Identification and Expression Analysis of MYB Transcription Factors Related to Rust Resistance in Foxtail Millet
BAI Hui1, SONG ZhenJun2, WANG YongFang1, QUAN JianZhang1, MA JiFang1, LIU Lei1, LI ZhiYong,1, DONG ZhiPing,11 Institute of Millet Crops, Hebei Academy of Agriculture and Forestry Sciences/National Foxtail Millet Improvement Center/Minor Cereal Crops Laboratory of Hebei Province, Shijiazhuang 050035 2 College of Communication and Information Engineering, Shanghai Technical Institute of Electronic & Information, Shanghai 201411;
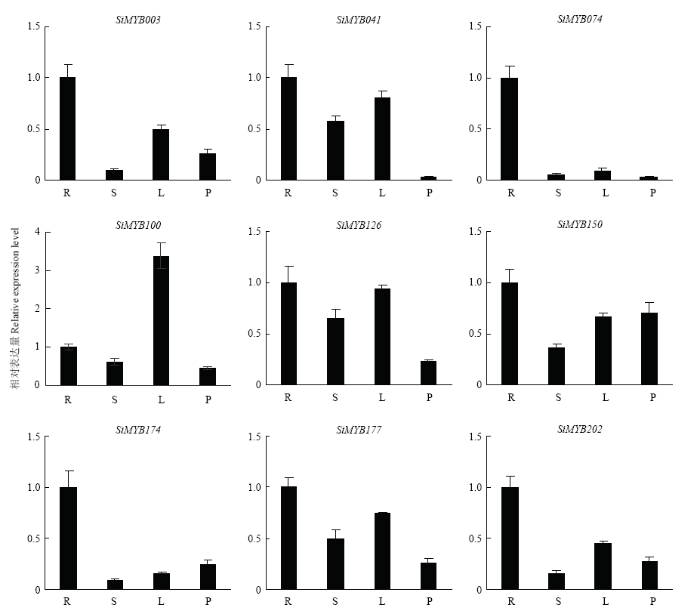
Abstract 【Objective】 Millet rust is one of the important factors affecting the yield and quality of foxtail millet. Identification of MYB transcription factors related to rust resistance in foxtail millet lays a foundation for the study of the mechanism of rust resistance in foxtail millet. 【Method】 In this study, we used real-time PCR to detect the expression patterns of 9 MYB transcription factors (1) in roots, stems, leaves and panicles at booting stage; (2) during 120 hours post-inoculated with Uromyces setariae-italicae urediniospores in resistance (R) and susceptible (S) reactions; (3) during 24 hours after treatment with salicylic acid (SA) and methyl jasmonate (MeJA) in foxtail millet. Then their expression patterns in four reactions of R, S, SA and MeJA were compared, and the resistance-related MYB transcription factors were selected for detection of transactivation activity and subcellular localization. 【Result】 The highest expression of SiMYB100 was in leaves, and the highest expression of the other eight genes, especially SiMYB074, was in roots. The expression of five genes was correlated with disease resistance. SiMYB074 and SiMYB202 were induced by rust fungus infection and their expression levels at early stage of disease resistance were significantly higher than that in the susceptible reaction. The expression of SiMYB041 and SiMYB177 was down-regulated in the early stage of resistance reaction and increased to pre-inoculation level in the later stage, while their expression remained low in susceptible reaction. SiMYB100 showed opposite expression pattern in resistance and susceptible responses. In response to exogenous SA and MeJA, the expression of SiMYBs gene changed in varying degrees. The expression patterns of four genes (SiMYB074, SiMYB100, SiMYB174 and SiMYB202) in the R, SA and MeJA reactions were identical, but different from the S reaction. Five resistance-related SiMYBs genes have the transactivation activity and their encoding proteins are located in the nucleus. 【Conclusion】 The expression of five genes, SiMYB041, SiMYB074, SiMYB100, SiMYB177 and SiMYB202, was identified to be associated with resistance to rust disease in foxtail millet. SiMYB074 and SiMYB100 play certain roles in the growth and disease resistance of foxtail millet. Four genes, SiMYB074, SiMYB100, SiMYB174 and SiMYB202, may participate in early disease resistance of foxtail millet through SA and JA signaling pathways. Keywords:Setaria italica;MYB transcription factors;rust disease;real-time PCR;disease resistance
