 ,山西省农业科学院农作物品种资源研究所/农业部黄土高原作物基因资源与种质创制重点实验室/杂粮种质资源发展与遗传改良山西省重点实验室,太原 030031
,山西省农业科学院农作物品种资源研究所/农业部黄土高原作物基因资源与种质创制重点实验室/杂粮种质资源发展与遗传改良山西省重点实验室,太原 030031Comprehensive Evaluation of Agronomic Traits and Quality Traits of Foxtail Millet Landrace in Shanxi
LIU SiChen, CAO XiaoNing, WEN QiFen, WANG HaiGang, TIAN Xiang, WANG JunJie, CHEN Ling, QIN HuiBin, WANG Lun, QIAO ZhiJun,1Institute of Crop Germplasm Resources, Shanxi Academy of Agricultural Sciences/Key Laboratory of Crop Gene Resources and Germplasm Enhancement of Loess Plateau, Ministry of Agriculture/Shanxi Key Laboratory of Genetic Resources and Genetic Improvement of Minor Crops, Taiyuan 030031通讯作者:
责任编辑: 李莉
收稿日期:2019-07-7接受日期:2020-01-13网络出版日期:2020-06-01
| 基金资助: |
Received:2019-07-7Accepted:2020-01-13Online:2020-06-01
作者简介 About authors
刘思辰,Tel:18235172361;E-mail: lsch209@163.com。

摘要
关键词:
Abstract
Keywords:
PDF (476KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
刘思辰, 曹晓宁, 温琪汾, 王海岗, 田翔, 王君杰, 陈凌, 秦慧彬, 王纶, 乔治军. 山西谷子地方品种农艺性状和品质性状的综合评价[J]. 中国农业科学, 2020, 53(11): 2137-2148 doi:10.3864/j.issn.0578-1752.2020.11.001
LIU SiChen, CAO XiaoNing, WEN QiFen, WANG HaiGang, TIAN Xiang, WANG JunJie, CHEN Ling, QIN HuiBin, WANG Lun, QIAO ZhiJun.
0 引言
【研究意义】谷子(Setaria italica)属于禾本科植物,古代称为“粟”,去壳后称为小米,是起源于中国的一种古老粮食作物,也是中国北方地区的主要粮食作物之一。谷子具有营养丰富均衡、抗旱节水、高光效、耐储存、粮草兼用的特点。中国谷子种植面积约200万hm2,约占全球谷子种植面积的80%,年总产量约350万t[1]。谷子中含有丰富的蛋白质、叶酸、维生素E、类胡萝卜素及硒,作为营养均衡作物对维持人体健康具有重要作用[2,3,4]。【前人研究进展】中国是谷子的一个遗传多样性中心,拥有谷子种质资源约2.7万余份,其中,地方品种超过2万份[5,6,7]。王晓娟等[8]对474份甘肃省谷子地方种质资源的22个性状进行了遗传多样性分析,结果表明,甘肃谷子地方种质资源各地区和各主要性状的多样性存在明显差异。闫锋等[9]对黑龙江省的谷子种质资源进行形态学上的遗传多样性分析,表明谷子材料具有广泛的形态变异。田伯红[10]对482份谷子地方品种和育成品种进行了遗传多样性分析,发现地方品种形态性状的多样性指数高于育成品种。王海岗等[11]选用来自不同国家和地区的878份谷子核心种质进行了综合鉴定和遗传多样性评估,结果表明,谷子资源的各性状均表现出丰富的变异。杨慧卿等[12]选用了来自国外及国内的68份分蘖型谷子进行遗传多样性分析,其数量性状遗传变异丰富。丁银灯等[13]对272份谷子育成品种、农家种及国外品种的主要农艺性状在新疆进行遗传多样性分析,发现不同地区谷子在新疆遗传变异丰富。谷子资源在形态性状方面具有较广泛的遗传多样性,地方品种形态性状的多样性指数大多高于育成品种。地方品种的广泛变异类型是现代谷子育种的重要遗传资源,重新评价和利用地方品种是拓展中国谷子育种遗传基础的重要途经[10]。主成分分析和聚类分析是种质资源遗传多样性分析中普遍采用的分析方法,已在多种作物种质资源评价中得到广泛应用[14,15,16,17,18,19]。山西省是中国谷子资源最丰富的地区之一,因其地形复杂气候多样,在漫长的农耕历史中,形成了丰富的谷子品种类型和繁多的谷子种质资源。1949年以来,山西省共进行了2次大规模的谷子资源征集,共收集谷子种质5 572份[20]。主要是地方品种和农家种,其中也有少量育成品种和一些珍贵稀有的优异种质。巨大的种质资源数量缺乏深入系统地研究,使得山西谷子地方品种利用效率比较低。温琪汾等[21]对山西谷子种质资源进行了表型鉴定、抗旱鉴定和抗黑穗病鉴定,从山西谷子地方品种中筛选出高抗黑穗病[22,23]和耐旱性强的资源[24,25]。王海岗等[26,27,28]开展山西谷子地方品种表型多样性和核心种质构建的研究,为山西谷子地方品种表型的统一鉴定奠定了基础。【本研究切入点】目前,山西谷子地方品种表型性状的多样性分析,主要集中在产量性状[29],对结合谷子籽粒品质性状综合评价分析的研究较少。【拟解决的关键问题】本研究旨在对山西谷子地方品种进行品质和农艺性状的综合鉴定,进一步了解山西谷子种质资源多样性的组成特点和分布状况及筛选出相对合理的表型评价指标,为山西谷子种质资源的评价和优异谷子新品种选育提供参考。1 材料与方法
1.1 试验材料
从山西谷子库存资源5 572份中选取212份山西谷子地方品种种质资源为材料,按地理来源从北到南取样,涵盖谷子基本的遗传多样性,其中,大同38份、朔州6份、阳泉12份、太原28份、榆次4份、晋中46份、临汾42份和运城36份。1.2 试验方法
2015—2016年在山西省农业科学院东阳试验基地对212份山西谷子资源12个农艺性状(株高、穗长、茎长、茎粗、穗粗、节数、码数、码粒数、单穗重、穗粒重、千粒重和粒色)进行调查。每份材料种2行,行长2.5 cm,依据《谷子种质资源描述规范和数据标准》[30],粒色各性状差别用数字表示,1=白色,2=黄色,3=红色,4=青色,5=褐色,6=黑色。籽粒脱壳磨粉,进行3个品质性状(蛋白质、粗淀粉和直链淀粉与支链淀粉的比值)的测定。参考GB/T5511—2008《谷物和豆类氮含量测定和粗蛋白质含量计算凯氏法》测定谷子蛋白质含量,利用全自动凯氏定氮仪测定粗蛋白质测定2次,2次测定的相对误差小于1.0%。参照GB5006—1985《谷物籽粒粗淀粉测定法》测定谷子淀粉含量,在室温下用旋光仪测定,计算粗淀粉百分含量,2次测定的相对误差小于1.0%。直/支比测定:参考水稻的国标碘比色法直链淀粉含量测定法,通过单波长比色法测定谷子中直链淀粉含量。由谷子中直链淀粉和总淀粉中支链淀粉的比例获得直/支比[31]。
1.3 数据分析
采用Microsoft Excel 2003和DPS7.05统计分析数据。采用DPS7.05进行聚类分析、相关性分析和主成分分析,并计算各主成分及表型性状的综合得分F值,再结合逐步回归分析筛选谷子资源综合性状评价指标。1.3.1 隶属函数分析 采用模糊隶属函数计算出各性状的隶属函数值,将各性状定义到[0,1]闭区间。
Μ(Xi)=(Xi-Xi min)/(Xi max-Xi min)(i=1,2,3,...,n)
式中,Μ(Xi)为某种质材料第i个性状的隶属函数值,Xi为某种质材料第i个性状值,Xi max和Xi min分别为所有种质材料第i个性状的最大值和最小值。
1.3.2 遗传多样性指数 通过隶属函数值得到各性状每一级别的相对频率,再采用Shannon-Wiener’s多样性指数(Shannon-Wiener diversity index,H′)进行遗传多样性评价。
${H}'=-\sum\limits_{i=1}^{n}{P{}_{i}\times\ln {{p}_{i}}(i=1,2,3,...,n)}$
式中,pi表示某性状第i级别内材料份数占总份数的百分比。
1.3.3 种质资源的稳定性评价 采用Origin 2018绘制2015—2016年不同性状的boxplot图。
2 结果
2.1 表型性状的遗传多样性分析
山西谷子地方种质资源不同材料之间存在较大差异,相同性状在不同材料间表现出不同程度的多样性。通过对15个性状进行遗传多样性分析(表1),结果表明,除粒色外,遗传多样性指数均大于1.00,其中,株高(2.15)最高。遗传多样性指数排序为:株高>码数>单穗重>茎长>穗长>蛋白质>淀粉>节数>千粒重>穗粒重>码粒数>穗粗>茎粗>直/支比>粒色。株高、码数、茎长等表型性状的多样性指数较高,而直/支比和粒色的多样性指数较低。Table 1
表1
表1212份山西谷子资源15个性状变化
Table 1
| 性状Trait | 平均值Mean±SD | 变幅Range | 变异系数CV (%) | 多样性指数H' |
|---|---|---|---|---|
| 株高PH (cm) | 120.14±16.35 | 63.50-158.67 | 13.61 | 2.15 |
| 穗长PL (cm) | 29.83±5.47 | 16.67-49.00 | 18.34 | 1.94 |
| 茎长SL (cm) | 31.55±4.63 | 21.28-46.00 | 14.66 | 1.95 |
| 茎粗SD (cm) | 0.77±0.11 | 0.52-1.12 | 14.56 | 1.10 |
