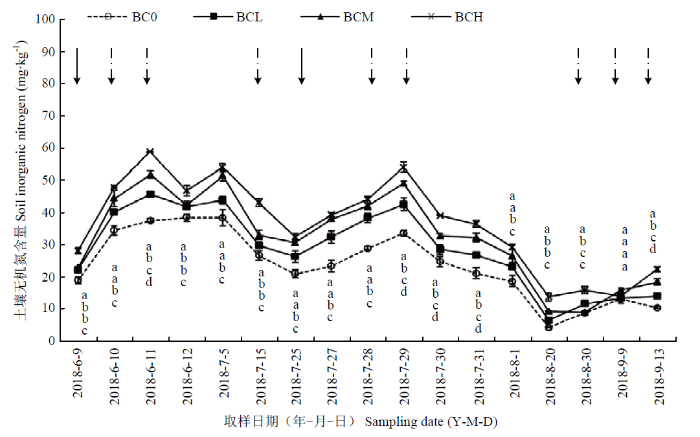 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同生物炭处理土壤耕层(0—20 cm)无机氮(Nmin)含量动态变化
实线箭头所指代表施肥时期,虚线箭头所指代表灌溉或降雨时期;同一取样时期同列不同小写字母表示不同处理在P<0.05水平上差异显著。下同
Fig. 3Dynamic changes of Nmin content of topsoil (0-20 cm) in different biochar treatments
The arrow of solid line represents fertilization, and the arrow of dotted line represents irrigation or rainfall. Different lowercase letters within a column in the same sampling date indicate significantly differences at P<0.05 among different treatments. The same as below
2.3 施用生物炭对土壤N2O排放及含水量的影响
由图4可以看出,与土壤Nmin变化趋势相似,在整个玉米生育期土壤N2O排放通量变化趋势同样均表现为在施肥后或者灌水(或降雨)后含量有所上升,且在多数生育期生物炭处理土壤N2O排放通量显著低于对照BC0处理。在玉米大喇叭口追施氮肥时(7月25日)未添加生物炭对照BC0处理土壤N2O排放通量达到峰值456.45 μg·m-2·h-1,且显著高于生物炭处理。到夏玉米成熟期,与BC0相比,3个生物炭处理显著降低土壤N2O排放通量,并以BCM处理减少N2O排放幅度较大,具体表现为BC0处理>BCL处理>BCH处理>BCM处理。该研究表明,生物炭施用具有降低土壤N2O排放的效果,但并非生物炭添加量越多效果越佳。图4
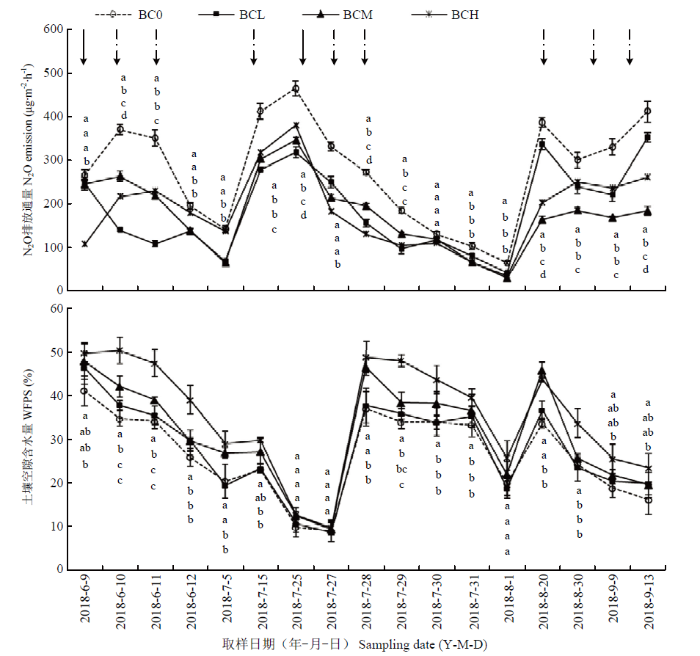 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同生物炭处理土壤耕层(0—20 cm)N2O排放通量和水分含量动态变化
Fig. 4Dynamic changes of N2O emission and moisture of topsoil (0-20 cm) in different biochar treatments
由图4还可以看出,各处理土壤空隙含水量(WFPS)变化趋势基本一致,均在施肥灌水后(6月9日和7月27日)和有效降雨后(8月20日)先升高后逐渐降低,且在多数监测时期中量和高量生物炭处理(BCM和BCH)土壤WFPS显著高于对照BC0处理。
此外,从图5也可看出,不同处理在2018年整个夏玉米生育期和2011—2018年连续7个夏玉米季具有相似的土壤N2O累积排放规律,均表现为BCH、BCM处理显著低于BC0处理,但BCL与BC0处理间差异不显著。其中,在2018年玉米季,BCM和BCH处理土壤N2O累积排放量分别较BC0降低31.3%和16.9%,并以BCM处理降低幅度最大。该研究结果表明,长期连续施用生物炭具有降低农田土壤N2O排放的效果。
图5
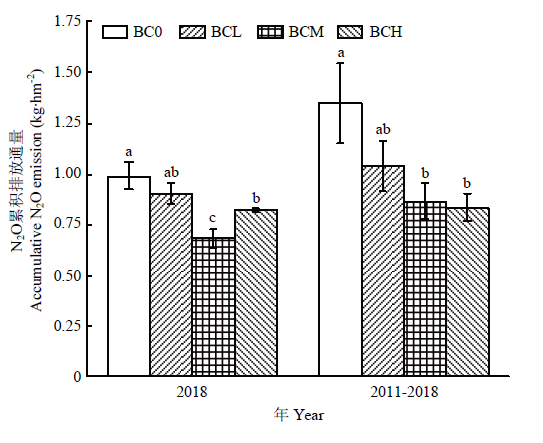 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5不同处理玉米生育期N2O累积排放量
Fig. 5N2O accumulation emission of different biochar treatments in maize season
2.4 施用生物炭对土壤AOA和AOB基因丰度的影响
图6表明,土壤AOA基因拷贝数均显著高于AOB基因。对于土壤AOA基因而言,在夏玉米大喇叭口期和成熟期均表现为随生物炭施用量增加呈现先上升后下降的趋势,且在该两个生育期均以BCM处理基因拷贝数较高。对于土壤AOB基因,大喇叭口期AOB基因拷贝数则表现为随生物炭施用量的增加呈现持续上升趋势,其中BCM和BCH处理分别较BC0显著增加101.5%和421.2%。与土壤AOA变化趋势相似,成熟期土壤AOB基因拷贝数也随生物炭施用量增加呈现先上升后下降趋势,且BCM和BCH处理也显著高于BC0处理,并以BCM处理最高。图6
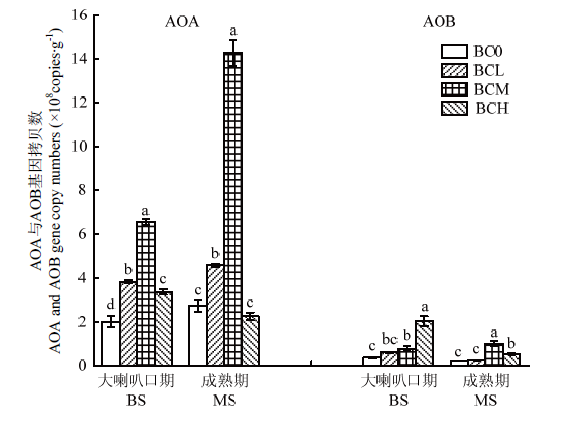 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6生物炭对土壤AOA和AOB基因丰度的影响
BS:大喇叭口期;MS:成熟期。下同
Fig. 6Effects of biochar amendment on the abundances of AOA and AOB in topsoil
BS: Bellmouth stage; MS: Maturity stage. The same as below
2.5 施用生物炭对土壤nirK、nirS和nosZ基因丰度的影响
图7表明,各处理土壤nirS基因拷贝数均远高于nirK和nosZ基因拷贝数,前者比后者高出1—3个数量级。土壤nirK基因拷贝数在夏玉米大喇叭口期和成熟期均表现为随着生物炭用量增加呈现先上升后下降的趋势,且在该两生育期均以BCM处理数值最高。对于土壤nirS基因拷贝数,与BC0比较,大喇叭口期BCL、BCM和BCH处理分别显著增加33.9%、31.9%和62.0%,成熟期BCM和BCH处理分别显著提高40.4%和32.4%。而对于土壤nosZ基因,在夏玉米大喇叭口期和成熟期则表现出不同的变化规律,大喇叭口期,BCL、BCM和BCH处理土壤nosZ基因拷贝数分别较BC0显著增加25.3%、34.5%和127.5%;成熟期,土壤nosZ基因拷贝数随着生物炭施用量的增加则同样呈现先上升后下降的趋势,并以BCM处理数值最高。图7
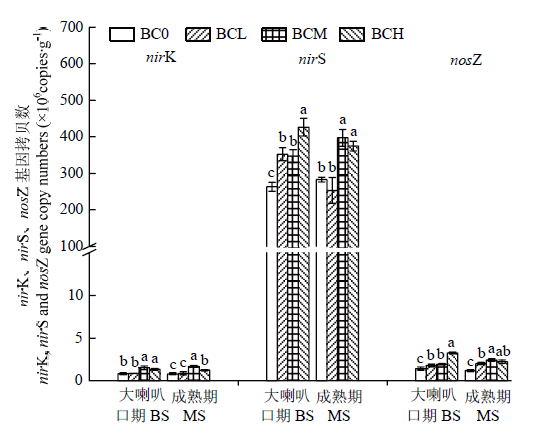 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7生物炭对土壤nirK、nirS和nosZ基因丰度的影响
