 ,1, �Ž���,1
,1, �Ž���,1Enzymatic Characteristics and Metabolic Analysis to Malathion and p,p��-DDT of LmGSTS2 from Locusta migratoria
MA Wen1, LIU Jiao2, ZHANG XueYao2, SHEN GuoHua3, QIN XueMei,1, ZHANG JianQin,1ͨѶ����:
�ո�����:2018-11-23��������:2018-12-27�����������:2019-04-16
| ��������: |
Received:2018-11-23Accepted:2018-12-27Online:2019-04-16
����� About authors
����,E-mail��

ժҪ
�ؼ��ʣ�
Abstract
Keywords��
PDF (1008KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر���
�������ø�ʽ
����, ����, ��ѧҢ, �����, ��ѩ÷, �Ž���. �ɻ�LmGSTS2��øѧ���Լ�����������ס� p,p��-DDT�Ĵ�л����[J]. �й�ũҵ��ѧ, 2019, 52(8): 1389-1399 doi:10.3864/j.issn.0578-1752.2019.08.009
MA Wen, LIU Jiao, ZHANG XueYao, SHEN GuoHua, QIN XueMei, ZHANG JianQin.
0 ����
���о����塿�ɻȣ�Locusta migratoria����һ����Ҫ��ũҵ����,��Ҫ�Ժ̱��ƺ�ɯ�ݿ�ֲ���С��ˮ���ȣ�Ϊʳ,����Σ���ҹ�����������ũҵ����[1]���ڿ��ƻ��ֹ�����,��ͬ����ɱ�����ʹ��,������Ⱦ��ũ���P����,����ɱ�����������Ӱ�����ཡ��,ͬʱʹ�ɻȶ�ɱ����Ŀ�������ǿ[2]���������ף�malathion��Ϊһ�ָ�Ч���Ͷ����л���ɱ���,��ũҵ�����б��㷺ʹ��[3];�ε��飨chlorophenothane,DDT��Ϊһ�ָ�Ч���߶����л���ɱ���,�����䲻����,Ŀǰ�ѽ�ֹ��ũҵ������Ӧ��,�����������Ȼ����IJ����ѵ�����ҩ�����л���ũҩ�IJ���[4]��������ǰ���о�����,�ɻ�����������Ʒϵ�Ĺ�������ת��ø��glutathione-S- transferases,GST,E.C.2.5.1.18���Ļ����������[5]����������ת��ø��һ����Ҫ�Ģ����лø,�ɲ��������������ʴ�л���������ܵĵ���[6]������GST�Ĵ��ڲ�λ��ҪΪ֬���塢�г�����Ƥ�����Ϲܵ���֯��,������Ҫ�ⶾ���ٵ���֯������ظ߱�����ʾGST����DZ�ڵĽⶾ����[7,8,9]���������о�����,��������GST����ͨ�����Ȼ�������,���߶���DDT��лΪ����DDE,�Լ�СDDT���������Ķ������á����,��ȷ�ɻ����ڽⶾøGST��ɱ����Ĵ�л����,һ������ڽ������͵ĺ��������ϵ���ٽ�������ɫ�Ѻü��ɳ�����չ������Ҫ����[10],��һ������ڿ������ڽⶾø�Ļ��̾�������ҩ��ũ���ѳ������о�������Ҫ����[4]����ǰ���о���չ������GST�����������������ԡ����������Ժ�����ѧ����,����GST��Ϊ6�ࣺsigma��epsilon��delta��theta��omega��zeta[11,12,13]��Ŀǰ�Ѿ���������sigma��epsilon��delta�����GST�㷺��������Զ���ɱ����Ĵ�л����[14,15,16]��RIVERON�Ƚ��������ã�Anopheles