| 穗粗PD (cm) | 2.81±0.54 | 1.25-4.27 | 19.19 | 1.31 |
| 节数SNN | 13.17±1.35 | 9.00-18.50 | 10.23 | 1.69 |
| 码数SN | 110.93±24.18 | 57.00-176.50 | 21.80 | 2.08 |
| 码粒数GNS | 126.32±24.18 | 54.00-238.00 | 28.17 | 1.50 |
| 单穗重SSW (g) | 32.29±7.99 | 13.23-55.89 | 24.74 | 1.97 |
| 穗粒重SGW (g) | 25.61±6.88 | 9.91-47.16 | 26.87 | 1.57 |
| 千粒重TGW (g) | 3.50±0.42 | 1.87-4.58 | 12.04 | 1.57 |
| 蛋白质Protein (%) | 12.60±1.07 | 10.34-15.87 | 8.45 | 1.82 |
| 淀粉Starch (%) | 72.63±2.44 | 51.35-76.31 | 3.35 | 1.71 |
| 直/支比Amylase/Amylopectin ratio | 0.26±0.05 | 0.10-0.39 | 17.47 | 1.07 |
| 粒色GC | - | - | 38.66 | 0.92 |
新窗口打开|下载CSV
不同材料的变异系数存在较大差异,为3.35%—38.66%。蛋白质和淀粉的变异系数较低,粒色、码粒数、穗粒重、单穗重、码数、穗粗、穗长和直/支比的变异系数较高。其中,直/支比最高达0.39,最低仅为0.10,变异系数为17.47%,而淀粉变异系数最低,仅为3.35%,这对谷子新品种选育,尤其对品质育种具有参考价值。
2.2 农艺性状的年度变化
在同一生态环境下,2015—2016年进行谷子资源农艺性状的调查(表2),2015年,码粒数、单穗重和穗粒重表现出较高的变异系数,而在2016年,穗长、茎长、茎粗、穗粗、节数、码数和千粒重表现出较高的变异系数,说明表型性状受环境影响大。结合2年11个农艺性状平均值的boxplot图(图1)可以看出,株高、码数、码粒数、单穗重和穗粒重年度间变化大,穗长、茎长、茎粗、穗粗、节数和千粒重受环境影响小。Table 2
表2
表22015—2016年主要农艺性状年度间变异
Table 2
| 性状 Trait | 2015 | 2016 | ||
|---|---|---|---|---|
| 平均值Mean±SD | 变异系数CV (%) | 平均值Mean±SD | 变异系数CV (%) | |
| 株高PH (cm) | 113.89±17.52 | 15.38 | 126.40±18.35 | 14.52 |
| 穗长PL (cm) | 30.08±5.59 | 18.59 | 29.59±6.23 | 21.06 |
| 茎长SL (cm) | 33.08±6.27 | 18.96 | 31.73±6.75 | 21.26 |
| 茎粗SD (cm) | 0.80±0.12 | 15.28 | 0.74±0.17 | 22.23 |
| 穗粗PD (cm) | 2.71±0.59 | 21.60 | 2.91±0.75 | 25.73 |
| 节数SNN | 13.06±1.38 | 10.54 | 13.29±1.54 | 11.60 |
| 码数SN | 102.23±21.27 | 20.81 | 119.62±30.93 | 25.86 |
| 码粒数GNS | 122.54±46.52 | 37.96 | 130.10±38.83 | 29.85 |
| 单穗重SSW (g) | 28.01±8.53 | 30.43 | 36.57±10.54 | 28.81 |
| 穗粒重SGW (g) | 23.42±8.67 | 36.99 | 27.79±8.41 | 30.27 |
| 千粒重TGW (g) | 3.39±0.45 | 13.31 | 3.61±0.53 | 14.56 |
新窗口打开|下载CSV
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图12015—2016年在东阳基地11个农艺性状的boxplot图
盒图两端表示性状的极值范围;□:平均值;中间直线:中位线;◆:个别极值。PH:株高;PL:穗长;SL:茎长;SD:茎粗;PD:穗粗;SNN:节数;SN:码数;GNS:码粒数;SSW:单穗重;SGW:穗粒重;TGW:千粒重
Fig. 1The boxplot of eleven agronomic traits in Dongyang from 2015-2016
Both ends of boxplot indicate the extreme range of traits; □: The mean of the trait; The lines in the middle: The median line; ◆: Individual extremum. PH: Plant height; PL: Panicle length; SL: Stem length; SD: Stem diameter; PD: Panicle diameter; SNN: Stem node number; SN: Spike number; GNS: Grain number per spike; SSW: Single spike weight; SGW: Spike grain weight; TGW: Thousand-grain weight
2.3 相关性分析
由表3表明,15个性状间存在不同程度的相关性。株高与节数、码数和码粒数呈极显著正相关,与茎长和千粒重呈极显著负相关,与茎粗、穗粗和粒色相关性显著;穗长与茎长、茎粗、码数、单穗重、穗粒重和千粒重极显著相关;穗粗与茎粗、码粒数、单穗重、穗粒重和千粒重极显著正相关;节数与株高、茎粗、码数和码粒数呈极显著正相关;码数与码粒数呈极显著负相关;单穗重与穗长、茎长、茎粗、穗粗、码粒数、穗粒重和千粒重呈极显著正相关;千粒重与穗长、茎长、穗粗、单穗重和穗粒重呈极显著正相关,与淀粉和直/支比相关性显著;蛋白质与单穗重和穗粒重呈极显著负相关;淀粉与蛋白质呈极显著负相关;直/支比与穗粗呈极显著正相关;粒色与节数极显著负相关。Table 3
表3
表315个性状的相关性分析
Table 3
| 性状 Traits | 株高 PH | 穗长 PL | 茎长 SL | 茎粗 SD | 穗粗 PD | 节数 SNN | 码数 SN | 码粒数 GSN | 单穗重 SSW | 穗粒重 SGW | 千粒重 TGW | 蛋白质 Protein | 淀粉 Starch | 直/支比 Amylase/ Amylopectin ratio |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 穗长PL | -0.11 | |||||||||||||
| 茎长SL | -0.25** | 0.60** | ||||||||||||
| 茎粗SD | 0.15* | 0.33** | 0.04 | |||||||||||
| 穗粗PD | -0.18* | 0.16* | 0.14* | 0.19** | ||||||||||
| 节数SNN | 0.81** | -0.08 | -0.36** | 0.30** | -0.05 | |||||||||
| 码数SN | 0.39** | 0.40** | 0.22** | 0.21** | -0.32** | 0.29** | ||||||||
| 码粒数GSN | 0.18** | -0.02 | -0.05 | 0.29** | 0.42** | 0.27** | -0.29** | |||||||
| 单穗重SSW | 0.06 | 0.38** | 0.25** | 0.46** | 0.42** | 0.15* | 0.13 | 0.60** | ||||||
| 穗粒重SGW | -0.06 | 0.39** | 0.27** | 0.34** | 0.42** | 0 | 0.09 | 0.53** | 0.89** | |||||
| 千粒重TGW | -0.44** | 0.30** | 0.42** | -0.05 | 0.26** | -0.42** | -0.22** | 0 | 0.32** | 0.38** | ||||
| 蛋白质Protein | 0.06 | 0.08 | 0.15* | -0.06 | -0.17* | -0.01 | 0.15* | -0.17* | -0.20** | -0.26** | -0.14* | |||
| 淀粉Starch | -0.11 | -0.1 | 0 | -0.02 | 0.07 | -0.05 | -0.17* | 0.08 | 0.09 | 0.13 | 0.16* | -0.41** | ||
| 直/支比 Amylase/ Amylopectin ratio | -0.12 | 0.04 | 0.13 | -0.07 | 0.21** | -0.03 | -0.21** | 0.1 | 0.05 | 0.05 | 0.15* | -0.04 | -0.11 | |
| 粒色GC | -0.17* | 0.1 | 0.06 | -0.07 | -0.01 | -0.22** | 0.04 | -0.14* | 0.02 | 0.04 | 0.09 | 0.05 | 0 | -0.01 |
新窗口打开|下载CSV
2.4 主成分分析
对212份山西谷子地方品种的15个性状进行主成分分析(表4),第Ⅰ主成分贡献率为23.02%,第Ⅱ主成分贡献率为18.15%,第Ⅲ主成分贡献率为14.12%,第Ⅳ主成分贡献率为8.82%,第Ⅴ主成分贡献率为6.51%,第Ⅵ主成分贡献率为5.86%,第Ⅶ主成分贡献率为4.90%,第Ⅷ主成分贡献率为4.26%,第Ⅸ主成分贡献率为3.62%,9个主成分的累计贡献率为89.26%,因此,将原来的15个性状转化为9个新的独立的综合指标,用于谷子种质资源的表型性状评价。Table 4