Fig. 7Effects of biochar amendment on the abundances of nirK, nirS and nosZ in topsoil
2.6 土壤N2O排放与各氮循环相关功能基因丰度的相关性分析
对成熟期土壤理化性质、N2O排放与各功能基因丰度进行相关性分析,由表2可知,N2O排放通量与土壤硝态氮、土壤含水量、AOA呈显著负相关关系(P<0.05),与AOB、nirK、nirS、nosZ呈极显著负相关关系(P<0.01)。土壤铵态氮含量与其他理化指标和基因拷贝数均未达到显著性水平(P>0.05),土壤硝态氮含量与AOA和AOB呈现极显著正相关关系(P<0.01),与土壤nirK呈现显著正相关关系(P<0.05);土壤空隙含水量(WFPS)与nirS、nosZ呈显著正相关关系(P<0.05);土壤AOA与AOB和nirK呈现极显著正相关关系(P<0.01);土壤AOB与nirK、nirS、nosZ呈极显著正相关关系(P<0.01);土壤nirK与nirS呈极显著负相关(P<0.01),与nosZ显著负相关(P<0.05);土壤nirS与nosZ呈显著负相关关系(P<0.05)。Table 2
表2
表2土壤性质、N2O排放与功能基因丰度的相关性分析
Table 2
| N2O | NH4+ | NO3- | WFPS | AOA | AOB | nirK | nirS | nosZ | |
|---|---|---|---|---|---|---|---|---|---|
| N2O | 1 | -0.098 | -0.611* | -0.645* | -0.598* | -0.866** | -0.863** | -0.915** | -0.886** |
| NH4+ | 1 | 0.231 | -0.289 | 0.168 | 0.248 | 0.167 | 0.231 | -0.18 | |
| NO3- | 1 | 0.121 | 0.764** | 0.731** | 0.651* | 0.564 | 0.512 | ||
| WFPS | 1 | 0.007 | 0.383 | 0.349 | 0.610* | 0.645* | |||
| AOA | 1 | 0.846** | 0.796** | 0.535 | 0.568 | ||||
| AOB | 1 | 0.960** | 0.821** | 0.720** | |||||
| nirK | 1 | 0.810** | 0.706* | ||||||
| nirS | 1 | 0.664* | |||||||
| nosZ | . | 1 |
新窗口打开|下载CSV
3 讨论
3.1 施用生物炭对作物产量的影响
关于生物炭对作物产量的影响,国内外有不同报道,但总体上以正向效应居多。UZOMA等[36]研究发现,在砂质土壤中施用生物炭,可显著提高玉米产量,王江伟等[37]发现,施用生物炭能够提高稻麦籽粒产量11.4%—60.5%。本研究发现,连续施用生物炭6年后可显著提高潮土农田夏玉米籽粒产量,且以6.75 t·hm-2生物炭用量处理增产效果较好。此外,本研究还发现,在玉米成熟期,施用生物炭可显著提高土壤Nmin,表明连续施用秸秆炭可以提升土壤无机氮库。生物炭较大的比表面积和丰富的微孔结构可增强土壤保墒能力[38]。本研究发现,在多数取样时期,添加生物炭土壤含水量普遍显著高于未添加生物炭对照处理,表明施用生物炭具有提高土壤有效水含量的效果,可对作物生长产生积极影响。因此,本研究分析认为,长期连续添加生物炭提高土壤Nmin含量和土壤含水量可能是促进作物产量提高的主要因素。3.2 施用生物炭对N2O排放与氮功能基因丰度的影响
目前,关于生物炭对土壤N2O排放影响的研究较多,但结果不一,且其作用机制各研究者的观点也不尽相同。有研究认为,在我国北方旱地土壤,硝化过程是N2O的主要来源且主导了N2O排放,反硝化作用产生的N2O较少[39,40];而在菜地土壤中,施用生物炭可显著提高土壤pH和SOC,增加amoA、nirK和nirS基因丰度从而间接促进N2O排放,同时增加 nosZ基因丰度直接促进N2O还原,综合效应表现为降低菜地土壤N2O排放[41]。关于AOA和AOB对土壤硝化作用影响N2O排放的贡献存在很大争议。有研究发现,当土壤AOA占据土壤硝化作用相关功能基因主导地位时,施用生物炭会抑制氨氧化作用[42]。ZHANG等[43]研究发现,酸性土壤AOA主导N2O排放,而碱性土壤N2O则由AOB主导。WU等[44]研究发现,新鲜生物炭和老化生物炭均可提高稻田土壤中AOA和AOB基因丰度,降低(nirS+nirK)/nosZ比值,但对N2O的季节性排放没有明显的影响。本研究中,施用生物炭可显著提高潮土农田(碱性土壤)夏玉米成熟期AOA和AOB基因丰度,这一发现与前人研究结果一致[45],但在本研究中施用生物炭可降低潮土农田N2O排放。相关分析也表明,土壤AOA和AOB基因丰度与N2O排放量呈显著负相关,说明在本试验中土壤AOA和AOB基因丰度的增加可能引起amoA基因丰度的增加促进了土壤硝化作用的氨氧化过程,从而引起NO3-含量增加,最终导致N2O排放量降低。同时,土壤NO3-含量与AOA和AOB呈显著正相关,而与N2O排放呈显著负相关,也印证了本研究结果。但仅仅对比基因丰度的前后变化不足以说明氨氧化微生物对生物炭的响应机制,未来针对土壤中AOA和AOB群落结构变化,及其如何影响氨氧化过程仍需进一步研究。贺超卉等[46]研究发现,向土壤中施用少量生物炭可促进N2O向N2的转化过程,增加nirS和nosZ反硝化基因丰度。WANG等[47]发现,土壤nirS和nosZ基因在控制N2O排放中起关键作用。HARTER等[48]研究表明,在中性和碱性土壤中N2O排放量主要受nosZ基因调控,而nosZ基因在反硝化作用中的主要功能是编码氧化亚氮还原酶,将N2O催化还原为N2,其基因丰度增加有助于促进反硝化作用过程,从而导致N2O排放量降低,这也部分解释了本文研究结果。与前人研究类似,在本研究发现,土壤N2O排放与nirK、nirS、nosZ呈极显著负相关关系,施用生物炭可显著提高土壤nosZ基因丰度,进而导致N2O排放量降低。