PDF (1303KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 白辉, 宋振君, 王永芳, 全建章, 马继芳, 刘磊, 李志勇, 董志平. 谷子抗锈病反应相关MYB转录因子的鉴定与表达[J]. 中国农业科学, 2019, 52(22): 4016-4026 doi:10.3864/j.issn.0578-1752.2019.22.007 BAI Hui, SONG ZhenJun, WANG YongFang, QUAN JianZhang, MA JiFang, LIU Lei, LI ZhiYong, DONG ZhiPing. Identification and Expression Analysis of MYB Transcription Factors Related to Rust Resistance in Foxtail Millet[J]. Scientia Acricultura Sinica, 2019, 52(22): 4016-4026 doi:10.3864/j.issn.0578-1752.2019.22.007
ZHAO LQ, PAN WJ, MA JF, WENG QY, DONGL, QUAN JZ, XING JH, DONG ZP, DONG JG . Identification of AFLP markers linked to a novel rust resistance gene in foxtail millet Scientia Agricultura Sinica, 2010,43(21):4349-4355. (in Chinese) [本文引用: 1]
DUBOSC, STRACKER, GROTEWOLDE, WEISSHAARB, MARTINC, LEPINIECL . MYB transcription factors in Arabidopsis. Trends in Plant Science, 2010,15(10):573-581. [本文引用: 2]
ZUOR, XU ML, CHAI GH, ZHOU GK . Function and regulation mechanism of plant MYB transcription factors Chinese Bulletin of Life Sciences, 2012,24(10):1133-1140. (in Chinese) [本文引用: 1]
CHEN YH, YANG XY, HEK, LIU MH, LI JG, GAO ZF, LIN ZQ, ZHANG YF, WANG XX, QIU XM, SHEN YP, ZHANGL, DENG XH, LUO JC, DENG XW, CHEN ZL, GU HY, QU LJ . The MYB transcription factor superfamily of arabidopsis: expression analysis and phylogenetic comparison with the rice MYB family Plant Molecular Biology, 2006,60(1):107-124. [本文引用: 1]
MATUS JT, AQUEAF, ARCE-JOHNSONP . Analysis of the grape MYB R2R3 subfamily reveals expanded wine quality-related clades and conserved gene structure organization across Vitis and Arabidopsis genomes. BMC Plant Biology, 2008. 8:83. [本文引用: 1]
SUO JF, LIANG XE, PUL, ZHANG YS, XUE YB . Identification of GhMYB109 encoding a R2R3 MYB transcription factor that expressed specifically in fiber initials and elongating fibers of cotton (Gossypium hirsutum L.). Biochimica et Biophysica Acta, 2003,1630(1):25-34. [本文引用: 1]
MUTHAMILARASANM, KHANDELWALR, YADAV CB, BONTHALA VS, KHANY, PRASADM . Identification and molecular characterization of MYB transcription factor superfamily in C4 model plant foxtail millet (Setaria italica L.). PLoS ONE, 2014,9(10):e109920. [本文引用: 6]
NIU YL, JIANG XM, XU XY . Research advances on transcription factor MYB gene family in plant Molecular Plant Breeding, 2016,14(8):2050-2059. (in Chinese) [本文引用: 1]
LEE MM, SCHIEFELBEINJ . Cell pattern in the Arabidopsis root epidermis determined by lateral inhibition with feedback. The Plant Cell, 2002,14(3):611-618. [本文引用: 1]
RYU KH, KANG YH, PARK YH, HWANGI, SCHIEFELBEINJ, LEE MM . The WEREWOLF MYB protein directly regulates CAPRICE transcription during cell fate specification in the Arabidopsis root epidermis. Development, 2005. 132(21):4765-4775. [本文引用: 1]
NIU YF . Effect of elevated carbon dioxide on root hair development and nutrient uptake and the response mechanisms of root growth to nutrient supply in Arabidopsis thaliana [D]. Hangzhou: Zhejiang University, 2013. (in Chinese) [本文引用: 1]
ZHANG DJ, XIA RX, CAOX . Root hair development and its genetic basis Plant Physiology Journal, 2015,51(1):9-20. (in Chinese) [本文引用: 1]
BYRNE ME, BARLEYR, CURTISM, ARROYO JM, DUNHAMM, HUDSONA, MARTIENSSEN RA . Asymmetric leaves1 mediates leaf patterning and stem cell function in Arabidopsis Nature, 2000,408(6815):967-971. [本文引用: 1]
CHENGH, SONGS, XIAOL, SOO HM, CHENGZ, XIED, PENGJ . Gibberellin acts through jasmonate to control the expression of MYB21, MYB24, and MYB57 to promote stamen filament growth in Arabidopsis PLoS Genetics, 2009,5(3):e1000440. [本文引用: 1]