表4
表4212份山西谷子资源性状的主成分分析
Table 4
| 性状Traits | Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | Ⅶ | Ⅷ | Ⅸ |
|---|---|---|---|---|---|---|---|---|---|
| 株高 PH | -0.1188 | 0.5085 | 0.0515 | 0.0099 | -0.0859 | 0.2071 | 0.3038 | 0.1324 | -0.1549 |
| 穗长PL | 0.2959 | -0.0209 | 0.4544 | -0.0099 | -0.119 | 0.0458 | -0.1833 | 0.1337 | -0.1555 |
| 茎长SL | 0.2564 | -0.2142 | 0.3829 | 0.044 | -0.2749 | 0.0018 | 0.1956 | 0.3161 | 0.0467 |
| 茎粗SD | 0.2361 | 0.2912 | 0.1097 | -0.0237 | 0.1128 | -0.2104 | -0.7207 | 0.1024 | 0.294 |
| 穗粗PD | 0.3319 | -0.0275 | -0.2258 | 0.2652 | 0.0842 | -0.059 | -0.1504 | 0.3252 | -0.724 |
| 节数SNN | -0.0614 | 0.5367 | -0.0392 | 0.039 | -0.0972 | 0.206 | 0.0573 | 0.1655 | -0.1175 |
| 码数SN | -0.023 | 0.2462 | 0.5179 | -0.2383 | -0.0843 | 0.173 | 0.0113 | -0.1526 | -0.1013 |
| 码粒数GSN | 0.2984 | 0.2511 | -0.3088 | 0.2056 | 0.1337 | -0.1389 | 0.2388 | 0.0895 | 0.2649 |
| 单穗重SSW | 0.4674 | 0.1951 | 0.0099 | -0.0345 | 0.1131 | -0.0081 | 0.175 | -0.1794 | 0.1386 |
| 穗粒重SGW | 0.4724 | 0.1055 | -0.0049 | -0.0882 | 0.0947 | 0.0013 | 0.2134 | -0.277 | 0.0825 |
| 千粒重TGW | 0.2999 | -0.3228 | 0.0188 | -0.064 | -0.1583 | 0.014 | 0.2151 | -0.1484 | -0.0567 |
| 蛋白质Protein | -0.1513 | -0.0143 | 0.3378 | 0.4467 | 0.1944 | -0.3336 | 0.2722 | 0.3972 | 0.2655 |
| 淀粉Starch | 0.0944 | -0.0664 | -0.2711 | -0.585 | -0.1982 | 0.0963 | 0.0436 | 0.6104 | 0.2353 |
| 直/支比Amylase/Amylopectin ratio | 0.0965 | -0.1058 | -0.1076 | 0.5063 | -0.3044 | 0.6684 | -0.1851 | 0.0029 | 0.2993 |
| 粒色GC | 0.0258 | -0.1693 | 0.1279 | -0.1236 | 0.7951 | 0.498 | 0.0252 | 0.1688 | 0.0089 |
| 特征值E | 3.4524 | 2.7222 | 2.1183 | 1.3234 | 0.9762 | 0.8787 | 0.7353 | 0.6384 | 0.5434 |
| 百分率CR (%) | 23.0161 | 18.1478 | 14.1222 | 8.8225 | 6.5083 | 5.8582 | 4.9021 | 4.2561 | 3.6226 |
| 累计百分率CCR (%) | 23.0161 | 41.1639 | 55.2861 | 64.1087 | 70.617 | 76.4752 | 81.3774 | 85.6335 | 89.256 |
新窗口打开|下载CSV
第Ⅰ主成分解释了15个性状23.02%的变化,特征向量绝对值最大的是穗粒重(0.47)和单穗重(0.47);第Ⅱ主成分特征向量绝对值最大的是节数(0.54)、株高(0.51)和千粒重(0.32);第Ⅲ主成分特征向量绝对值最大的是码数(0.52)、穗长(0.45)、茎长(0.38)和码粒数(0.31);第Ⅳ主成分特征向量绝对值最大的是蛋白质(0.45);第Ⅴ主成分特征向量绝对值最大的是粒色(0.80);第Ⅵ主成分特征向量绝对值最大的是直/支比(0.67);第Ⅶ主成分特征向量绝对值最大的是茎粗(0.72);第Ⅷ主成分特征向量绝对值最大的是淀粉(0.61);第Ⅸ主成分特征向量绝对值最大的是穗粗(0.72)。
2.5 综合评价
对15个表型性状值进行标准化处理,求得各种质的9个主成分得分,将9个主成分得分归一化处理,计算各主成分权重系数(0.258、0.203、0.158、0.099、0.073、0.066、0.055、0.048和0.041),最后求得每份种质材料的综合得分F值,进而对212份山西谷子种质资源进行综合评价。山西谷子的性状综合F值为0.521,排在前五位的分别是黄圪塔、大青谷、小穗谷、红小围谷和平遥谷。来自临汾的黄疙瘩F值(0.709)最高,来自大同的牛毛黄F值(0.315)最低,说明黄圪塔的综合性状最好。相关性分析表明(表5),除了千粒重和粒色外,其余10个农艺性状(株高、穗长、茎长、茎粗、穗粗、节数、码数、码粒数、单穗重和穗粒重)呈极显著正相关。F值和蛋白质无显著相关性,与淀粉含量呈显著负相关,与粒色呈显著正相关,与直/支比呈极显著正相关。利用综合得分F值和15个性状构建最优回归方程为Y=(-277.482+1.100Χ1 + 3.226Χ2+1.574Χ3+0.523Χ7+0.533Χ8+4.670Χ9+12.537Χ12+203.952Χ14 +17.694Χ15)×10-3,式中,Χ1、Χ2、Χ3、Χ7、Χ8、Χ9、Χ12、Χ14和Χ15分别代表株高、穗长、茎长、码数、码粒数、单穗重、蛋白质、直/支比和粒色。方程相关系数r和决定系数R2分别为0.9876和0.9754,表明9个自变量可决定F值总变异的97.54%,F值为891.3208,方差极显著。由回归方程可知9个性状对表型多样性综合值影响显著,可以作为山西谷子综合评价指标。Table 5
表5
表515个性状与表型综合得分(F值)的相关系数
Table 5
| 性状 Trait | 相关系数 Correlation coefficient | 性状 Trait | 相关系数 Correlation coefficient | |
|---|---|---|---|---|
| 株高PH | 0.32** | 单穗重SSW | 0.82** | |
| 穗长PL | 0.59** | 穗粒重SGW | 0.73** | |
| 茎长SL | 0.40** | 千粒重TGW | 0.14* | |
| 茎粗SD | 0.52** | 蛋白质Protein | 0.12 | |
| 穗粗PD | 0.31** | 淀粉Starch | -0.14* | |
| 节数SNN | 0.34** | 直/支比 Amylase/Amylopectin ratio | 0.18** | |
| 码数SN | 0.39** | 粒色GC | 0.15* | |
| 码粒数GNS | 0.53** |
新窗口打开|下载CSV
2.6 聚类分析
基于8个地区山西谷子15个性状的聚类分析,在欧式距离26.54将山西谷子资源划分为3类群(图2)。第一类群北部品种包括大同和朔州;第二类群中部品种包括阳泉、太原、晋中和榆次;第三类群南部品种包括临汾和运城。山西谷子地方品种的8个来源地,分别属于山西北部地区(大同和朔州)、南部地区(临汾和运城)和中部地区(太原、阳泉、晋中和榆次),说明山西谷子地方品种表型性状和地理来源一致。从表6可知,北部品种中穗长、穗粗、单穗重、穗粒重和千粒重等性状的平均值更高,株高、茎粗、穗粗、码数和穗粒重等性状的变异系数较高;南部品种株高、茎粗、节数、码数和蛋白质的平均值更高,码粒数、单穗重、千粒重、蛋白质、淀粉和直/支比的变异系数都高于中部品种和北部品种;中部品种中码粒数和直/支比的平均值较高,穗长、茎长、节数和粒色的变异系数高于南部品种和北部品种。图2
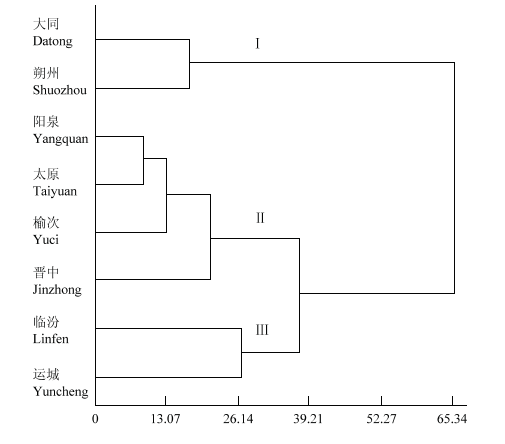 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2山西不同地区谷子资源的系统聚类图
Fig. 2Clustering dendrogram of foxtail millet germplasm from different areas of Shanxi
Table 6
表6
表6山西不同地区谷子资源15个性状的比较
Table 6
| 性状 Traits | 株高 PH (cm) | 穗长 PL (cm) | 茎长 SL (cm) | 茎粗 SD (cm) | 穗粗 PD (cm) | 节数 SNN | 码数 SN | 码粒数 GNS | 单穗重 SSW (g) | 穗粒重 SGW (g) | 千粒重 TGW (g) | 蛋白质 Protein (%) | 淀粉 Starch (%) | 直/支比 Amylase/ Amylopectin | 粒色 GC |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 山西北部North | |||||||||||||||