综上所述,本研究通过长期定位试验发现,在黄淮潮土农田连续多年施用生物炭可以更好的吸附储存水分及无机氮素养分,为土壤微生物提供良好的栖息环境,进而提高土壤的硝化作用和反硝化作用,多因素协同作用最终降低土壤N2O排放量。然而,由于土壤微生物种类及群落结构的复杂性,不同类型土壤对生物炭的响应机制也不尽相同,关于生物炭如何调控土壤硝化反硝化相关微生物功能基因丰度来减少N2O排放仍需更深入研究。另外,截至目前,本生物炭长期定位试验已经进行接近8周年,而有关生物炭在研究地区土壤连续施用作物所能承受的饱和点(或阈值)仍未发现,今后仍需继续观测,生物炭施用达到阈值后对作物产量和温室气体减排的影响也是今后需要重点关注的方向。
4 结论
生物炭在保墒增产、养分储存以及N2O减排效应方面具有长期累积效应。土壤保墒能力和农田无机氮含量提高可能是施用生物炭促进夏玉米产量提高的主要原因。连续多年施用生物炭通过促进参与土壤N2O排放的硝化作用(AOA、AOB)和反硝化作用(nosZ)相关功能基因丰度的提高,改变了土壤氮素循环状况,进而起到降低潮土农田土壤N2O排放的效果,但过量施用生物炭(11.25 t·hm-2),土壤N2O排放量则略有增加。总体而言,中等生物炭用量(6.75 t·hm-2)对实现潮土农田夏玉米籽粒产量提高和土壤N2O减排效果最优。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:21807245 [本文引用: 1]
DOI:10.1126/science.1186120URLPMID:20929768 [本文引用: 1]

Atmospheric reactions and slow geological processes controlled Earth's earliest nitrogen cycle, and by ~2.7 billion years ago, a linked suite of microbial processes evolved to form the modern nitrogen cycle with robust natural feedbacks and controls. Over the past century, however, the development of new agricultural practices to satisfy a growing global demand for food has drastically disrupted the nitrogen cycle. This has led to extensive eutrophication of fresh waters and coastal zones as well as increased inventories of the potent greenhouse gas nitrous oxide (N(2)O). Microbial processes will ultimately restore balance to the nitrogen cycle, but the damage done by humans to the nitrogen economy of the planet will persist for decades, possibly centuries, if active intervention and careful management strategies are not initiated.
DOI:10.1098/rstb.2011.0415URLPMID:22451101 [本文引用: 1]
Nitrous oxide (N(2)O) is a powerful atmospheric greenhouse gas and cause of ozone layer depletion. Global emissions continue to rise. More than two-thirds of these emissions arise from bacterial and fungal denitrification and nitrification processes in soils, largely as a result of the application of nitrogenous fertilizers. This article summarizes the outcomes of an interdisciplinary meeting, 'Nitrous oxide (N(2)O) the forgotten greenhouse gas', held at the Kavli Royal Society International Centre, from 23 to 24 May 2011. It provides an introduction and background to the nature of the problem, and summarizes the conclusions reached regarding the biological sources and sinks of N(2)O in oceans, soils and wastewaters, and discusses the genetic regulation and molecular details of the enzymes responsible. Techniques for providing global and local N(2)O budgets are discussed. The findings of the meeting are drawn together in a review of strategies for mitigating N(2)O emissions, under three headings, namely: (i) managing soil chemistry and microbiology, (ii) engineering crop plants to fix nitrogen, and (iii) sustainable agricultural intensification.