VAILLEAUF, DANIELX, TRONCHETM, MONTILLET JL, TRIANTAPHYLIDèSC, ROBYD . A R2R3-MYB gene, AtMYB30, acts as a positive regulator of the hypersensitive cell death program in plants in response to pathogen attack. Proceedings of the National Academy of Sciences of the United States of America, 2002,99(15):10179-10184. [本文引用: 2]
AL-ATTALA MN, WANGX, ABOU-ATTIA MA, DUANX, KANGZ . A novel TaMYB4 transcription factor involved in the defence response against Puccinia striiformis f. sp. tritici and abiotic stresses. Plant Molecular Biology, 2014,84(4/5):589-603. [本文引用: 1]
LIUH, ZHOUX, DONGN, LIUX, ZHANGH, ZHANGZ . Expression of a wheat MYB gene in transgenic tobacco enhances resistance to Ralstonia solanacearum, and to drought and salt stresses. Functional & Integrative Genomics, 2011,11(3):431-443. [本文引用: 1]
SHAN TL, HONG YT, DU LP, XU HJ, WEI XN, ZHANG ZY . Development and characterization of TaMYB86-overexpressing transgenic wheat lines with resistance to common root rot. Acta Agronomica Sinica, 2016,42(10):1429-1436. (in Chinese) [本文引用: 1]
CAO WL, CHU RZ, ZHANGY, LUOJ, SU YY, XIE LJ, ZHANG HS, WANG JF, BAO YM . OsJAMyb, a R2R3-type MYB transcription factor, enhanced blast resistance in transgenic rice Physiological and Molecular Plant Pathology, 2015,92:154-160. [本文引用: 1]
LIUZ, LUAN YS, LI JB, YIN YL . Expression of a tomato MYB gene in transgenic tobacco increases resistance to Fusarium oxysporum and Botrytis cinerea. European Journal of Plant Pathology, 2016,144(3):607-617. [本文引用: 1]
DONGJ, CHENC, CHENZ . Expression profiles of the Arabidopsis WRKY gene superfamily during plant defense response. Plant Molecular Biology, 2003,51(1):21-37. [本文引用: 1]
CHEN NL, HUM, QIAO CP, NAI XY, WANGR . Effects of BTH, SA, and SiO2 treatment on disease resistance and leaf HRGP and lignin contents of melon seedlings Scientia Agricultura Sinica, 2010,43(3):535-541. (in Chinese) [本文引用: 1]
NIY, WANGJ, SONGC, XIA RE, SUN ZY, GUO YJ, LI JN . Effect of SA induction on leaf cuticular wax and resistance to Sclerotinia sclertiorurn in Brassica napus. Acta Agronomica Sinica, 2013,39(01):110-117. (in Chinese) [本文引用: 1]
NIU JS, LIUR, ZHENGL . Expression analysis of wheat PR-1, PR-2, PR-5 activated by Bgt and SA, and powdery mildew resistance. Journal of Triticeae Crops, 2007,27(6):1132-1137. (in Chinese) [本文引用: 1]
NIU JS, LIUJ, NI YJ, YINJ . Induction of PR-1, PR-2, PR-5, Ta-JA2 and wheat powdery mildew resistance in response to MeJA treatment. Acta Phytopathologyca Sinica, 2011,41(3):270-277. (in Chinese) [本文引用: 1]
NIU JS, NI YJ, LIUJ, WANG ZY, YINJ . Inducing effect of JA on wheat powdery mildew resistance Chinese Agricultural Science Bulletin, 2010,26(4):254-257. (in Chinese) [本文引用: 1]
PENG XX, HU YJ, TANG XK, ZHOU PL, DENG XB, WANG HH . Isolation and expression profiles of rice WRKY30 induced by jasmonic acid application and fungal pathogen infection. Scientia Agricultura Sinica, 2011,44(12):2454-2461. (in Chinese) [本文引用: 1]
ZHANGY, XUS, DINGP, WANGD, CHENGYT, HEJ, GAOM, XUF, LIY, ZHUZ, LIX, ZHANGY . Control of salicylic acid synthesis and systemic acquired resistance by two members of a plant-specific family of transcription factors Proceedings of the National Academy of Sciences of the United States of America, 2010,107(42):18220-18225.
