Splicing Property Analyses of the NRSE1 Element from Tobacco PR3b mRNA After Fusion Expression with GUS Gene
ZHAO Xue1, WANG Feng2, WANG WenJing1, LIU XiaoFeng1, BIAN ShiQuan1, LIU YanHua1, LIU XinMin1, DU YongMei1, ZHANG ZhongFeng1, ZHANG HongBo,11 Tobacco Research Institute, Chinese Academy of Agricultural Sciences, Qingdao 266101, Shandong 2 College of Agronomy and Biotechnology, Southwest University, Chongqing 400716
Received:2019-09-5Accepted:2019-12-18Online:2020-04-16 作者简介 About authors 赵雪,E-mail: zhaoxue1007@163.com。
摘要 【目的】烟草(Nicotiana tabacum L.)碱性几丁质酶基因PR3b在低烟碱突变体(nic1和nic2)中存在转录后mRNA可变剪切现象,但其可变剪切的发生机制仍不清楚。将PR3b的可变剪切元件NRSE1(nicotine- synthesis related splicing element 1)与GUS融合表达,分析NRSE1元件的独立可变剪切特性,以揭示其作用机制。【方法】利用PCR扩增方法获得PR3b cDNA序列中的NRSE1元件片段,并利用基因重组技术构建了烟草PR3b可变剪切元件NRSE1与GUS的融合表达载体。将融合表达载体导入农杆菌LBA4404后,通过农杆菌介导的叶盘转化法培育了表达NRSE1与GUS融合子的低烟碱突变体nic1和nic2及野生型烟草转基因植株;通过RT-PCR检测及GUS染色鉴定出阳性植株后,利用RT-PCR分析NRSE1与GUS融合表达后在低烟碱突变体和野生型烟草中的可变剪切特性;对转基因植株的幼苗进行乙烯(ET)和茉莉酸(JA)处理,通过GUS染色方法分析ET和JA处理对转基因植株中GUS活性的影响,并通过RT-PCR方法分析ET和JA处理对转基因植株中NRSE1与GUS融合子的可变剪切特性影响,以及对转基因植株中NRSE1与GUS融合子表达水平的影响。【结果】通过RT-PCR检测及GUS染色鉴定出表达NRSE1元件与GUS融合子的低烟碱突变体和野生型烟草转基因植株;RT-PCR检测及测序分析证明,NRSE1元件与GUS融合表达后仍能在低烟碱突变体发生高水平的可变剪切,剪切修饰区段的序列变化与烟草中PR3b的mRNA可变剪切修饰一致;利用ET和JA处理转基因植株进行的GUS染色表明,ET和JA处理对转基因植株的GUS活性有不同程度的影响;但利用ET和JA处理转基因植株进行的RT-PCR分析表明,ET和JA处理不改变NRSE1元件原有的诱导剪切特性,也不影响转基因植株中NRSE1元件与GUS融合子的表达水平。【结论】PR3b的可变剪切元件NRSE1与GUS在烟草中融合表达后,仍能在低烟碱突变体nic1和nic2中发生高水平的可变剪切;NRSE1在烟草中的可变剪切不依赖PR3b的其他mRNA区段,是烟草PR3b发生可变剪切的独立元件;ET和JA处理对NRSE1元件与GUS融合表达植株的GUS活性具有一定影响,可能存在翻译水平的调控作用。 关键词:烟草;PR3b;低烟碱突变体;可变剪切;茉莉酸;乙烯
Abstract 【Objective】Previously, a post-transcriptional splicing of tobacco PR3b gene was observed in the low-nicotine mutants (nic1, nic2) of Nicotiana tabacum L. cv. Burley 21, yet the mechanism underlying this phenomenon is still unclear. In this study, we developed transgenic plants expressing the fusion of the alternative splicing element NRSE1 (nicotine-synthesis related splicing element 1) from PR3b and the GUS gene to investigate the splicing properties of the NRSE1 element after excising from PR3b mRNA, in order to reveal its regulatory mechanism. 【Method】The NRSE1 element was amplified from PR3b cDNA by PCR amplification, and the vector for expressing the fusion of NRSE1 element and the GUS gene was constructed by molecular methods. And, the vector was used to develop transgenic plants expressing the fusion of NRSE1 element and GUS gene with wild type tobacco and the low-nicotine mutants nic1 and nic2 via agrobacterium (LBA4404) mediated transformation method. The transgenic plants were identified by RT-PCR and GUS staining, and the splicing of the fusion of NRSE1 element and GUS gene in the transgenic plants of wild type tobacco and low-nicotine mutants were then analyzed by RT-PCR. Seedlings of the transgenic plants were treated with ethylene (ET) and jasmonic acid (JA), respectively. And, the effects of ET and JA treatment on the GUS activity and the splicing of the fusion of NRSE1 element and GUS gene in the transgenic plants were analyzed by GUS staining and RT-PCR, respectively. The effects of ET and JA treatment on the expression level of the fusion of NRSE1 element and GUS gene were analyzed as well. 