[D].
[本文引用: 1]
[D].
[本文引用: 1]
DOI:10.1007/s11104-007-9391-5URL [本文引用: 1]
[本文引用: 1]
DOI:10.3864/j.issn.0578-1752.2019.07.013URL [本文引用: 1]
【目的】氨氧化作用是硝化作用的第一步,也是硝化作用的限速步骤,是全球氮循环的关键环节。本试验旨在研究在我国不同类型土壤中添加花生壳生物炭对细菌和氨氧化作用的影响,为生物炭的推广使用提供理论依据。【方法】试验以黄棕壤、潮土、黑土为供试土壤,通过短期培养试验,利用16SrRNA测序研究生物炭对不同类型土壤氨氧化微生物、细菌群落结构以及相关酶基因表达量的影响。每种土壤设置4个处理:CK(不施用化肥和生物炭),F(单施化肥),C(单施2%花生壳生物炭),FC(施用化肥+2%花生壳生物炭)。【结果】施用生物炭后(C、FC)酸性土壤pH显著提高了0.5—1.0个单位,但碱性土壤pH显著降低了0.5—0.6个单位;单施生物炭(C)造成黄棕壤的微生物丰富度和多样性显著提高,潮土在单施生物炭(C)时仅显著提高了土壤的微生物多样性指数,在黑土中施用生物炭和化肥都未显著改变土壤微生物的丰富度和多样性;在3种土壤中氨氧化细菌的丰度皆高于氨氧化古菌,测得的氨氧化细菌的OTU丰度约为氨氧化古菌的8.1倍;生物炭和化肥并未显著改变奇古菌门中的OTU丰度,却对β和γ变形菌中的OTU丰度产生了显著性影响;3种土壤的氨氧化细菌都以β变形菌为主,约占60%;另外,生物炭的施用(C、FC)在PC1(40.4%)上显著改变了黄棕壤的微生物群落结构,在PC1(42.3%)和PC2(21.3%)上都显著改变了潮土的微生物群落结构;施用生物炭后(C、FC),短期内潮土中氨合成相关酶基因表达量显著降低14.7%—39.9%,氨氧化古菌丰度在单一施炭(C)和化肥与生物炭同施(FC)时分别降低了70.5%和48.7%。【结论】施用生物炭后,短期内显著改变了黄棕壤和潮土的微生物群落结构,并明显抑制了潮土的氨氧化作用。
DOI:10.3864/j.issn.0578-1752.2019.07.013URL [本文引用: 1]
【目的】氨氧化作用是硝化作用的第一步,也是硝化作用的限速步骤,是全球氮循环的关键环节。本试验旨在研究在我国不同类型土壤中添加花生壳生物炭对细菌和氨氧化作用的影响,为生物炭的推广使用提供理论依据。【方法】试验以黄棕壤、潮土、黑土为供试土壤,通过短期培养试验,利用16SrRNA测序研究生物炭对不同类型土壤氨氧化微生物、细菌群落结构以及相关酶基因表达量的影响。每种土壤设置4个处理:CK(不施用化肥和生物炭),F(单施化肥),C(单施2%花生壳生物炭),FC(施用化肥+2%花生壳生物炭)。【结果】施用生物炭后(C、FC)酸性土壤pH显著提高了0.5—1.0个单位,但碱性土壤pH显著降低了0.5—0.6个单位;单施生物炭(C)造成黄棕壤的微生物丰富度和多样性显著提高,潮土在单施生物炭(C)时仅显著提高了土壤的微生物多样性指数,在黑土中施用生物炭和化肥都未显著改变土壤微生物的丰富度和多样性;在3种土壤中氨氧化细菌的丰度皆高于氨氧化古菌,测得的氨氧化细菌的OTU丰度约为氨氧化古菌的8.1倍;生物炭和化肥并未显著改变奇古菌门中的OTU丰度,却对β和γ变形菌中的OTU丰度产生了显著性影响;3种土壤的氨氧化细菌都以β变形菌为主,约占60%;另外,生物炭的施用(C、FC)在PC1(40.4%)上显著改变了黄棕壤的微生物群落结构,在PC1(42.3%)和PC2(21.3%)上都显著改变了潮土的微生物群落结构;施用生物炭后(C、FC),短期内潮土中氨合成相关酶基因表达量显著降低14.7%—39.9%,氨氧化古菌丰度在单一施炭(C)和化肥与生物炭同施(FC)时分别降低了70.5%和48.7%。【结论】施用生物炭后,短期内显著改变了黄棕壤和潮土的微生物群落结构,并明显抑制了潮土的氨氧化作用。
DOI:10.1016/j.geoderma.2013.10.010URL [本文引用: 1]
DOI:10.1007/s00374-012-0745-7URL [本文引用: 1]
Biochar produced from plant biomass through pyrolysis has been shown to be much more resistant to biodegradation in the soil as compared with the raw biomass, such as cereal straw that is routinely shredded and discharged on to farm fields in large amounts. Biochar application to soil has also been reported to decrease greenhouse gas (GHG) emissions, although the mechanisms are not fully understood. In this study, the emissions of three main GHGs (CO2, CH4, and N2O) and enzyme activities (urease, beta-glycosidase, and dehydrogenase) were measured during a 100-day laboratory incubation of a Chernozemic soil amended with either straw or its biochar at rates of 0.67 and 1.68 % (based on the amount of C added) for the low and high rates, respectively. The biochar application dramatically reduced N2O emissions, but CO2 or CH4 emissions were not different, as compared with the un-amended soil. At the same C equivalent application rate, CO2 and N2O emission rates were greater while CH4 emission rates were lower in straw than in biochar application treatments. The activities of both the dehydrogenase and beta-glycosidase significantly declined while that of urease significantly increased with the biochar as compared with the straw treatment. We conclude that pyrolysis of cereal straw prior to land application would significantly reduce CO2 and N2O emissions, in association with changed enzyme activities, while increasing the soil C pool through the addition of stable C in the form of biochar.