WANG MF . The role of WRKY transcription factors in the elicitor-triggered hypersensitive response and stomatal closure in Nicotiana benthamiana [D]. Nanjing: Nanjing Agricultural University, 2011. (in Chinese) [本文引用: 1]
LIANG KG, LIUW, WANG YR, FENG LY, CUI GX, SONG YC, WU XF, ZHENG GC, DONG ZP . Rust resistance evaluation for millet varieties Journal of Shenyang Agricultural University, 1992,23(1):13-18. (in Chinese) [本文引用: 1]
YOO SD, CHO YH, SHEENJ . Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis Nature Protocols, 2007,2(7):1565-1572. [本文引用: 1]
BILAUDT, KOERING CE, BINET-BRASSELETE, ANCELINK, POLLICEA, GASSER SM, GILSONE . The telobox, a Myb-related telomeric DNA binding motif found in proteins from yeast, plants and human Nucleic Acids Research, 1996,24(7):1294-1303. [本文引用: 1]
CHENJ, WANG ZY . Progress in the study of plant MYB transcription factors Journal of Plant Physiology and Molecular Biology, 2002,28(2):81-88. (in Chinese) [本文引用: 1]
LIBAULTM, BRECHENMACHERL, CHENGL, XU JL, XUD, STACEYG . Root hair systems biology Trends in Plant Science, 2010,15(11):641-650. [本文引用: 1]
WANG LJ, RAN LY, HOU YS, TIAN QY, LI CF, LIUR, FAND, LUO KM . The transcription factor MYB115 contributes to the regulation of proanthocyanidin biosynthesis and enhances fungal resistance in poplar New Phytologist, 2017,215(1):351-367. [本文引用: 1]
VLEESSCHAUWER DD, GHEYSENGODELIEVE, H?FTEMONICA . Hormone defense networking in rice: Tales from a different world Trends in Plant Science, 2013,18(10):555-565. [本文引用: 1]