【Result】A set of transgenic wild type tobacco and low-nicotine mutants expressing the fusion of NRSE1 element and GUS gene were identified by RT-PCR and GUS staining. Further RT-PCR and sequencing analyses showed that the NRSE1 element could be alternatively spliced at higher levels in the low-nicotine mutants after fusion with the GUS gene, in a pattern consistent with its alternative splicing in the PR3b mRNA as previously report. ET and JA treatments could alter the GUS activity in the transgenic plants, but did not affect the inducible splicing of the NRSE1 element or the expression level of the fusion of NRSE1 element and GUS gene in the transgenic plants. 【Conclusion】 A highly splicing of the NRSE1 element was observed in the low-nicotine mutants after fusion expression with GUS gene. The alternatively splicing of NRSE1 element is independent of the rest regions of PR3b mRNA. ET and JA treatments had an effect on the GUS activities of the transgenic plants expressing the fusion of NRSE1 element and GUS gene, which may result from a translational regulation. Keywords:Nicotiana tabacum L.;PR3b gene;low-nicotine mutant;alternative splicing;jasmonic acid;ethylene
PDF (1403KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 赵雪, 王锋, 王文静, 刘晓峰, 卞士权, 刘艳华, 刘新民, 杜咏梅, 张忠锋, 张洪博. 烟草PR3b转录后剪切元件NRSE1与GUS融合表达后的可变剪切[J]. 中国农业科学, 2020, 53(8): 1524-1531 doi:10.3864/j.issn.0578-1752.2020.08.003 ZHAO Xue, WANG Feng, WANG WenJing, LIU XiaoFeng, BIAN ShiQuan, LIU YanHua, LIU XinMin, DU YongMei, ZHANG ZhongFeng, ZHANG HongBo. Splicing Property Analyses of the NRSE1 Element from Tobacco PR3b mRNA After Fusion Expression with GUS Gene[J]. Scientia Acricultura Sinica, 2020, 53(8): 1524-1531 doi:10.3864/j.issn.0578-1752.2020.08.003
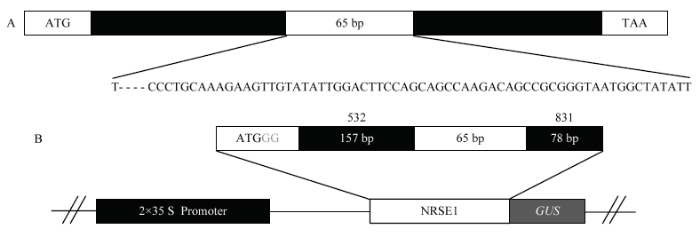
A:PR3b示意图以及可变剪切区65 bp碱基序列,剪切的第1个与后64个碱基间有4个碱基间隔;B:NRSE1元件与GUS的融合表达载体结构示意图 Fig. 1Schematic diagram of vector construction for the fusion of NRSE1 element and GUS gene
A: The diagram of PR3b gene and the alternative splicing region, and the first spliced base are separated by 4 bases from the left 64 bases; B: Structural diagram of the vector expressing the fusion of NRSE1 element and GUS gene
A:转基因植株的RT-PCR分子鉴定;B:转基因材料GUS染色鉴定。Ctrl为非转基因野生型烟草对照;WT+为转基因野生型烟草;nic1和nic2为低烟碱突变体转基因材料。PR3b-GUS表示PR3b的NRSE1元件与GUS融合表达 Fig. 3Identification of transgenic plants
A: Identification of transgenic plants by RT-PCR; B: Identification of transgenic plants by GUS staining. Ctrl indicates untransgenic wild type tobacco. WT+ indicates transgenic wild type tobacco; nic1 and nic2 indicate the transgenic plants of low-nicotine mutants. PR3b-GUS indicates the fusion of NRSE1 element and GUS gene
WT+为野生型转基因烟草,nic1和nic2为低烟碱突变体转基因材料,Ctrl为未转基因野生型烟草对照。NRSE1-GUS表示PR3b的NRSE1元件与GUS融合表达。星号为引物二聚体 Fig. 4Molecular splicing analysis of the fusion of NRSE1 element and GUS in nic1 and nic2 mutants
WT+ indicates transgenic wild type tobacco; nic1 and nic2 indicate the transgenic plants of low-nicotine mutants. Ctrl indicates untransgenic wild type tobacco. PR3b-GUS indicates the fusion of NRSE1 element and GUS gene. Asterisk indicates primer dimers
A:转基因烟草的GUS活性检测;B:融合表达基因的分子剪切分析,星号为引物二聚体;C:ET、JA处理后转基因植株的GUS基因相对表达量。WT+为转基因野生型烟草,nic1和nic2为转基因低烟碱突变体材料;Mock为未处理材料,ET和JA分别为乙烯和茉莉酸处理材料 Fig. 5Effects of ET and JA treatment on the GUS activities of transgenic tobacco plants
A: GUS activity of transgenic tobacco; B: Splicing analysis of the fusion of NRSE1 element and GUS. Asterisk indicates primer dimers; C: Relative expression of GUS gene after transgenic plants treated with ET and JA. WT+ indicates transgenic plant of wild type; nic1 and nic2 indicate transgenic plant of low-nicotine mutants. Mock indicates control treatment; ET and JA indicate ethylene and jasmonate treatment, respectively