| 平均值±标准误 Mean±SD | 98.64± 10.94 | 31.73± 4.05 | 32.66± 3.15 | 0.78± 0.13 | 3.06± 0.58 | 11.70± 0.96 | 92.92± 17.98 | 125.34± 33.28 | 33.66± 8.01 | 29.06± 8.28 | 3.88± 0.32 | 12.12± 0.90 | 73.23± 1.54 | 0.26± 0.04 | 1.93± 0.38 |
| 范围 Range | 63.50- 116.84 | 26.84- 49.00 | 27.17- 43.67 | 0.53- 1.12 | 1.97- 4.22 | 9.00- 13.67 | 59.67- 151.00 | 72.33- 200.17 | 13.23- 51.65 | 9.91- 47.16 | 2.84- 4.58 | 10.44- 14.64 | 67.70- 75.68 | 0.14- 0.36 | 1.50- 3.50 |
| 变异系数 CV (%) | 11.10 | 12.77 | 9.63 | 16.29 | 18.92 | 8.23 | 19.35 | 26.55 | 23.79 | 28.49 | 8.20 | 7.44 | 2.11 | 14.65 | 19.81 |
| 山西中部Central | |||||||||||||||
| 平均值±标准误 Mean±SD | 124.02± 13.36 | 28.99± 5.75 | 32.25± 5.01 | 0.73± 0.09 | 2.91± 0.51 | 13.43± 1.22 | 107.51± 20.10 | 132.35± 34.82 | 32.88± 7.33 | 25.75± 5.65 | 3.53± 0.36 | 12.55± 0.98 | 73.10± 1.33 | 0.27± 0.04 | 1.92± 0.92 |
| 范围 Range | 77.20- 158.67 | 16.67- 45.17 | 22.78- 42.00 | 0.52- 1.00 | 1.25- 4.27 | 10.83- 18.50 | 58.50- 154.17 | 69.00- 207.00 | 16.91- 50.51 | 14.37- 39.19 | 2.67- 4.16 | 10.34- 14.74 | 69.11- 76.31 | 0.10- 0.39 | 1.00- 6.00 |
| 变异系数 CV (%) | 10.78 | 19.82 | 15.54 | 12.43 | 17.65 | 9.12 | 18.70 | 26.31 | 22.29 | 21.93 | 10.32 | 7.84 | 1.82 | 16.00 | 48.25 |
| 山西南部South | |||||||||||||||
| 平均值±标准误 Mean±SD | 127.79± 10.86 | 29.74± 5.64 | 30.11± 4.55 | 0.82± 0.11 | 2.55± 0.44 | 13.71± 1.06 | 125.03± 23.54 | 119.92± 36.96 | 30.84± 8.57 | 23.49± 6.57 | 3.25± 0.36 | 12.93± 1.14 | 71.75± 3.41 | 0.25± 0.05 | 1.73± 0.56 |
| 范围 Range | 92.00- 151.83 | 16.67- 44.00 | 21.28- 46.00 | 0.53- 1.08 | 1.50- 3.65 | 11.00- 16.50 | 57.00- 176.50 | 54.00- 238.00 | 13.45- 55.89 | 10.17- 43.43 | 1.87- 3.98 | 10.68- 15.87 | 51.35- 75.91 | 0.17- 0.39 | 1.00- 3.50 |
| 变异系数 CV (%) | 8.50 | 18.96 | 15.12 | 13.40 | 17.06 | 7.71 | 18.83 | 30.82 | 27.77 | 27.98 | 11.17 | 8.78 | 4.75 | 19.98 | 32.15 |
新窗口打开|下载CSV
3 讨论
3.1 谷子地方品种的重新评价和利用
谷子地方品种是在自然和人工的选择中演化而来,它们保持了较高的遗传多样性。多年来,地方品种受到的关注度低于选育品种,缺乏系统深入的研究,导致利用效率较低[10]。地方品种虽然在产量相关性状上与育成品种有一定的差距,但地方品种在品质和特色育种上有很强的优势。近年来,随着谷子育种的目标向优质转变,重新评价和利用地方品种是拓展中国谷子育种遗传基础的重要途经[10]。此外,谷子种质资源的表型及农艺性状遗传多样性研究多集中在特定地区[9,10],这与谷子种质资源地区多样性高于品种多样性,且品种聚类结果与其来源地生态类型一致有关[32]。山西作为中国谷子资源最丰富的地区之一,同时包含中国谷子主产区的5个生态类型,多变的地形和气候,形成了品种类型繁多的山西谷子种质资源。本研究对212份山西谷子种质资源进行遗传多样性分析,发现除了粒色(H′<1.00)外,其他性状的多样性均高于(H′>1.00),株高的遗传多样性指数最高(2.15)(表1)。这些结果与前人对中国谷子核心种质表型多样性研究结果整体趋势一致[10-11,13],说明山西谷子地方品种的表型具有丰富的遗传变异。3.2 基于表型性状的山西谷子资源分类
聚类分析可将山西谷子种质资源分为3类(图2),即南部品种、中部品种和北部品种3大类。结合山西不同地区谷子地方品种性状的变化(表6),发现山西谷子种质资源性状由北向南成规律性变化。以千粒重为例,山西谷子种质资源性状由北向南成规律性变化。其中,特大粒品种(4.6 g)和小粒品种(2.5 g)在212份材料中占比很小,特大粒品种北部仅有1份,小粒品种南部有2份。北部材料中大粒品种占84.1%,中粒品种占13.6%;中部材料中大粒品种占60.0%,中粒品种占40.0%;南部材料中大粒品种占23.1%,中粒品种占74.4%。这与山西北部地区高寒生态区有关,因其昼夜温差大、日照时间长,有利于有机物的积累,因此,北部地区谷子大粒品种占比多。与之相反,南部地区小粒品种占比多。这一结果与张耀文等[34]对山西谷子研究得出的结论一致,谷子是喜温作物,山西地形由北向南,气温由低到高,日照由长变短,导致籽粒大小的差异,山西谷子呈现由北到南千粒重减小趋势。山西谷子地方品种表型聚类结果(图2)与山西不同地区谷子地方品种性状的变化趋势一致(表6),更能说明山西谷子地方品种表型性状的分类和地理来源吻合。北部品种单穗重、穗粒重和千粒重等产量相关性状的平均值更高,与地理环境和人工选择有关。中部品种在穗长、茎长、节数和粒色等性状上表现出更高变异性。南部品种株高和节数平均值更高,并在产量相关性状(单穗重和千粒重)以及品质性状(蛋白质、淀粉和直/支比)上表现出更高的变异性,显示出山西谷子南部品种性状多样性丰富(表6)。这一发现与王海岗等[27]对山西谷子地方品种不同地区多样性指数统计结果一致,山西同地区谷子地方品种多样性指数变化范围在1.425—1.855,运城(南部)平均多样性指数最高为1.858,南部品种可作为本生态区特异种质在谷子高产优质的新品种选育中加以利用。
3.3 山西谷子种质资源表型性状的综合评价及指标筛选
本研究利用表型性状综合得分F值进行山西谷子种质资源评价,上述方法已在水稻[19]、陆地棉[35]、谷子[11]、花生[36]等作物表型性状的综合评价中得到应用。综合评价结果显示,山西谷子的性状综合得分F值为0.521,与王海岗等[11]对来自世界各地的878份谷子核心种质的性状综合得分F值0.555相近。来自临汾的黄疙瘩F值(0.709)最高,F值排名前五的谷子品种有3个来源于山西南部品种,说明山西南部品种综合表现更好。本研究中排名靠前的山西谷子南部品种可作为本生态区特异种质在谷子新品种选育中加以利用。利用综合得分F值和表型性状构建最优回归方程,株高、穗长、茎长、码数、码粒数、单穗重、蛋白质、直/支比和粒色9个性状可作为山西谷子综合评价指标,为山西谷子地方品种的深入评价和新品种选育提供了参考依据。3.4 直/支比可作为谷子种质资源综合评价指标
谷子品质是一个综合性状,包括营养品质、商品外观、加工品质与食味品质等[37]。目前,鉴定小米食味品质已由感官评价转为理化指标分析,在优质谷子育种研究上,也开始从只关注外观品质转向理化指标与外观品质并重[38]。淀粉的结构与性质直接影响小米的食用品质和加工工艺,直链淀粉和支链淀粉的含量及比例影响着小米蒸煮品质及食味品质。研究发现,山西谷子种质资源淀粉含量变幅较小,变异程度低,变异系数仅为3.35%,是15个性状中变异系数最低的;而直/支比的变异系数高达17.47%(表1),说明直/支比可能是山西谷子蒸煮品质及食味品质表现优异的一个重要因素。相关性分析表明,表型综合得分F值与直/支比呈显著正相关(表5),说明直/支比可作为评价指标引入到谷子种质资源的综合评价中。主成分分析显示,由直/支比组成的第Ⅵ主成分对山西谷子品种的贡献率为5.86%(表4),在山西谷子地方品种资源综合评价中具有重要作用。因此,直/支比可以为山西谷子地方品种优质谷子的选育提供理论指导。4 结论
山西谷子地方品种变异幅度较大,群体多样性丰富。将山西谷子资源划分为3大类群,南部品种、中部品种和北部品种,划分结果与地理来源吻合。南部品种在产量相关性状和品质性状的变异高于北部品种,显示山西谷子南部资源多样性更丰富,能更好地作为谷子品质和特色育种的资源材料。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]