YANG YN, QIM, MEI CS . Endogenous salicylic acid protects rice plants from oxidative damage caused by aging as well as biotic and abiotic stress The Plant Journal, 2004,40(6):909-919. [本文引用: 1]
HAMMOND-KOSACK KE, PARKER JE . Deciphering plant- pathogen communication: Fresh perspectives for molecular resistance breeding Current Opinion in Biotechnology, 2003,14(2):177-193. [本文引用: 1]
LIU XQ, LIF, TANG JY, WANG WH, ZHANG FX, WANG GD, CHU JF, YAN CY, WANG TQ, CHU CC, LI CY . Activation of the jasmonic acid pathway by depletion of the hydroperoxide lyase OsHPL3 reveals crosstalk between the HPL and AOS branches of the oxylipin pathway in rice PLoS ONE, 2012,7(11):e50089. [本文引用: 1]
TONG XH, QI JF, ZHU XD, MAO BZ, ZENG LJ, WANG BH, LIQ, ZHOU GX, XU XJ, LOU YG, HE ZH . The rice hydroperoxide lyase OsHPL3 functions in defense responses by modulating the oxylipin pathway The Plant Journal, 2012,71(5):763-775. [本文引用: 1]
 ,1, 董志平
,1, 董志平
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT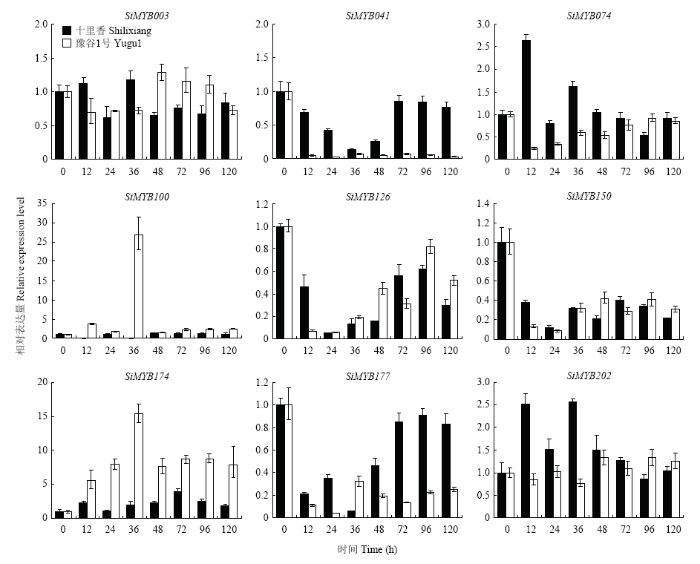 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT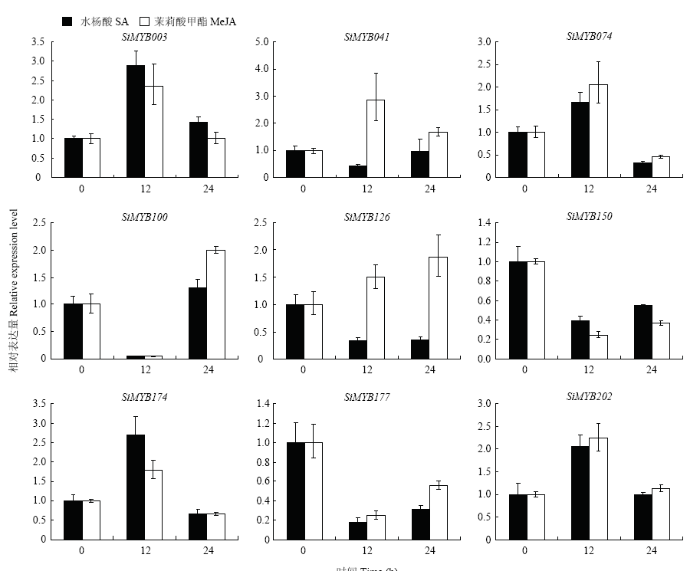 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT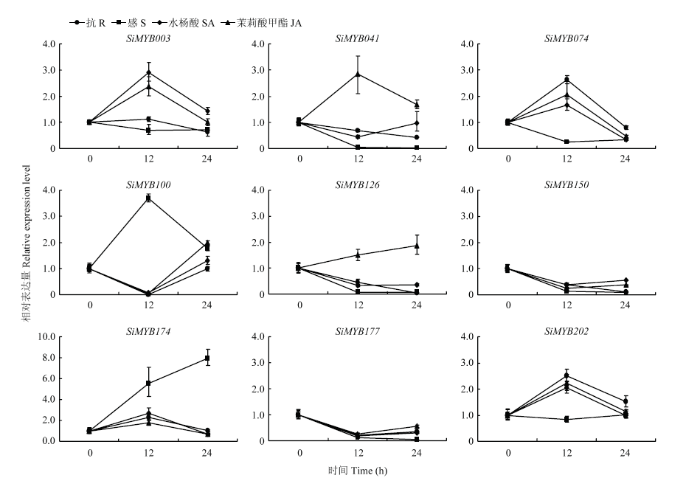 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT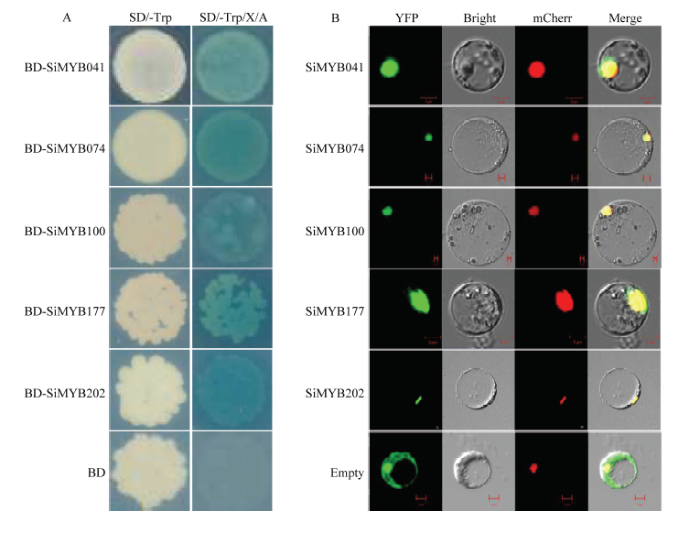 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}