JALALI BL, BHARGAVAS, KAMBLEA . Signal transduction and transcriptional regulation of plant defence responses Journal of Phytopathology, 2006,154:65-74. [本文引用: 1]
ZHANGY, YANG AG, FENG QF, JIANG CH, GENG RM, LUO CG . Plant pathogenesis-related proteins and research progress in tobacco Biotechnology Bulletin, 2012(5):20-24. (in Chinese) [本文引用: 1]
MARTIN GB, BOGDANOVE AJ, SESSAG . Understanding the functions of plant disease resistance proteins Annual Review of Plant Biology, 2003,54:23-61. [本文引用: 1]
GLAZEBROOKJ . Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens Annual Review of Phytopathology, 2005,43:205-227. [本文引用: 1]
MA HR, WANGF, WANG WJ . Alternative splicing of basic chitinase gene PR3b in the low-nicotine mutants of Nicotiana tabacum L. cv. Burley 21 Journal of Experimental Botany, 2016,67:5799-5809. [本文引用: 4]
RUBéNA, TIBURCIO AF . Determination of arginine and ornithine decarboxylase activities in plants Methods in Molecular Biology (Clifton, N.J.), 2018,1694:117-122. [本文引用: 1]
PATELJ, ARIYARATNEM, AHMEDS, GEL, PHUNTUMARTV, KALINOSKIA, MORRIS PF . Dual functioning of plant arginases provides a third route for putrescine synthesis Plant Science, 2017,262:62-73. [本文引用: 1]
SHUNSUKEI, KATSUHITOH, MAKIKON, HISAEK, YOSHIKATSUM, TAKASHIH, YOUJIS, YASUYUKIY, KENZON . Differential induction by methyl jasmonate of genes encoding ornithine decarboxylase and other enzymes involved in nicotine biosynthesis in tobacco cell cultures Plant Molecular Biology, 1998,38(6):1101-1111. [本文引用: 1]
CHATTOPADHYAY MK, GHOSHB . Molecular analysis of polyamine biosynthesis in higher plants Current Science, 1998,74:517-522. [本文引用: 1]
DALTON HL, BLOMSTEDT CK, NEALE AD, GLEADOWR, DEBOER KD, HAMILL JD . Effects of down-regulating ornithine decarboxylase upon putrescine-associated metabolism and growth in Nicotiana tabacum L Journal of Experimental Botany, 2016,67(11):3367-3381. [本文引用: 1]
KAJIKAWAM, SIERRON, HASHIMOTOT, SHOJIT . A model for evolution and regulation of nicotine biosynthesis regulon in tobacco Plant Signaling & Behavior, 2017,12(6):e1338225. [本文引用: 1]
NACONSIEM, KATOK, SHOJIT . Molecular evolution of N-methylputrescine oxidase in tobacco Plant and Cell Physiology, 2014,55:436-444. [本文引用: 1]
AKIRAK . Molecular biology of pyridine nucleotide and nicotine biosynthesis Frontiers in Bioscience, 2004,9(1-3):1577-1586. [本文引用: 1]
RALPH ED, XIE JH . Molecular genetics of alkaloid biosynthesis in Nicotiana tabacum Phytochemistry, 2013,94:10-27. [本文引用: 1]
DEBOERK, LYEJ, AITKENC, SUA, HAMILLJ . The A622 gene in Nicotiana glauca (tree tobacco): Evidence for a functional role in pyridine alkaloid synthesis Plant Molecular Biology, 2009,69(3):299-312. [本文引用: 1]
KAJIKAWAM, HIRAIN, HASHIMOTOT . A PIP-family protein is required for biosynthesis of tobacco alkaloids Plant Molecular Biology, 2009,69:287-298. [本文引用: 1]
KAJIKAWAM, SHOJIT, KATOA . Vacuole-localized berberine bridge enzyme-like proteins are required for a late step of nicotine biosynthesis in tobacco Plant Physiology, 2011,155:2010-2022. [本文引用: 1]
SHOJIT, HASHIMOTOT . Tobacco MYC2 regulates jasmonate inducible nicotine biosynthesis genes directly and by way of the NIC2 locus ERF genes Plant & Cell Physiology, 2011,52(6):1117-1130. [本文引用: 1]