【目的】对中国谷子地方品种资源进行硒和蛋白质含量的测定与评价,以了解谷子品种资源中硒和蛋白质含量的变异及其与地理分布的关系,为谷子种质创新和育种提供基础。【方法】硒和蛋白质含量测定分别采用氢化物原子荧光法和微量凯氏定氮法。【结果】①中国谷子地方品种资源硒含量平均53.3 μg?kg-1,变异范围20.8~89.2 μg?kg-1;蛋白质含量平均15.8%,变异范围11.01%~20.77%。②不同生态区的材料硒和蛋白质含量存在明显差异,西北内陆和黄土高原来源的品种硒含量分别排列第一和第二,以下依次是东北平原、华北平原,最低是内蒙古高原;不同生态区来源的品种蛋白质含量排列次序与硒的排列较为一致,依次为西北内陆、黄土高原、华北平原、东北平原和内蒙古高原。③2个品质性状的相关分析表明,硒和蛋白质含量存在一定的正相关性,相关系数为0.534,且达到极显著水平。④硒含量与谷子粒色相关联,相联系数为0.385,达到0.05的显著水平,红粒谷子硒含量明显高于黄、青、褐等粒色的品种。【结论】谷子品种资源的硒和蛋白质含量存在较为丰富的遗传变异,在鉴定评价基础上,筛选出了一批品质优异的谷子种质资源,为富硒和高蛋白优质品种的培育和种质创新奠定了基础。
URL [本文引用: 1]
【目的】对中国谷子地方品种资源进行硒和蛋白质含量的测定与评价,以了解谷子品种资源中硒和蛋白质含量的变异及其与地理分布的关系,为谷子种质创新和育种提供基础。【方法】硒和蛋白质含量测定分别采用氢化物原子荧光法和微量凯氏定氮法。【结果】①中国谷子地方品种资源硒含量平均53.3 μg?kg-1,变异范围20.8~89.2 μg?kg-1;蛋白质含量平均15.8%,变异范围11.01%~20.77%。②不同生态区的材料硒和蛋白质含量存在明显差异,西北内陆和黄土高原来源的品种硒含量分别排列第一和第二,以下依次是东北平原、华北平原,最低是内蒙古高原;不同生态区来源的品种蛋白质含量排列次序与硒的排列较为一致,依次为西北内陆、黄土高原、华北平原、东北平原和内蒙古高原。③2个品质性状的相关分析表明,硒和蛋白质含量存在一定的正相关性,相关系数为0.534,且达到极显著水平。④硒含量与谷子粒色相关联,相联系数为0.385,达到0.05的显著水平,红粒谷子硒含量明显高于黄、青、褐等粒色的品种。【结论】谷子品种资源的硒和蛋白质含量存在较为丰富的遗传变异,在鉴定评价基础上,筛选出了一批品质优异的谷子种质资源,为富硒和高蛋白优质品种的培育和种质创新奠定了基础。
DOI:10.3724/SP.J.1006.2013.00398URL [本文引用: 1]
采用正相高效液相色谱法测定200份不同省份来源的谷子育成品种籽粒中的4种维生素E(α-VE、β-VE、γ-VE和δ-VE)及叶黄素和玉米黄素2种类胡萝卜素组分的含量,并分析了谷子维生素E含量与类胡萝卜素含量和主要农艺性状的相关性。结果显示,我国谷子育成品种籽粒中VE总量、α-VE含量及(β+γ)-VE含量均呈正态分布,δ-VE含量呈偏态分布,且(β+γ)-VE是维生素E的主要组分; 陕西和吉林谷子的VE总量和(β+γ)-VE含量显著高于其他省份品种,东北三省谷子的α-VE含量显著高于其他省份,而不同省份谷子品种中δ-VE含量差异不显著;不同省份谷子品种间叶黄素含量差异达极显著(P<0.01),玉米黄素含量差异不显著,类胡萝卜素总含量差异显著(P<0.05)。相关分析表明,参试品种籽粒中叶黄素含量与VE总量呈显著正相关,与α-VE含量呈极显著正相关,与δ-VE含量呈极显著负相关,与(β+γ)-VE含量不相关;玉米黄素与VE总量和α-VE含量呈显著正相关,与δ-VE含量呈极显著负相关,与(β+γ)-VE含量不著相关;VE总量和(β+γ)-VE含量与千粒重呈显著负相关,δ-VE含量与粒重和穗重呈极显著正相关,类胡萝卜素各组分与草重呈极显著正相关。
DOI:10.3724/SP.J.1006.2013.00398URL [本文引用: 1]
采用正相高效液相色谱法测定200份不同省份来源的谷子育成品种籽粒中的4种维生素E(α-VE、β-VE、γ-VE和δ-VE)及叶黄素和玉米黄素2种类胡萝卜素组分的含量,并分析了谷子维生素E含量与类胡萝卜素含量和主要农艺性状的相关性。结果显示,我国谷子育成品种籽粒中VE总量、α-VE含量及(β+γ)-VE含量均呈正态分布,δ-VE含量呈偏态分布,且(β+γ)-VE是维生素E的主要组分; 陕西和吉林谷子的VE总量和(β+γ)-VE含量显著高于其他省份品种,东北三省谷子的α-VE含量显著高于其他省份,而不同省份谷子品种中δ-VE含量差异不显著;不同省份谷子品种间叶黄素含量差异达极显著(P<0.01),玉米黄素含量差异不显著,类胡萝卜素总含量差异显著(P<0.05)。相关分析表明,参试品种籽粒中叶黄素含量与VE总量呈显著正相关,与α-VE含量呈极显著正相关,与δ-VE含量呈极显著负相关,与(β+γ)-VE含量不相关;玉米黄素与VE总量和α-VE含量呈显著正相关,与δ-VE含量呈极显著负相关,与(β+γ)-VE含量不著相关;VE总量和(β+γ)-VE含量与千粒重呈显著负相关,δ-VE含量与粒重和穗重呈极显著正相关,类胡萝卜素各组分与草重呈极显著正相关。
DOI:10.3864/j.issn.0578-1752.2014.07.003URL [本文引用: 1]
【目的】通过对山西谷子资源叶酸含量的测定与评价,了解谷子叶酸含量的变异及其与地理分布的关系,为谷子种质营养含量和育种提供依据。【方法】分别在谷子的研究基地长治、汾阳和太原采集目前山西育种和种植中常用品种245个,记录谷子颜色后于60℃下烘干,采用常规方法研磨脱壳去糠,记录米粒颜色后研磨米粒,全部过100目筛子,测定其叶酸含量。叶酸用磷酸二氢钾溶液恒温水浴浸提,加苯胺处理过的活性炭吸附,用3%氨—70%乙醇洗脱,采用高锰酸钾氧化—间接荧光法测定。【结果】①山西省245份不同品种谷子叶酸含量平均为1.53 μg•g-1。谷子叶酸含量数值服从正态分布且为左偏态,说明谷子叶酸含量较多集中在平均值偏高水平。②不同地区谷子叶酸含量不同。同一品种在汾阳种植其叶酸含量显著低于太原和长治。日均温、日照时数和相对湿度对叶酸含量影响不显著,降雨量则显著影响叶酸含量。③谷粒颜色对叶酸含量影响不显著,小米颜色差异显著影响叶酸含量,从高到低依次为:褐色、绿色、黄色、鲜黄、浅黄和白色米粒品种。【结论】山西省谷子资源的叶酸含量存在较为丰富的遗传变异,变异范围0.37—2.37 μg•g-1,变异系数为26.2%。不同生态区谷子叶酸含量存在明显差异,春播晚熟区的叶酸含量显著高于春播中熟区。降雨量显著影响谷子叶酸含量。小米颜色差异对叶酸含量有显著影响。在鉴定评价基础上,以样品叶酸含量的平均数及其标准差( ±s)为分类依据,筛选了一批高叶酸的谷子种质资源总计24份,占参试材料的9.8%。目前山西省农业生产常用谷子品种晋谷21,其叶酸含量约为2.0 μg•g-1,属于高叶酸含量品种。
DOI:10.3864/j.issn.0578-1752.2014.07.003URL [本文引用: 1]
【目的】通过对山西谷子资源叶酸含量的测定与评价,了解谷子叶酸含量的变异及其与地理分布的关系,为谷子种质营养含量和育种提供依据。【方法】分别在谷子的研究基地长治、汾阳和太原采集目前山西育种和种植中常用品种245个,记录谷子颜色后于60℃下烘干,采用常规方法研磨脱壳去糠,记录米粒颜色后研磨米粒,全部过100目筛子,测定其叶酸含量。叶酸用磷酸二氢钾溶液恒温水浴浸提,加苯胺处理过的活性炭吸附,用3%氨—70%乙醇洗脱,采用高锰酸钾氧化—间接荧光法测定。【结果】①山西省245份不同品种谷子叶酸含量平均为1.53 μg•g-1。谷子叶酸含量数值服从正态分布且为左偏态,说明谷子叶酸含量较多集中在平均值偏高水平。②不同地区谷子叶酸含量不同。同一品种在汾阳种植其叶酸含量显著低于太原和长治。日均温、日照时数和相对湿度对叶酸含量影响不显著,降雨量则显著影响叶酸含量。③谷粒颜色对叶酸含量影响不显著,小米颜色差异显著影响叶酸含量,从高到低依次为:褐色、绿色、黄色、鲜黄、浅黄和白色米粒品种。【结论】山西省谷子资源的叶酸含量存在较为丰富的遗传变异,变异范围0.37—2.37 μg•g-1,变异系数为26.2%。不同生态区谷子叶酸含量存在明显差异,春播晚熟区的叶酸含量显著高于春播中熟区。降雨量显著影响谷子叶酸含量。小米颜色差异对叶酸含量有显著影响。在鉴定评价基础上,以样品叶酸含量的平均数及其标准差( ±s)为分类依据,筛选了一批高叶酸的谷子种质资源总计24份,占参试材料的9.8%。目前山西省农业生产常用谷子品种晋谷21,其叶酸含量约为2.0 μg•g-1,属于高叶酸含量品种。
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/BF00132958URL [本文引用: 1]
DOI:10.1104/pp.108.129627URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 2]
对41份谷子种质资源的7个农艺性状进行了遗传多样性分析,结果表明:谷子种质资源遗传变异丰富,株高的多样性指数最高,为2.08,穗长的多样性指数最低,为1.82.基于农艺性状的聚类分析把41份材料分为4大组群,其中第一组群和第二组群分别只有一份材料,其穗粒重都很低;第三组群包括28分材料,7个农艺性状在四个组群中均是最高的;第四组群包括11分材料,7个性状值都高于第一组群和第二组群,但低于第二三组群.
.
URL [本文引用: 2]
对41份谷子种质资源的7个农艺性状进行了遗传多样性分析,结果表明:谷子种质资源遗传变异丰富,株高的多样性指数最高,为2.08,穗长的多样性指数最低,为1.82.基于农艺性状的聚类分析把41份材料分为4大组群,其中第一组群和第二组群分别只有一份材料,其穗粒重都很低;第三组群包括28分材料,7个农艺性状在四个组群中均是最高的;第四组群包括11分材料,7个性状值都高于第一组群和第二组群,但低于第二三组群.