DOI:10.3864/j.issn.0578-1752.2015.07.11URL [本文引用: 1]
【Objective】The various effects of different straw biochar on nutrient content and microbial community structure were studied in order to provide information for soil amelioration and proper management of straw residue.【Method】Through a 135-day laboratory incubation experiment and used a red paddy soil that originated from the Quaternary, the influences of rice and corn straw biochar that pyrolyzed at 300℃, 400℃ and 500℃ on soil pH, organic carbon, nutrient content, microbial biomass carbon and profile of microbial PLFAs community structure were investigated. The experiment consisted of seven treatments: control soil (CK), soil amended with 300℃ (RB300), 400℃ (RB400) and 500℃ (RB500) rice straw biochar, soil incorporated with 300℃ (CB300), 400℃ (CB400) and 500℃ (CB500) corn straw biochar. 【Result】 Feedstock type and pyrolysis temperature had a significant influence on the nutrient contents and chemical properties of biochar products. Compared with control, the two straw biochar amendments increased pH value by 0.16 unit and enhanced the contents of soil organic carbon, available P and available K by 26.1%, 20.6% and 281.8%, respectively. Under the same pyrolysis temperature, the application of rice straw biochar mainly promoted the level of available K while corn straw biochar improved the content of available P. Application of 300℃ straw biochar had no significant effect on soil available and mineral N contents. Compared with the control, soils amended with RB500 and CB500 were, respectively, 10.4% and 8.1% less in available N, while significantly increased by 63.6% and 100.7% in NO3--N concentration. Although the concentrations of microbial biomass carbon and total phospholipid fatty acids for soils amended with straw biochar were 63.4% and 47.5% higher than control soil, there was no significant difference between the control soil and soils with 300℃ straw biochar. Both the two types of biochar enhanced the contents of G-, G+, fungi and actinobacteria and shown as 300℃<400℃<500℃. Results of PCA indicated that rice straw biochar amendment had more effect on the structure of soil microbial community than corn straw biochar. The microbial community compositions of three rice straw biochar were separated from each other while no distinctive recognized among the three corn biochar. Results of CCA suggested that straw biochars can affect the composition of microbial community through altering soil chemical and nutrient properties, as soil available P, soil organic carbon and available had significant correlation with the distribution of soil microbial community. 【Conclusion】 Both the two straw biochars could ameliorate the acidity and nutrient content of red paddy soil, and enhance the level of soil microbial biomass. Soil microbial community structure had been affected in the presence of straw biochars and rice straw biochar had more effective influence than corn straw biochars.
DOI:10.3864/j.issn.0578-1752.2015.07.11URL [本文引用: 1]
【Objective】The various effects of different straw biochar on nutrient content and microbial community structure were studied in order to provide information for soil amelioration and proper management of straw residue.【Method】Through a 135-day laboratory incubation experiment and used a red paddy soil that originated from the Quaternary, the influences of rice and corn straw biochar that pyrolyzed at 300℃, 400℃ and 500℃ on soil pH, organic carbon, nutrient content, microbial biomass carbon and profile of microbial PLFAs community structure were investigated. The experiment consisted of seven treatments: control soil (CK), soil amended with 300℃ (RB300), 400℃ (RB400) and 500℃ (RB500) rice straw biochar, soil incorporated with 300℃ (CB300), 400℃ (CB400) and 500℃ (CB500) corn straw biochar. 【Result】 Feedstock type and pyrolysis temperature had a significant influence on the nutrient contents and chemical properties of biochar products. Compared with control, the two straw biochar amendments increased pH value by 0.16 unit and enhanced the contents of soil organic carbon, available P and available K by 26.1%, 20.6% and 281.8%, respectively. Under the same pyrolysis temperature, the application of rice straw biochar mainly promoted the level of available K while corn straw biochar improved the content of available P. Application of 300℃ straw biochar had no significant effect on soil available and mineral N contents. Compared with the control, soils amended with RB500 and CB500 were, respectively, 10.4% and 8.1% less in available N, while significantly increased by 63.6% and 100.7% in NO3--N concentration. Although the concentrations of microbial biomass carbon and total phospholipid fatty acids for soils amended with straw biochar were 63.4% and 47.5% higher than control soil, there was no significant difference between the control soil and soils with 300℃ straw biochar. Both the two types of biochar enhanced the contents of G-, G+, fungi and actinobacteria and shown as 300℃<400℃<500℃. Results of PCA indicated that rice straw biochar amendment had more effect on the structure of soil microbial community than corn straw biochar. The microbial community compositions of three rice straw biochar were separated from each other while no distinctive recognized among the three corn biochar. Results of CCA suggested that straw biochars can affect the composition of microbial community through altering soil chemical and nutrient properties, as soil available P, soil organic carbon and available had significant correlation with the distribution of soil microbial community. 【Conclusion】 Both the two straw biochars could ameliorate the acidity and nutrient content of red paddy soil, and enhance the level of soil microbial biomass. Soil microbial community structure had been affected in the presence of straw biochars and rice straw biochar had more effective influence than corn straw biochars.
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.agwat.2006.12.012URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1073/pnas.0506625102URLPMID:16186488
Nitrification, the microbial oxidation of ammonia to nitrite and nitrate, occurs in a wide variety of environments and plays a central role in the global nitrogen cycle. Catalyzed by the enzyme ammonia monooxygenase, the ability to oxidize ammonia was previously thought to be restricted to a few groups within the beta- and gamma-Proteobacteria. However, recent metagenomic studies have revealed the existence of unique ammonia monooxygenase alpha-subunit (amoA) genes derived from uncultivated, nonextremophilic Crenarchaeota. Here, we report molecular evidence for the widespread presence of ammonia-oxidizing archaea (AOA) in marine water columns and sediments. Using PCR primers designed to specifically target archaeal amoA, we find AOA to be pervasive in areas of the ocean that are critical for the global nitrogen cycle, including the base of the euphotic zone, suboxic water columns, and estuarine and coastal sediments. Diverse and distinct AOA communities are associated with each of these habitats, with little overlap between water columns and sediments. Within marine sediments, most AOA sequences are unique to individual sampling locations, whereas a small number of sequences are evidently cosmopolitan in distribution. Considering the abundance of nonextremophilic archaea in the ocean, our results suggest that AOA may play a significant, but previously unrecognized, role in the global nitrogen cycle.