LEGG PD, CHAPLIN JF, COLLINS GB . Inheritance of percent total alkaloids in Nicotiana tabacum L.: Populations derived from crosses of low alkaloid lines with burley and flue-cured varieties Journal of Heredity, 1969,60:213-217. [本文引用: 1]
HIBIN, HIGASHIGUCHIS, HASHIMOTOT, YAMADAY . Gene expression in tobacco low-nicotine mutants The Plant Cell, 1994,6(5):723-735. [本文引用: 1]
WASTERNACKC, SONGS . Jasmonates: Biosynthesis, metabolism, and signaling by proteins activating and repressing transcription Journal of Experimental Botany, 2017,68(6):1303-1321. [本文引用: 1]
MEMELINK, JOHAN . Regulation of gene expression by jasmonate hormones Phytochemistry, 2009,70(13):1560-1570. [本文引用: 1]
ZHANG HB, BOKOWIEC MT, RUSHTON PJ . Tobacco transcription factors NtMYC2a and NtMYC2b form nuclear complexes with the NtJAZ1 repressor and regulate multiple jasmonate-inducible steps in nicotine biosynthesis Molecular Plant, 2012,5(1):73-84. [本文引用: 1]
SHOJIT, KAJIKAWAM, HASHIMOTOT . Clustered transcription factor genes regulate nicotine biosynthesis in tobacco The Plant Cell, 2010,22(10):3390-3409. [本文引用: 2]
DONGN, ZHANG ZY, XIN ZY . Isolation and expression analysis of a pathogen-induced ERF gene in Triticum aestivum L Scientia Agricultura Sinica, 2008,41(4):946-953. (in Chinese) [本文引用: 1]
BOER KD, TILLEMANS, PAUWELSL . APETALA2/ETHYLENE RESPONSE FACTOR and basic helix-loop-helix tobacco transcription factors cooperatively mediate jasmonate-elicited nicotine biosynthesis The Plant Journal: for Cell and Molecular Biology, 2011,66(6):1053-1065. [本文引用: 1]
GILMOUR SJ, SEBOLT AM, SALAZAR MP . Overexpression of the Arabidopsis CBF3 transcriptional activator mimics multiple biochemical changes associated with cold acclimation Plant Physiology, 2000,124(4):1854-1865. [本文引用: 1]
SHARABI SM, SAMACHA, PORATR . Overexpression of the CBF2 transcriptional activator in Arabidopsis suppresses the responsiveness of leaf tissue to the stress hormone ethylene Plant Biology, 2010,12(4):630-638. [本文引用: 1]
SHOJIT, HASHIMOTOT . Expression of a tobacco nicotine biosynthesis gene depends on the JRE4 transcription factor in heterogenous tomato Journal of Plant Research, 2019,132(2):173-180. [本文引用: 1]
ZHAO YX, XU ZS, CHENM, LI LC, CHEN YF, QIU ZG, XIONG XJ, MA YZ . Analysis of specific binding and subcellular localization of wheat ERF transcription factor W17 Scientia Agricultura Sinica, 2008,41(6):1575-1582. (in Chinese) [本文引用: 1]
ZHANG HB, ZHANG DB, CHENJ, YANG YH, HUANG ZJ, HUANG DF, WANG XC, HUANG RF . Tomato stress-responsive factor TSRF1 interacts with ethylene responsive element GCC box and regulates pathogen resistance to Ralstonia solanacearum Plant Molecular Biology, 2004,55(6):825-834. [本文引用: 1]
CHAKRAVARTHYS, TUORI RP, D'ASCENZO MD, FOBERT PR, DESPRESC, MARTIN GB . The tomato transcription factor Pti4 regulates defense-related gene expression via GCC box and non-GCC box cis elements The Plant Cell, 2003,15(12):3033-3050. [本文引用: 1]
SHOJIT, HASHIMOTOT . Stress-induced expression of NICOTINE2- locus genes and their homologs encoding ethylene response factor transcription factors in tobacco Phytochemistry, 2015,113:41-49. [本文引用: 1]
LEGG PD, COLLINS GB, LITTON CC . Registration of La Burley 21 tobacco germplasm1 registration No. (GP 8) Crop Science, 1970,10(2):212. [本文引用: 1]
 ,11
,11 
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT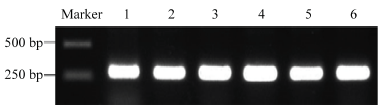 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}