[本文引用: 6]
[本文引用: 6]
DOI:10.3724/SP.J.1006.2016.00019URL [本文引用: 4]
通过遗传种质的多样性评估可以指导深入研究资源和育种中优异互补亲本的选择进而提高优异基因的交流累加和新品种培育的效率。本研究选用了来自世界各地的份谷子核心种质通过个表型性状的综合鉴定评估遗传多样性和筛选优异种质资源结果表明我国谷子资源的表型遗传多样性丰富单穗粒重、穗长、穗粗、株高、茎节数和生育期均表现了丰富的变异谷子育成品种遗传多样性相比农家品种下降明显育种的遗传增益主要体现在株高和穗长的适度减低,以及茎粗、茎节数、穗粗、单穗粒重、单穗重及生育期的适度增加系统聚类分析将谷子资源分成类第类以来源为东北欧国家的品种为主第类以北美和非洲的品种为主第类以东亚、南亚的品种为主我国种质可划分为春播型、春夏兼播型和南方型类型采用主成分分析法和逐步回归分析法综合评判表明叶鞘色、刚毛长度、粒色、米色、株高、穗长、茎粗和单穗粒重个性状可作为谷子表型鉴定的主要指标。
DOI:10.3724/SP.J.1006.2016.00019URL [本文引用: 4]
通过遗传种质的多样性评估可以指导深入研究资源和育种中优异互补亲本的选择进而提高优异基因的交流累加和新品种培育的效率。本研究选用了来自世界各地的份谷子核心种质通过个表型性状的综合鉴定评估遗传多样性和筛选优异种质资源结果表明我国谷子资源的表型遗传多样性丰富单穗粒重、穗长、穗粗、株高、茎节数和生育期均表现了丰富的变异谷子育成品种遗传多样性相比农家品种下降明显育种的遗传增益主要体现在株高和穗长的适度减低,以及茎粗、茎节数、穗粗、单穗粒重、单穗重及生育期的适度增加系统聚类分析将谷子资源分成类第类以来源为东北欧国家的品种为主第类以北美和非洲的品种为主第类以东亚、南亚的品种为主我国种质可划分为春播型、春夏兼播型和南方型类型采用主成分分析法和逐步回归分析法综合评判表明叶鞘色、刚毛长度、粒色、米色、株高、穗长、茎粗和单穗粒重个性状可作为谷子表型鉴定的主要指标。
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.7606/j.issn.1009-1041.2012.06.009URL [本文引用: 1]
为了解当前青海省普通小麦种质材料醇溶蛋白的遗传多样性,利用酸性聚丙烯酰胺凝胶电泳( A-PAGE )技术对青海省77份普通小麦种质材料进行了醇溶蛋白遗传多样性分析。结果表明, 供试材料中共分离出蛋白谱带1 237条,迁移率不同的谱带类型37种,其中迁移率编号为2、19和3号的谱带出现频率最高,分别为98.7%、98.7%和97.4%;3条谱带(15、16和17号)出现频率低于10.5%;其余31条谱带出现频率为19.5%~84.4%。供试材料醇溶蛋白谱带多态性较高,每个材料可电泳分离出11~21条谱带,其中具有14~18条谱带的材料居多。不同材料间的遗传相似系数变化范围为0. 55~0.94,说明供试材料具有丰富的遗传多样性。聚类分析将供试材料分成6大类,聚类结果在一定程度上反映了供试材料间的亲缘关系。
DOI:10.7606/j.issn.1009-1041.2012.06.009URL [本文引用: 1]
为了解当前青海省普通小麦种质材料醇溶蛋白的遗传多样性,利用酸性聚丙烯酰胺凝胶电泳( A-PAGE )技术对青海省77份普通小麦种质材料进行了醇溶蛋白遗传多样性分析。结果表明, 供试材料中共分离出蛋白谱带1 237条,迁移率不同的谱带类型37种,其中迁移率编号为2、19和3号的谱带出现频率最高,分别为98.7%、98.7%和97.4%;3条谱带(15、16和17号)出现频率低于10.5%;其余31条谱带出现频率为19.5%~84.4%。供试材料醇溶蛋白谱带多态性较高,每个材料可电泳分离出11~21条谱带,其中具有14~18条谱带的材料居多。不同材料间的遗传相似系数变化范围为0. 55~0.94,说明供试材料具有丰富的遗传多样性。聚类分析将供试材料分成6大类,聚类结果在一定程度上反映了供试材料间的亲缘关系。
[本文引用: 1]
[本文引用: 1]
DOI:10.3724/SP.J.1006.2012.00829URL [本文引用: 2]
种质资源是水稻遗传育种和解析复杂性状的重要基础,而对遗传多样评估有利于鉴定最优亲本组合以产生遗传变异最大的后代群体和促进不同资源的有利基因渗透到栽培品种。选用14个表型性状对美国农业部水稻核心种质中的6大洲1 579份水稻种质, 分析与评价其遗传多样性和优良稻种资源,主要结果如下: (1)亚洲、非洲与大洋洲间遗传距离较远,且亚洲、非洲和大洋洲的水稻资源具有较丰富的表型遗传多样性,而不同性状的遗传多样性在洲际间表现不同;粒长宽比、碱消值、株高、粒宽、千粒重和淀粉含量等6个性状具有很高的表型遗传多样性。(2)采用主成分分析法和逐步回归分析法综合评判表明,越南的PI392768的综合性状表现最好,法国的PI281760综合性状表现最差,同时淀粉含量、抽穗期、株高、倒伏性、糙米色和颖壳色等6个性状可作为种质资源综合评价指标。在水稻育种中应注重利用具有丰富遗传多样性的种质资源,并在亲本选配时适当选择遗传距离较远且综合性状表现差异大的种质材料。
DOI:10.3724/SP.J.1006.2012.00829URL [本文引用: 2]
种质资源是水稻遗传育种和解析复杂性状的重要基础,而对遗传多样评估有利于鉴定最优亲本组合以产生遗传变异最大的后代群体和促进不同资源的有利基因渗透到栽培品种。选用14个表型性状对美国农业部水稻核心种质中的6大洲1 579份水稻种质, 分析与评价其遗传多样性和优良稻种资源,主要结果如下: (1)亚洲、非洲与大洋洲间遗传距离较远,且亚洲、非洲和大洋洲的水稻资源具有较丰富的表型遗传多样性,而不同性状的遗传多样性在洲际间表现不同;粒长宽比、碱消值、株高、粒宽、千粒重和淀粉含量等6个性状具有很高的表型遗传多样性。(2)采用主成分分析法和逐步回归分析法综合评判表明,越南的PI392768的综合性状表现最好,法国的PI281760综合性状表现最差,同时淀粉含量、抽穗期、株高、倒伏性、糙米色和颖壳色等6个性状可作为种质资源综合评价指标。在水稻育种中应注重利用具有丰富遗传多样性的种质资源,并在亲本选配时适当选择遗传距离较远且综合性状表现差异大的种质材料。
[本文引用: 1]
[本文引用: 1]
DOI:10.3321/j.issn:1000-7091.1999.03.006URL [本文引用: 1]
对山西省3039份谷子品种资源以米质、穗型、千粒重、生育期、幼苗叶色、幼苗叶鞘色、粒色、米色进行了分类。并对不同类型品种在山西省大同地区、忻州地区、晋中地区、吕梁地区、长治地区、晋南地区的分布进行了统计分析。归纳了不同类型品种与生态环境相互关系的规律性,从而更好地利用这些规律性,为谷子高产栽培、异地种植提供科学依据。
DOI:10.3321/j.issn:1000-7091.1999.03.006URL [本文引用: 1]
对山西省3039份谷子品种资源以米质、穗型、千粒重、生育期、幼苗叶色、幼苗叶鞘色、粒色、米色进行了分类。并对不同类型品种在山西省大同地区、忻州地区、晋中地区、吕梁地区、长治地区、晋南地区的分布进行了统计分析。归纳了不同类型品种与生态环境相互关系的规律性,从而更好地利用这些规律性,为谷子高产栽培、异地种植提供科学依据。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.3864/j.issn.0578-1752.2019.22.013URL [本文引用: 1]
【目的】分析山西谷子地方品种遗传多样性和群体遗传结构,筛选与谷子农艺性状相关联的分子标记,为谷子杂交组合亲本选配及分子标记辅助育种提供依据。【方法】 利用96对SSR标记对595份山西谷子核心资源进行全基因组扫描,采用PowerMarker 3.25软件分析群体遗传多样性,利用STRUCTURE 2.3.4软件分析群体遗传结构,使用TASSEL 2.1软件中GLM(general linear model,Q)和MLM(mixed linear model,Q+K)2种方法,进行表型和标记关联分析。【结果】 96对SSR引物共扩增出828个等位变异,平均每对引物扩增到8.6个,变化范围为2—26;基因多样性指数变化范围为0.005—0.941,平均为0.610;多态信息量变化范围为0.005—0.938,平均为0.577;各位点杂合度变化范围为0—0.050,平均位点杂合度仅为0.016。群体结构分析将595份核心资源分为3个亚群。4 560个SSR位点成对组合中,共线性组合和非共线性组合之间都存在一定的连锁不平衡。D′统计概率(P<0.01)支持的LD成对位点1 955个,占全部位点组合的42.9%,D′平均值为0.23。通过GLM方法共检测到12个极显著性位点(P<0.01),表型变异解释率为2.34%—13.94%,平均为6.33%,贡献率较高的等位变异位点是CAAS2050(R 2=13.94%)和B153(R 2=11.36%);通过MLM方法共检测到9个极显著性位点(P<0.01),表型变异解释率为2.80%—9.22%,平均为5.16%,贡献率较高的等位变异位点是P89(R 2=9.22%)和P3*(R 2=8.28%);2种方法共同检测到的极显著性位点有7个。 【结论】 利用SSR标记分析了595份山西谷子核心资源的遗传多样性和群体遗传结构。2种关联分析模型中,GLM方法关联到12个标记与节数、株高、颈长、茎粗、穗长、穗粗、码数、码粒数、蛋白质含量9个性状相关;MLM方法关联到9个标记与节数、颈长、叶宽、茎粗、穗粗、码数、码粒数、千粒重8个性状相关。
DOI:10.3864/j.issn.0578-1752.2019.22.013URL [本文引用: 1]