DOI:10.1128/AEM.63.12.4704-4712.1997URLPMID:9406389
The naturally occurring genetic heterogeneity of autotrophic ammonia-oxidizing populations belonging to the beta subclass of the Proteobacteria was studied by using a newly developed PCR-based assay targeting a partial stretch of the gene which encodes the active-site polypeptide of ammonia monooxygenase (amoA). The PCR yielded a specific 491-bp fragment with all of the nitrifiers tested, but not with the homologous stretch of the particulate methane monooxygenase, a key enzyme of methane-oxidizing bacteria. The assay also specifically detected amoA in DNA extracted from various aquatic and terrestrial environments. The resulting PCR products retrieved from rice roots, activated sludge, a freshwater sample, and an enrichment culture were used for the generation of amoA gene libraries. No false positives were detected in a set of 47 randomly selected clone sequences that were analyzed further. The majority of the environmental sequences retrieved from rice roots and activated sludge grouped within the phylogenetic radiation defined by cultured strains of the genera Nitrosomonas and Nitrosospira. The comparative analysis identified members of both of these genera in activated sludge; however, only Nitrosospira-like sequences with very similar amino acid patterns were found on rice roots. Further differentiation of these molecular isolates was clearly possible on the nucleic acid level due to the accumulation of synonymous mutations, suggesting that several closely related but distinct Nitrosospira-like populations are the main colonizers of the rhizosphere of rice. Each of the amoA gene libraries obtained from the freshwater sample and the enrichment culture was dominated by a novel lineage that shared a branch with the Nitrosospira cluster but could not be assigned to any of the known pure cultures. Our data suggest that amoA represents a very powerful molecular tool for analyzing indigenous ammonia-oxidizing communities due to (i) its specificity, (ii) its fine-scale resolution of closely related populations, and (iii) the fact that a functional trait rather than a phylogenetic trait is detected.
DOI:10.1128/AEM.65.4.1652-1657.1999URLPMID:10103263
Using consensus regions in gene sequences encoding the two forms of nitrite reductase (Nir), a key enzyme in the denitrification pathway, we designed two sets of PCR primers to amplify cd1- and Cu-nir. The primers were evaluated by screening defined denitrifying strains, denitrifying isolates from wastewater treatment plants, and extracts from activated sludge. Sequence relationships of nir genes were also established. The cd1 primers were designed to amplify a 778 to 799-bp region of cd1-nir in the six published sequences. Likewise, the Cu primers amplified a 473-bp region in seven of the eight published Cu-nir sequences. Together, the two sets of PCR primers amplified nir genes in nine species within four genera, as well as in four of the seven sludge isolates. The primers did not amplify genes of nondenitrifying strains. The Cu primers amplified the expected fragment in all 13 sludge samples, but cd1-nir fragments were only obtained in five samples. PCR products of the expected sizes were verified as nir genes after hybridization to DNA probes, except in one case. The sequenced nir fragments were related to other nir sequences, demonstrating that the primers amplified the correct gene. The selected primer sites for Cu-nir were conserved, while broad-range primers targeting conserved regions of cd1-nir seem to be difficult to find. We also report on the existence of Cu-nir in Paracoccus denitrificans Pd1222.
DOI:10.1016/j.femsec.2004.04.011URLPMID:19712290
We re-evaluated PCR primers targeting nirS, nirK and nosZ genes for denaturing gradient gel electrophoresis as a tool to survey denitrifying community composition in environmental samples. New primers for both nirS and nosZ were combined with existing primers, while for nirK the previously published F1aCu:R3Cu set was chosen for denaturing electrophoresis. All three sets yielded amplicons smaller than 500 bp and amplified the correct fragment in all environmental samples. The denaturing gradient gel electrophoresis worked satisfactorily for nirK and nosZ, but not for nirS. This was probably due to the multiple melting domains in this particular nirS fragment. From the excised and sequenced bands, only sequences related to the target genes were detected and tree analysis showed that the selected primers acted as broad range primers for each of the three genes. By use of the new nirS primers it was demonstrated that agricultural soil harbours a substantial diversity of nirS denitrifiers.
DOI:10.1071/PP01071URL
DOI:10.1111/j.1475-2743.2011.00340.xURL [本文引用: 1]
In this study, we performed a greenhouse experiment to investigate the effect of cow manure biochar on maize yield, nutrient uptake and physico-chemical properties of a dryland sandy soil. Biochar was derived from dry cow manure pyrolysed at 500 degrees C. Cow manure biochar was mixed with a sandy soil at the rate equivalent to 0, 10, 15 and 20 t biochar per hectare. Maize was used as a test crop. Results of the study indicated that cow manure biochar contains some important plant nutrients which significantly affected the maize crop growth. Maize yield and nutrient uptake were significantly improved with increasing the biochar mixing rate. Application of biochar at 15 and 20 t/ha mixing rates significantly increased maize grain yield by 150 and 98% as compared with the control, respectively. Maize net water use efficiency (WUE) increased by 6, 139 and 91% as compared with the control, with the 10, 15 and 20 t/ha mixing rate, respectively. Nutrient uptake by maize grain was significantly increased with higher biochar applications. Application of cow manure biochar improved the field-saturated hydraulic conductivity of the sandy soil, as a result net WUE also increased. Results of the soil analysis after the harvesting indicated significant increase in the pH, total C, total N, Oslen-P, exchangeable cations and cation exchange capacity. The results of this study indicated that application of cow manure biochar to sandy soil is not only beneficial for crop growth but it also significantly improved the physico-chemical properties of the coarse soil.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.soilbio.2010.09.030URL [本文引用: 1]
DOI:10.1016/j.envpol.2010.10.040URLPMID:21251741 [本文引用: 1]
An automated system for continuous measurement of N(2)O fluxes on an hourly basis was employed to study N(2)O emissions in an intensively managed low carbon calcareous soil under sub-humid temperate monsoon conditions. N(2)O emissions occurred mainly within two weeks of application of NH(4)(+) based fertilizer and total N(2)O emissions in wheat (average 0.35 or 0.21 kg N ha(-)(1) season(-)(1)) and maize (average 1.47 or 0.49 kg N ha(-)(1) season(-)(1)) under conventional and optimum N fertilization (300 and 50-122 kg N ha(-)(1), respectively) were lower than previously reported from low frequency measurements. Results from closed static chamber showed that N(2)O was produced mainly from nitrification of NH(4)(+)-based fertilizer, with little denitrification occurring due to limited readily oxidizable carbon and low soil moisture despite consistently high soil nitrate-N concentrations. Significant reductions in N(2)O emissions can be achieved by optimizing fertilizer N rates, using nitrification inhibitors, or changing from NH(4)(+)- to NO(3)(-)-based fertilizers.
URL [本文引用: 1]
A 6-week incubation experiment with vegetable soils was conducted using real-time quantitative polymerase chain reaction (PCR) method to investigate the effects of biochar and organic manure on N2O emissions and functional marker genes of ammonia monooxygenase gene (amoA), nirK, nirS and nosZ, which are responsible for nitrification and denitrification. The field experiment included five treatments as following, CK:control, N:urea nitrogen fertilizer, N+BC:urea nitrogen fertilizer+biochar, N+M:urea nitrogen fertilizer+organic manure fertilizer and N+BC+M:urea nitrogen fertilizer+biochar+organic manure fertilizer. Results showed that compared with the CK treatment, the treatments receiving urea (N, N+BC, N+M and N+BC+M) greatly inhibited the copies of ammonia oxidizing bacteria (AOB) and ammonia oxidizing archaea (AOA), stimulated the copies of nirS, nirK and nosZ genes and increased N2O cumulative emissions throughout the incubation period. Compared with the N treatment, the N+BC treatment significantly increased soil pH by 11.1%, and simulated the copies of AOB, AOA, nirS, nirK and nosZ genes by 105.8%, 57.3%, 22.0%, 176.2% and 204.9%, respectively, thus decreased cumulative N2O emissions by 58.1%. Although it greatly increased nirK and nirS genes by 58.8% and 7.1%, respectively, the N+M treatment had no significant influence on N2O emissions as compared with the N treatment. The N+BC+M treatment greatly stimulated AOB, nirK, nirS and nosZ gene copies by 30.7%, 68.7%, 6.5% and 84.5%, respectively, and finally decreased cumulative N2O emissions by 14.4% as compared with the N treatment. Results suggested that biochar addition decreased overall N2O emissions by enhancing the expression of nosZ gene although it enhanced N2O production by stimulating the expression of amoA, nirK and nirS genes. In conclusion, the addition of biochar could serve as an appropriate practice for mitigating N2O emissions and increasing N-cycling microbial community under intensive vegetable fields. However, the addition of organic manure had little effects on N2O emission although it increased the gene abundance of denitrification microbes.
URL [本文引用: 1]
A 6-week incubation experiment with vegetable soils was conducted using real-time quantitative polymerase chain reaction (PCR) method to investigate the effects of biochar and organic manure on N2O emissions and functional marker genes of ammonia monooxygenase gene (amoA), nirK, nirS and nosZ, which are responsible for nitrification and denitrification. The field experiment included five treatments as following, CK:control, N:urea nitrogen fertilizer, N+BC:urea nitrogen fertilizer+biochar, N+M:urea nitrogen fertilizer+organic manure fertilizer and N+BC+M:urea nitrogen fertilizer+biochar+organic manure fertilizer. Results showed that compared with the CK treatment, the treatments receiving urea (N, N+BC, N+M and N+BC+M) greatly inhibited the copies of ammonia oxidizing bacteria (AOB) and ammonia oxidizing archaea (AOA), stimulated the copies of nirS, nirK and nosZ genes and increased N2O cumulative emissions throughout the incubation period. Compared with the N treatment, the N+BC treatment significantly increased soil pH by 11.1%, and simulated the copies of AOB, AOA, nirS, nirK and nosZ genes by 105.8%, 57.3%, 22.0%, 176.2% and 204.9%, respectively, thus decreased cumulative N2O emissions by 58.1%. Although it greatly increased nirK and nirS genes by 58.8% and 7.1%, respectively, the N+M treatment had no significant influence on N2O emissions as compared with the N treatment. The N+BC+M treatment greatly stimulated AOB, nirK, nirS and nosZ gene copies by 30.7%, 68.7%, 6.5% and 84.5%, respectively, and finally decreased cumulative N2O emissions by 14.4% as compared with the N treatment. Results suggested that biochar addition decreased overall N2O emissions by enhancing the expression of nosZ gene although it enhanced N2O production by stimulating the expression of amoA, nirK and nirS genes. In conclusion, the addition of biochar could serve as an appropriate practice for mitigating N2O emissions and increasing N-cycling microbial community under intensive vegetable fields. However, the addition of organic manure had little effects on N2O emission although it increased the gene abundance of denitrification microbes.
URL [本文引用: 1]
微生物是驱动土壤元素生物地球化学循环的引擎。氮循环是土壤生态系统元素循环的核心之一, 其四个主要过程, 即生物固氮作用、氨化作用、硝化作用、反硝化作用, 均由微生物所驱动。近10年来, 随着免培养的分子生态学技术和高通量测序技术等的发展, 在硝化微生物多样性及其作用机理、厌氧氨氧化过程和机理等研究方面取得了突破性进展。本文重点阐述了我国有关土壤硝化微生物方面的研究进展, 在此基础上, 简要介绍了反硝化微生物和厌氧氨氧化及硝酸盐异化还原成铵作用的研究进展, 并对今后的研究工作提出了展望。今后土壤氮素转化微生物生态学的研究, 应瞄准国际微生生态学发展的前沿, 加强新技术新方法的应用, 结合我国农业可持续发展、资源环境保护和全球变化研究的重大需求, 重点开展以下几方面的工作: (1) 开展大尺度上土壤硝化作用及氨氧化微生物分布的时空演变特征及驱动因子的研究; (2) 加强氮素转化关键微生物过程与机理的研究, 并与相关过程的通量(如氨挥发、N2O释放)和反应速率(如矿化速率、硝化速率)关联起来; (3) 在特定生态系统中系统研究各个氮转化过程的耦合关系, 构建相关氮素转化和氮素平衡模型, 为定向调控土壤氮素转化过程, 提高氮素利用效率并减少其负面效应提供科学依据。
URL [本文引用: 1]
微生物是驱动土壤元素生物地球化学循环的引擎。氮循环是土壤生态系统元素循环的核心之一, 其四个主要过程, 即生物固氮作用、氨化作用、硝化作用、反硝化作用, 均由微生物所驱动。近10年来, 随着免培养的分子生态学技术和高通量测序技术等的发展, 在硝化微生物多样性及其作用机理、厌氧氨氧化过程和机理等研究方面取得了突破性进展。本文重点阐述了我国有关土壤硝化微生物方面的研究进展, 在此基础上, 简要介绍了反硝化微生物和厌氧氨氧化及硝酸盐异化还原成铵作用的研究进展, 并对今后的研究工作提出了展望。今后土壤氮素转化微生物生态学的研究, 应瞄准国际微生生态学发展的前沿, 加强新技术新方法的应用, 结合我国农业可持续发展、资源环境保护和全球变化研究的重大需求, 重点开展以下几方面的工作: (1) 开展大尺度上土壤硝化作用及氨氧化微生物分布的时空演变特征及驱动因子的研究; (2) 加强氮素转化关键微生物过程与机理的研究, 并与相关过程的通量(如氨挥发、N2O释放)和反应速率(如矿化速率、硝化速率)关联起来; (3) 在特定生态系统中系统研究各个氮转化过程的耦合关系, 构建相关氮素转化和氮素平衡模型, 为定向调控土壤氮素转化过程, 提高氮素利用效率并减少其负面效应提供科学依据。
[本文引用: 1]
DOI:10.1007/s11356-018-3112-xURLPMID:30194577 [本文引用: 1]