【目的】分析山西谷子地方品种遗传多样性和群体遗传结构,筛选与谷子农艺性状相关联的分子标记,为谷子杂交组合亲本选配及分子标记辅助育种提供依据。【方法】 利用96对SSR标记对595份山西谷子核心资源进行全基因组扫描,采用PowerMarker 3.25软件分析群体遗传多样性,利用STRUCTURE 2.3.4软件分析群体遗传结构,使用TASSEL 2.1软件中GLM(general linear model,Q)和MLM(mixed linear model,Q+K)2种方法,进行表型和标记关联分析。【结果】 96对SSR引物共扩增出828个等位变异,平均每对引物扩增到8.6个,变化范围为2—26;基因多样性指数变化范围为0.005—0.941,平均为0.610;多态信息量变化范围为0.005—0.938,平均为0.577;各位点杂合度变化范围为0—0.050,平均位点杂合度仅为0.016。群体结构分析将595份核心资源分为3个亚群。4 560个SSR位点成对组合中,共线性组合和非共线性组合之间都存在一定的连锁不平衡。D′统计概率(P<0.01)支持的LD成对位点1 955个,占全部位点组合的42.9%,D′平均值为0.23。通过GLM方法共检测到12个极显著性位点(P<0.01),表型变异解释率为2.34%—13.94%,平均为6.33%,贡献率较高的等位变异位点是CAAS2050(R 2=13.94%)和B153(R 2=11.36%);通过MLM方法共检测到9个极显著性位点(P<0.01),表型变异解释率为2.80%—9.22%,平均为5.16%,贡献率较高的等位变异位点是P89(R 2=9.22%)和P3*(R 2=8.28%);2种方法共同检测到的极显著性位点有7个。 【结论】 利用SSR标记分析了595份山西谷子核心资源的遗传多样性和群体遗传结构。2种关联分析模型中,GLM方法关联到12个标记与节数、株高、颈长、茎粗、穗长、穗粗、码数、码粒数、蛋白质含量9个性状相关;MLM方法关联到9个标记与节数、颈长、叶宽、茎粗、穗粗、码数、码粒数、千粒重8个性状相关。
DOI:10.3864/j.issn.0578-1752.2016.17.001URL [本文引用: 1]
谷子(Setaria italica Beauv.)和黍稷(Panicum miliaceum L.)是中国起源的古老农作物,栽培历史超过8 000年,粟(谷子)、黍(黍稷、糜子)、稻(水稻Oryza sativa L.)、麦(小麦Triticum aestivum L.)、菽(大豆Glycine max(Linn.) Merr.)被称为中国传统农耕文明的“五谷”[1-2],而谷子和黍稷在“五谷”中位列重要位置,可见这两种作物对中华文明发展的重要性。考古学证据的积累证明,黍稷的驯化早于谷子,在8 000以前的新石器时代黍稷就已经在黄河流域广泛栽培;而谷子的驯化略晚于黍稷,在距今6 000— 7 000年前被广泛栽培。已有证据表明,这两个作物均起源于中国北方的黄河流域,在水稻尚未传到北方,小麦尚未引入中国的农耕文化形成早期,对中华文明的形成起到了决定性作用,因此,谷子和黍稷被誉为中华民族的哺育作物[3-4],数千年来它们一直是中国北方旱作生态农业的主栽作物。随着小麦的引入特别是玉米(Zea mays L.)和甘薯(Dioscorea esculenta (Lour.) Burkill)引入中国后,谷子和黍稷的栽培面积逐渐减小,但仍然在旱作农业中占有重要地位,是干旱半干旱地区的主要作物。谷子黍稷均抗旱性突出,早在1927年,美国****Shantz等[5]研究证明谷子和黍稷是禾谷类作物中水分利用效率最高的环境友好作物。随着世界范围的水资源减少和生态环境日益恶化,以及对水肥高消耗主栽作物所存在的环境恶化问题认识的加深,大家重新认识到谷子和黍稷在环境友好型可持续农业中的重要性,特别是谷子和其近缘野生种青狗尾草(Setaria viridis),因其小的二倍体基因组和易于实验室操作的株型,正在迅速发展成为世界广泛关注的抗旱耐逆研究和C4光合作用研究的模式作物[6-7]。 由于谷子和黍稷只是北方旱作区的区域经济重要作物,研究投入和团队力量有限,这两个作物的基础研究同主要农作物水稻、玉米、小麦相差很远,在国家级刊物上发表的正规论文一年也没有几篇,基础研究极为薄弱。在国家产业技术体系、国家科技支撑计划和国家自然科学基金的资助下,谷子和黍稷的基础研究有了显著提升。谷子野生祖先青狗尾草、农家品种和育成品种的群体结构得到了解析[8-11],促进了谷子和黍稷资源研究和育种水平的提升。《中国农业科学》本期以谷子黍稷专题形式发表谷子和黍稷基础研究的5篇文章,报道了从品种资源多样性、抗旱耐逆、营养高效和主要病害抗性资源鉴定等方面的工作进展,说明中国谷子和黍稷的基础研究近年来已有较大的起色。
中国有着丰富的黍稷资源,虽然已构建了黍稷的核心种质[12],并且也开展了一些资源的多样性研究和分类,但主要是依据农艺表型性状[13],覆盖较广范围资源基于基因组变异的遗传多样性分析尚未见报道。本期专题发表的连帅等作者利用SSR标记对黍稷资源多样性的研究,分析了来自国内外的192份黍稷地方品种和野生种质,资源的地理来源广泛;所用SSR标记覆盖整个基因组,较全面地解析了黍稷基因组变异的本底,基本清楚了10个地理群黍稷资源的遗传关系。该工作对于黍稷资源管理、育种和遗传研究杂交组合配置,以及黍稷的起源分析有重要参考价值。黑穗病是黍稷的最主要病害,本期发表的周瑜等作者的工作,不仅为读者提供了黍稷黑穗病抗性鉴定的新结果,更给出了抗病品种和感病品种在接种病菌后的苯丙氨酸解氨酶(phenylalanine ammonia lyase,PAL)等活性变化,得出了拔节期和灌浆期PAL活性、抽穗期和灌浆期抗坏血酸过氧化物酶(ascorbate peroxidase,APX)活性、抽穗期谷胱甘肽还原酶(glutathione reductase,GR)活性、灌浆期的还原型谷胱甘肽(reduced glutathione,GSH)等在抗病品种和感病品种间存在显著差异,可作为鉴定黍稷对黑穗病抗性的生理生化指标的结果,这对黍稷抗病育种和资源鉴定有重要参考意义,是对黍稷资源抗黑穗病鉴定[14]研究的发展和深入。
谷子是公认的抗旱耐逆营养高效作物,传统的谷子抗旱耐逆研究主要是资源的抗旱耐盐鉴定[15-16],虽然近年来发表了几篇抗旱相关QTL发掘和表达分析的文章[17-18],但从单基因水平分析的研究还很少。本期谷子糜子专题发表了秦玉海等在谷子耐逆相关基因SibZIP42转录因子的功能分析研究结果,证明 SibZIP42转基因拟南芥株系相对于野生型在种子萌发时期耐盐性显著提高,在种子萌发后期SibZIP42转基因株系相比于野生型对ABA处理的敏感性增强,说明SibZIP42可能是通过ABA信号途径正向调控植物的耐盐性。《中国农业科学》本期发表的陈二影等利用79个夏谷品种对苗期氮素利用效率分析,发现不同谷子品种苗期氮效率差异显著,且西北春谷类型品种间氮素吸收效率差异最大,华北夏谷类型品种间氮素利用效率差异最大,为深入研究谷子的氮素营养机理和克隆氮素营养高效相关基因提供了基础,也是在谷子营养高效资源鉴定方面的首篇文章。谷瘟病是谷子的主要病害,而且近年来有愈发严重的趋势[19],本期谷子黍稷专题发表的李志江等作者在谷瘟病方面的工作不仅鉴定出了多个高抗谷瘟病的抗源品种,而且构建了谷瘟病菌生理小种鉴别的谷子标准品种体系,对今后的谷瘟病小种鉴定和谷子不同产区小种类型分析和抗病育种与栽培有指导意义。
作为中国最早驯化的作物,谷子和黍稷是中国传统农耕文化的重要载体作物。谷子和黍稷的抗旱耐逆性强,合成干物质需水少[5],被称为应对未来日益干旱环境的战略储备作物和环境友好作物[20]。近年来,谷子和野生祖先青狗尾草正在迅速发展成为禾谷类耐逆和C4光合作用研究新的模式作物[6],这些都增加了谷子和黍稷的世界关注度。中国是谷子和黍稷的遗传多样性中心,拥有最丰富的遗传资源[2],也是世界上谷子和黍稷栽培面积最大的国家,中国的谷子和黍稷基础研究理应走在国际前列,为国家粮食安全和农作物及膳食多样性发展作出贡献。谷子方面中国的基础研究进展已获得国际高度关注,并于2014年在北京召开了首届国际谷子遗传学会议[6]。《中国农业科学》本期谷子黍稷专题发表的5篇文章,必将进一步促进谷子和黍稷的基础研究工作,提升中国在相关领域的国际竞争力。
DOI:10.3864/j.issn.0578-1752.2016.17.001URL [本文引用: 1]
谷子(Setaria italica Beauv.)和黍稷(Panicum miliaceum L.)是中国起源的古老农作物,栽培历史超过8 000年,粟(谷子)、黍(黍稷、糜子)、稻(水稻Oryza sativa L.)、麦(小麦Triticum aestivum L.)、菽(大豆Glycine max(Linn.) Merr.)被称为中国传统农耕文明的“五谷”[1-2],而谷子和黍稷在“五谷”中位列重要位置,可见这两种作物对中华文明发展的重要性。考古学证据的积累证明,黍稷的驯化早于谷子,在8 000以前的新石器时代黍稷就已经在黄河流域广泛栽培;而谷子的驯化略晚于黍稷,在距今6 000— 7 000年前被广泛栽培。已有证据表明,这两个作物均起源于中国北方的黄河流域,在水稻尚未传到北方,小麦尚未引入中国的农耕文化形成早期,对中华文明的形成起到了决定性作用,因此,谷子和黍稷被誉为中华民族的哺育作物[3-4],数千年来它们一直是中国北方旱作生态农业的主栽作物。随着小麦的引入特别是玉米(Zea mays L.)和甘薯(Dioscorea esculenta (Lour.) Burkill)引入中国后,谷子和黍稷的栽培面积逐渐减小,但仍然在旱作农业中占有重要地位,是干旱半干旱地区的主要作物。谷子黍稷均抗旱性突出,早在1927年,美国****Shantz等[5]研究证明谷子和黍稷是禾谷类作物中水分利用效率最高的环境友好作物。随着世界范围的水资源减少和生态环境日益恶化,以及对水肥高消耗主栽作物所存在的环境恶化问题认识的加深,大家重新认识到谷子和黍稷在环境友好型可持续农业中的重要性,特别是谷子和其近缘野生种青狗尾草(Setaria viridis),因其小的二倍体基因组和易于实验室操作的株型,正在迅速发展成为世界广泛关注的抗旱耐逆研究和C4光合作用研究的模式作物[6-7]。 由于谷子和黍稷只是北方旱作区的区域经济重要作物,研究投入和团队力量有限,这两个作物的基础研究同主要农作物水稻、玉米、小麦相差很远,在国家级刊物上发表的正规论文一年也没有几篇,基础研究极为薄弱。在国家产业技术体系、国家科技支撑计划和国家自然科学基金的资助下,谷子和黍稷的基础研究有了显著提升。谷子野生祖先青狗尾草、农家品种和育成品种的群体结构得到了解析[8-11],促进了谷子和黍稷资源研究和育种水平的提升。《中国农业科学》本期以谷子黍稷专题形式发表谷子和黍稷基础研究的5篇文章,报道了从品种资源多样性、抗旱耐逆、营养高效和主要病害抗性资源鉴定等方面的工作进展,说明中国谷子和黍稷的基础研究近年来已有较大的起色。