Knowledge about the impacts of fresh and field-aged biochar amendments on greenhouse gas (CH4, N2O) emissions is limited. A field experiment was initiated in 2012 to study the effects of fresh and field-aged biochar additions on CH4 and N2O emissions and the associated microbial activity during the entire rice-growing season in typical rice-wheat rotation system in Southeast China. CH4 and N2O fluxes were monitored, and the abundance of methanogen (mcrA), methanotrophy (pmoA), ammonia-oxidizing archaea (AOA), ammonia-oxidizing bacteria (AOB), nitrite reductase (nirS, nirK), N2O reductase (nosZ), and potential soil enzyme activities related to CH4 and N2O were simultaneously measured throughout different rice developmental stages. There were three treatments: control (urea without biochar), fresh BC (urea with fresh biochar added in 2015), and aged BC (urea with 3-year field-aged biochar added in 2012). Results showed that field-aged biochar significantly decreased seasonal CH4 emissions by 16.8% in relation to the fresh biochar, though no significant differences were detected between biochars and control treatment. The structural equation model indicated that soil pH, microbial biomass carbon (MBC), pmoA, and mcrA were the main factors directly influenced by fresh and aged biochar amendments; aged biochar showed a negative effect while fresh biochar showed positive effects on CH4 fluxes. Both fresh and field-aged biochar obviously increased AOA and AOB abundances and reduced the (nirS+nirK)/nosZ ratio during the entire rice-growing season, although no significant effects were observed on seasonal N2O emissions. Therefore, biochar amendment produced long-term effects on total CH4 and N2O emissions through observed influences of soil pH and functional gene abundance. The figure shows how fresh and field-aged biochar differentially affected CH4 production and oxidation and N2O production and reduction through related functional gene abundances. Blue arrows indicate suppressing while pink arrows indicate promoting effect.
DOI:10.1111/j.1462-2920.2008.01578.xURLPMID:18336563 [本文引用: 1]
The abundance and composition of soil ammonia-oxidizing bacteria (AOB) and ammonia-oxidizing archaea (AOA) communities under different long-term (17 years) fertilization practices were investigated using real-time polymerase chain reaction and denaturing gradient gel electrophoresis (DGGE). A sandy loam with pH (H(2)O) ranging from 8.3 to 8.7 was sampled in years 2006 and 2007, including seven fertilization treatments of control without fertilizers (CK), those with combinations of fertilizer nitrogen (N), phosphorus (P) and potassium (K): NP, NK, PK and NPK, half chemical fertilizers NPK plus half organic manure (1/2OMN) and organic manure (OM). The highest bacterial amoA gene copy numbers were found in those treatments receiving N fertilizer. The archaeal amoA gene copy numbers ranging from 1.54 x 10(7) to 4.25 x 10(7) per gram of dry soil were significantly higher than those of bacterial amoA genes, ranging from 1.24 x 10(5) to 2.79 x 10(6) per gram of dry soil, which indicated a potential role of AOA in nitrification. Ammonia-oxidizing bacteria abundance had significant correlations with soil pH and potential nitrification rates. Denaturing gradient gel electrophoresis patterns revealed that the fertilization resulted in an obvious change of the AOB community, while no significant change of the AOA community was observed among different treatments. Phylogenetic analysis showed a dominance of Nitrosospira-like sequences, while three bands were affiliated with the Nitrosomonas genus. All AOA sequences fell within cluster S (soil origin) and cluster M (marine and sediment origin). These results suggest that long-term fertilization had a significant impact on AOB abundance and composition, while minimal on AOA in the alkaline soil.
[本文引用: 1]
[本文引用: 1]
DOI:10.1021/es305293hURLPMID:23745957 [本文引用: 1]
Although nitrous oxide (N2O) emissions from composting contribute to the accelerated greenhouse effect, it is difficult to implement practical methods to mitigate these emissions. In this study, the effects of biochar amendment during pig manure composting were investigated to evaluate the inter-relationships between N2O emission and the abundance of denitrifying bacteria. Analytical results from two pilot composting treatments with (PWSB, pig manure + wood chips + sawdust + biochar) or without (PWS, pig manure + wood chips + sawdust) biochar (3% w/w) demonstrated that biochar amendment not only lowered NO2(-)-N concentrations but also lowered the total N2O emissions from pig manure composting, especially during the later stages. Quantification of functional genes involved in denitrification and Spearman rank correlations matrix revealed that the N2O emission rates correlated with the abundance of nosZ, nirK, and nirS genes. Biochar-amended pig manure had a higher pH and a lower moisture content. Biochar amendment altered the abundance of denitrifying bacteria significantly; less N2O-producing and more N2O-consuming bacteria were present in the PWSB, and this significantly lowered N2O emissions in the maturation phase. Together, the results demonstrate that biochar amendment could be a novel greenhouse gas mitigation strategy during pig manure composting.
DOI:10.1016/j.scitotenv.2016.03.220URLPMID:27100017 [本文引用: 1]
Soil biochar amendment has been described as a promising tool to improve soil quality, sequester carbon, and mitigate nitrous oxide (N2O) emissions. N2O is a potent greenhouse gas. The main sources of N2O in soils are microbially-mediated nitrogen transformation processes such as nitrification and denitrification. While previous studies have focused on the link between N2O emission mitigation and the abundance and activity of N2O-reducing microorganisms in biochar-amended soils, the impact of biochar on the taxonomic composition of the nosZ gene carrying soil microbial community has not been subject of systematic study to date. We used 454 pyrosequencing in order to study the microbial diversity in biochar-amended and biochar-free soil microcosms. We sequenced bacterial 16S rRNA gene amplicons as well as fragments of common (typical) nosZ genes and the recently described 'atypical' nosZ genes. The aim was to describe biochar-induced shifts in general bacterial community diversity and taxonomic variations among the nosZ gene containing N2O-reducing microbial communities. While soil biochar amendment significantly altered the 16S rRNA gene-based community composition and structure, it also led to the development of distinct functional traits capable of N2O reduction containing typical and atypical nosZ genes related to nosZ genes found in Pseudomonas stutzeri and Pedobacter saltans, respectively. Our results showed that biochar amendment can affect the relative abundance and taxonomic composition of N2O-reducing functional microbial traits in soil. Thus these findings broaden our knowledge on the impact of biochar on soil microbial community composition and nitrogen cycling.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}