中国有着丰富的黍稷资源,虽然已构建了黍稷的核心种质[12],并且也开展了一些资源的多样性研究和分类,但主要是依据农艺表型性状[13],覆盖较广范围资源基于基因组变异的遗传多样性分析尚未见报道。本期专题发表的连帅等作者利用SSR标记对黍稷资源多样性的研究,分析了来自国内外的192份黍稷地方品种和野生种质,资源的地理来源广泛;所用SSR标记覆盖整个基因组,较全面地解析了黍稷基因组变异的本底,基本清楚了10个地理群黍稷资源的遗传关系。该工作对于黍稷资源管理、育种和遗传研究杂交组合配置,以及黍稷的起源分析有重要参考价值。黑穗病是黍稷的最主要病害,本期发表的周瑜等作者的工作,不仅为读者提供了黍稷黑穗病抗性鉴定的新结果,更给出了抗病品种和感病品种在接种病菌后的苯丙氨酸解氨酶(phenylalanine ammonia lyase,PAL)等活性变化,得出了拔节期和灌浆期PAL活性、抽穗期和灌浆期抗坏血酸过氧化物酶(ascorbate peroxidase,APX)活性、抽穗期谷胱甘肽还原酶(glutathione reductase,GR)活性、灌浆期的还原型谷胱甘肽(reduced glutathione,GSH)等在抗病品种和感病品种间存在显著差异,可作为鉴定黍稷对黑穗病抗性的生理生化指标的结果,这对黍稷抗病育种和资源鉴定有重要参考意义,是对黍稷资源抗黑穗病鉴定[14]研究的发展和深入。
谷子是公认的抗旱耐逆营养高效作物,传统的谷子抗旱耐逆研究主要是资源的抗旱耐盐鉴定[15-16],虽然近年来发表了几篇抗旱相关QTL发掘和表达分析的文章[17-18],但从单基因水平分析的研究还很少。本期谷子糜子专题发表了秦玉海等在谷子耐逆相关基因SibZIP42转录因子的功能分析研究结果,证明 SibZIP42转基因拟南芥株系相对于野生型在种子萌发时期耐盐性显著提高,在种子萌发后期SibZIP42转基因株系相比于野生型对ABA处理的敏感性增强,说明SibZIP42可能是通过ABA信号途径正向调控植物的耐盐性。《中国农业科学》本期发表的陈二影等利用79个夏谷品种对苗期氮素利用效率分析,发现不同谷子品种苗期氮效率差异显著,且西北春谷类型品种间氮素吸收效率差异最大,华北夏谷类型品种间氮素利用效率差异最大,为深入研究谷子的氮素营养机理和克隆氮素营养高效相关基因提供了基础,也是在谷子营养高效资源鉴定方面的首篇文章。谷瘟病是谷子的主要病害,而且近年来有愈发严重的趋势[19],本期谷子黍稷专题发表的李志江等作者在谷瘟病方面的工作不仅鉴定出了多个高抗谷瘟病的抗源品种,而且构建了谷瘟病菌生理小种鉴别的谷子标准品种体系,对今后的谷瘟病小种鉴定和谷子不同产区小种类型分析和抗病育种与栽培有指导意义。
作为中国最早驯化的作物,谷子和黍稷是中国传统农耕文化的重要载体作物。谷子和黍稷的抗旱耐逆性强,合成干物质需水少[5],被称为应对未来日益干旱环境的战略储备作物和环境友好作物[20]。近年来,谷子和野生祖先青狗尾草正在迅速发展成为禾谷类耐逆和C4光合作用研究新的模式作物[6],这些都增加了谷子和黍稷的世界关注度。中国是谷子和黍稷的遗传多样性中心,拥有最丰富的遗传资源[2],也是世界上谷子和黍稷栽培面积最大的国家,中国的谷子和黍稷基础研究理应走在国际前列,为国家粮食安全和农作物及膳食多样性发展作出贡献。谷子方面中国的基础研究进展已获得国际高度关注,并于2014年在北京召开了首届国际谷子遗传学会议[6]。《中国农业科学》本期谷子黍稷专题发表的5篇文章,必将进一步促进谷子和黍稷的基础研究工作,提升中国在相关领域的国际竞争力。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
.
[本文引用: 1]
DOI:10.3321/j.issn:1000-7091.1992.04.022URL
以160个代表点的14个与谷子适应性有关的生态因子综合效应因子荷载值、全国谷子生态联合试验所确定的谷子品种光温生态类型以及谷子品种全国统一熟期分类三者结合划区。将中国谷子主产区(占全国谷子播种面积的97%)划分为春播特早熟区、早熟区,中熟区、晚熟区和夏谷区等5个生态区11个亚区。分区分析了生态特点和品种特性,指出存在的问题。本区划具有以下特点:(1)以气候地理因子与谷子生态特性,特别是按谷子统一熟期分类结合划区,使相同生态条件地区连片,突破了大行政区的框架;(2)代表地点和因子多,且又通过运算综合成因子荷载值划区,代表性大;(3)解决了过去划区中承德、朝阳、赤峰、铁岭、锦州等重要产谷区归属模糊的问题。
DOI:10.3321/j.issn:1000-7091.1992.04.022URL
以160个代表点的14个与谷子适应性有关的生态因子综合效应因子荷载值、全国谷子生态联合试验所确定的谷子品种光温生态类型以及谷子品种全国统一熟期分类三者结合划区。将中国谷子主产区(占全国谷子播种面积的97%)划分为春播特早熟区、早熟区,中熟区、晚熟区和夏谷区等5个生态区11个亚区。分区分析了生态特点和品种特性,指出存在的问题。本区划具有以下特点:(1)以气候地理因子与谷子生态特性,特别是按谷子统一熟期分类结合划区,使相同生态条件地区连片,突破了大行政区的框架;(2)代表地点和因子多,且又通过运算综合成因子荷载值划区,代表性大;(3)解决了过去划区中承德、朝阳、赤峰、铁岭、锦州等重要产谷区归属模糊的问题。
[本文引用: 1]
[本文引用: 1]
DOI:10.3864/j.issn.0578-1752.2016.19.003URL [本文引用: 1]
【目的】分析陆地棉核心种质的遗传多样性和表型性状遗传变异规律,并探讨核心种质的综合评价方法。【方法】利用17个表型性状数据分析419份陆地棉核心种质的遗传多样性。用Shannon-weaver信息多样性指数计算表型性状的遗传多样性,用Nei’s 1973法计算表型性状遗传距离,并使用NTSYS-pc 2.20q软件对核心种质进行聚类分析;用SAS9.2对表型性状数据进行最佳线性无偏估计(BLUE),然后根据最佳线性无偏估计值计算出表型性状的最佳值。同时,结合主成分、回归和相关分析,研究核心种质的综合评价指标和方法。【结果】核心种质表型性状分析发现,单株铃数、单铃重、衣分、子指等性状的变异系数均较大,变异系数超过10%。而断裂比强度、马克隆值以及上半部平均长度的变异程度较小,变异系数均在10%以下。方差分析发现,各表型性状地点间、年份间、地点和年份间、品种间均有极显著差异;不同地理来源的种质表型性状差异较大,长江流域地理来源的种质生育期、伸长率、上半部平均长度、衣分等性状均高于其他的地理来源,西北内陆地理来源的种质纤维强度,单铃重、整齐度指数、株高、纺纱均匀性指数等综合性状最好,美国种质的产量和纤维品质的性状优于其他国家的总和。表型性状的遗传多样性指数范围为0.351—3.796,平均为1.715。分析不同地理来源种质的遗传多样性,发现黄河流域的遗传多样性和遗传丰富度最高,中国南部区域最低。类群聚类结果发现陆地棉整体分散,没有比较明显的类群关系,部分具有相似特点的种质聚类13个组群。核心种质综合评价表明在累计贡献百分比高于85%时,共发现7个主成分,陆地棉核心种质的表型性状综合值(F值)平均为1.740,来自澳大利亚的N74-250F值最高(2.302),辽阳绿绒棉的F值最低(0.624)。对17个表型性状与F值的相关分析,发现除马克隆值、子指和黄度外,单铃重、衣分、断裂比强度、上半部纤维长度等14个表型性状与F值间的相关性具有极显著差异,最后构建了以吐絮期、单铃重、伸长率、花期、马克隆值、株高、果枝数、纺纱均匀性指数8个表型性状为自变量的回归方程,综合评价核心种质资源。【结论】中国保存的陆地棉核心种质具有较为丰富的遗传多样性,不同地理来源遗传变异有较大的差异,不同生态区的核心种质具有独特的性状特性。
DOI:10.3864/j.issn.0578-1752.2016.19.003URL [本文引用: 1]
【目的】分析陆地棉核心种质的遗传多样性和表型性状遗传变异规律,并探讨核心种质的综合评价方法。【方法】利用17个表型性状数据分析419份陆地棉核心种质的遗传多样性。用Shannon-weaver信息多样性指数计算表型性状的遗传多样性,用Nei’s 1973法计算表型性状遗传距离,并使用NTSYS-pc 2.20q软件对核心种质进行聚类分析;用SAS9.2对表型性状数据进行最佳线性无偏估计(BLUE),然后根据最佳线性无偏估计值计算出表型性状的最佳值。同时,结合主成分、回归和相关分析,研究核心种质的综合评价指标和方法。【结果】核心种质表型性状分析发现,单株铃数、单铃重、衣分、子指等性状的变异系数均较大,变异系数超过10%。而断裂比强度、马克隆值以及上半部平均长度的变异程度较小,变异系数均在10%以下。方差分析发现,各表型性状地点间、年份间、地点和年份间、品种间均有极显著差异;不同地理来源的种质表型性状差异较大,长江流域地理来源的种质生育期、伸长率、上半部平均长度、衣分等性状均高于其他的地理来源,西北内陆地理来源的种质纤维强度,单铃重、整齐度指数、株高、纺纱均匀性指数等综合性状最好,美国种质的产量和纤维品质的性状优于其他国家的总和。表型性状的遗传多样性指数范围为0.351—3.796,平均为1.715。分析不同地理来源种质的遗传多样性,发现黄河流域的遗传多样性和遗传丰富度最高,中国南部区域最低。类群聚类结果发现陆地棉整体分散,没有比较明显的类群关系,部分具有相似特点的种质聚类13个组群。核心种质综合评价表明在累计贡献百分比高于85%时,共发现7个主成分,陆地棉核心种质的表型性状综合值(F值)平均为1.740,来自澳大利亚的N74-250F值最高(2.302),辽阳绿绒棉的F值最低(0.624)。对17个表型性状与F值的相关分析,发现除马克隆值、子指和黄度外,单铃重、衣分、断裂比强度、上半部纤维长度等14个表型性状与F值间的相关性具有极显著差异,最后构建了以吐絮期、单铃重、伸长率、花期、马克隆值、株高、果枝数、纺纱均匀性指数8个表型性状为自变量的回归方程,综合评价核心种质资源。【结论】中国保存的陆地棉核心种质具有较为丰富的遗传多样性,不同地理来源遗传变异有较大的差异,不同生态区的核心种质具有独特的性状特性。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}