funestus��GSTe2�ڹ�Ӭ�й������,��ǿ�˹�Ӭ��DDT���Ⱦ����Ŀ���,ͨ��������P����֤ʵGSTe2��ֱ�Ӷ��Ⱦ������д�л,��ͨ��L119F��ͻ���һ��֤ʵ��GSTe2����DDT�Ľⶾ����[17];LUMJUAN�ȷ��ְ������ã�Aedes aegypti����GSTe2��GSTe5�������Ȼ���ø����,�ɽ�DDT��лΪ����DDE;ͨ��RNAi����֤ʵGSTe7��GSTe2���ܲ�������������ɱ����Ĵ�л����[18]��Ŀǰ�ɻ�����֪��GST����32��,���а���GST��28��,����10��sigma����GST,5��epsilon����GST,7��delta����GST,3��omega����GST,2��theta����GST��1��zeta����GST[7]��QIN����������������ɻ�48 h����������GST���������ֶ���sigma����GST���������ϵ�,˵��sigma����GST�ܿ��ܹ�ͬ�����˷ɻ�������������Ĵ�л�ⶾ[19]������,ͨ��RNAi��ĬLmGSTs5��,�ɻȶ�������������ά������ж�����,�����ʷֱ�����28%��26%;��ĬLmGSTs3��,��ά���������������38.7%,����sigma����GST���ܲ�������������ά��Ľⶾ����[20,21]�������о�����㡿Sigma����GST�ڸ���ɱ����Ĵ�л�ⶾ�����о���ͬ�̶ȷ���������,���ɻ�����ȱ����GST�ⶾø���ں������л�ⶾ������ϵͳ�о���ZHANG��[7]ͨ��RT-qPCR�о�����LmGSTs2��HM131837.1�����г������Ϲܺ�֬����ȸ��������ͽⶾ����֯�б��������,���,����ѡ��ɻ�sigma�����е�LmGSTS2��AEB91974.1�������о�,������������ﲢ����,ͨ������ЧҺ��ɫ�ף�ultra performance liquid chromatography,UPLC�����LmGSTS2��ɱ�����������p,p��-DDT�Ĵ�л����,���ڷɻ����ڶԸý��������֤���������Ĺؼ����⡿LmGSTS2�����,̽�����ʵĴ���������øߴ��ȵĵ���,���ó���ЧҺ��ɫ�ֱ����������p,p��-DDT���м��,����LmGSTS2��ɱ����Ĵ�л��������һ��ͨ��RNAi��Ĭ�ɻ����ڵ�LmGSTs2,������ɱ��������жȱ仯,�Ӷ�ͨ�����ڡ���������ϵͳ����LmGSTS2��ɱ����Ĵ�л�ⶾ���á�1 �����뷽��
������2017��2018����ɽ����ѧ��ҽҩ�ִ��о����ļ�Ӧ������ѧ�о�����ɡ�1.1 �������������Լ�
������?KTATM pure���״���ϵͳ����GE Healthcare��˾;Synergy MX Microplate Readerø���ǹ���BioTek��˾;1290 ��ЧҺ��ɫ���ǹ���Agilent��˾;LightCycler? 480����ROCHE��˾�����ϣ�pET28a/BL21(DE3)-LmGSTS2����Ϊʵ����ǰ�ڹ���[22]���Լ���T7 RNA�ϳ��Լ��й���NEB��˾;SYBR? Green real-time PCR Master Mix����TOYOBO��˾;2��Taq PCR Master Mix���������˾;�������ס�p,p��-DDT��Ʒ����CATO��˾��1.2 LmGSTS2��ԭ�˱��P����
�������Һ�����ڹ���LB�������и��ա����պ�,��ȡ�����������Һ��LB����������50 ��g��mL-1����ù�أ��Թ���,��37�桢200 r/min����������������14��16 h,��Ϊ���Ӿ�Һ����1�s100�ı�����ȡ���Ӿ�Һ������Һ��LB�������У���50 ��g��mL-1����ù�أ�,����37��������OD600��0.6,����IPTG����Ũ��Ϊ0.5 mmol��L-1��,����16������,�յ�Ŀ�군�״�������;20 h��,�����ռ��������,��PBS������PBS��Ϊ0.1 mol��L-1,pH 7.0����ϴ3��;����PBS 20 mL,���ڱ��ϳ�����������Һ����12 000 r/min,4������30 min,�ռ����弴Ϊ��øҺ����SDS-�۱�ϩ����������Ӿ��SDS-PAGE������LmGSTS2�ɹ������,ʹ��HisTrap FFԤװ��,ͨ��?KTATM pure���״���ϵͳ���д������ú��в�ͬŨ������60��250 mmol��L-1���Ļ���Һϴ�ѡ��Դֵ�����ͬϴ����ֽ��е�Ӿ���,�Ժ��нϵ�һĿ��������ϴ�����ͨ��PBS����Һ���г���,-20�涳�汸�á�
1.3 LmGSTS2�����������ⶨ
1.3.1 ø���Բⶨ ����HABIG��[23]�ķ�������LmGSTS2���Բⶨ��ȡ10 ��L����øҺ��190 ��L���Һ��5 mmol��L-1 GSH�s200 mmol��L-1 CDNB=188�s2��,������ø������340 nm����¼ODֵ��ÿ�� 10 s����,������1 min����10 ��L PBS����Һ��Ϊ�հ���,��Ʒ�ⶨʱ����3���ظ���CDNB��Ħ������ϵ��Ϊ��340=9.6 mmol��L-1��cm-1��1.3.2 pH��LmGSTS2���Ե�Ӱ�� ����pH Ϊ3��9�Ļ���Һ,����1.3.1��CDNBΪ����ⶨLmGSTS2�ڲ�ͬpH����Һ�еĻ��ԡ���PBS��Ϊ�հ���,��Ʒ�ⶨʱ����3���ظ���
1.3.3 �¶ȶ�LmGSTS2���Ե�Ӱ�� ȡLmGSTS2øҺ��װ�ڶ��PCR����,�ֱ�����20��30��40��50��60�����ˮԡ���б���30 min,4�汣�档����1.3.1�ⶨ��ͬ�¶ȴ�����LmGSTS2���ԡ���Ʒ�ⶨʱ����3���ظ���
1.4 LmGSTS2����������p,p��-DDT�Ĵ�л���
1.4.1 ����ЧҺ��ɫ�����Ľ��� ��Ʒ���ɫ����������1������ɱ���ϴ�ѷ�ʽ��Ϊ�ȶ�ϴ��,������AΪ����,BΪ0.1%����ˮ��Table 1
��1
��1����ЧҺ��ɫ���ɱ�������
Table 1
| ɱ��� Insecticide | ������ Mobile phase (A%) | ϴ��ʱ�� Elution time (min) | ��Ⲩ�� Detection wavelength (nm) | ������ Sample application (��L) | ���� Temperature (��) |
|---|---|---|---|---|---|
| ��������Malathion | 80 | 8 | 210 | 2 | 35 |
| p,p��-DDT | 92 | 10 | 232 | 2 | 35 |
�´��ڴ�|����CSV
1.4.2 ����LmGSTS2���Բ�ͬɱ����Ĵ�л�ⶾ���ܷ��� ��Ʒ��ΪA��B��C��D 4�顣A��Ϊ������Ӧ�飨active LmGSTS2+GSH+insecticide��;B����PBS����Һ���浰��,Ϊ�������飨GSH+ insecticide��;C��Ϊ����GSH�Ķ����飨active LmGSTS2+insecticide��;D�齫��к�ĵ�����Ϊ�����飨inactive LmGSTS2+GSH+insecticide�������Ⱥ�,����27��,1 200 r/min,����3 h;��Ӧ���������400 ��L����,27��,1 200 r/min,����5 min��ֹ��Ӧ;����Ʒ��13 000 r/min,��������20 min,ȡ����Һ��0.22 ��m����Ĥ���˺�ת����1.5 mLɫ��ƿ,����1.4.1�������м�⡣ÿ����Ʒ��ƽ���Ʊ�3��,֮��Բ�ͬ��ɱ���������仯ͨ��Student��s t-test���������Է�����ÿ����Ʒ�Ʊ�ʱ����2���ظ���
1.5 LmGSTs2�����Ĭ��ɻȶ������������жȷ���
1.5.1 LmGSTs2 PCR������dsRNA�ϳ� ������֪��LmGSTs2��������,ʹ��Primer Premier5.0 �������dsRNA�ϳ����PRT-qPCR����,�������ϸ��Ϣ����2,������������������﹫˾�ϳɡ�Table 2
��2
��2LmGSTs2 dsRNA���PRT-qPCR��������
Table 2
| ���� Primer | ��������Sequence of primer (5��-3��) |
|---|---|
| dsLmGSTs2F | taatacgactcactatagggGAAGAAGACATGGCAGTCTC |
| dsLmGSTs2R | taatacgactcactatagggCTGGTATTTCAAGTACAGTACGT |
| RT-LmGSTs2F | ATGGCGCCAAAATTCAAGTTAC |
| RT-LmGSTs2R | TCACAGAGGCCACTGCTGTAGA |
�´��ڴ�|����CSV
�Ժ���Ŀ�Ļ���ȫ��cDNA ���еľ�ҺΪģ��,��dsRNA��������˫��DNA��ȡPCR����,��1%����֬��������Ӿ���Ŀ��Ƭ�δ�С,�����л��մ���������T7 RNA�ϳ��Լ�˵����ϳ�dsLmGSTs2,����Ũ�ȵ���Ϊ1.5 ��g����L-1����,���á�
1.5.2 dsLmGSTs2 24 h����Ч�ʼ�� ѡȡ�ɻ�2���3���С��һ������״�����õ�����Ϊ�Գ档ʹ����ע�����ص�2��3���ڽڼ�Ĥ��ע��,������ע��2 ��L��3 ��g��Ŀ�Ļ���dsRNA,������ע��2 ��L ddH2O,ÿ��ע��12ͷ����,24 h���ռ��ɻ�,3ͷΪһ������ѧ�ظ�,��4������ѧ�ظ�����ȡRNAͨ��RT-qPCR���dsLmGSTs2��ĬЧ�ʡ�
RT-qPCR��Ӧ��ϵ��20 ��L����4.0 ��L cDNA��10��ϡ�ͣ�,�ϡ����������0.4 ��mol��L-1,10 ��L SYBR? Green real-time PCR Master Mix,���������ddH2O���㡣��Ӧ����95�� 1 min;40 cycles��95�� 15 s,60�� 31 s;95�� 1 min,55�� 1 min,97�� continuous;37�� 30 s���ڲλ���Ϊrpl32��
1.5.3 dsLmGSTs2��Ĭ��ɻȶ������������жȱ仯��� �����Ϊ���飺�����飨ע��2 ��L ddH2O��,�����飨ע��3 ��g dsLmGSTs2��,ÿ��ע��50ͷ2������,ÿ������ѧ�ظ�12��14ͷ,��3������ѧ�ظ�����ע��dsRNA���24 h,ʹ����Һǹ�ڳ��帹�������2��3���ڵ��Ҳ����������ף�0.1 mg��mL-1��3 ��L�����ɱ������24 h,��¼�������������������Student��s t-test�����ݽ���ͳ�Ʒ�����
2 ���
2.1 LmGSTS2��ԭ�˱��P����
����SDS-PAGE�����LmGSTS2�ı���,�����ʾ�����BL21(DE3)����̬ϸ����תȾpET-28a�������BL21(DE3)����̬ϸ����δ�յ���pET28a/ BL21(DE3)-LmGSTS2,�յ����pET28a/BL21(DE3)- LmGSTS2��25 kD���ҳ���һ�����Ե���������ɫ��ͷ��ָλ�ã�,ΪLmGSTS2Ŀ��������ͼ1-A����Ni-NTA�Ͳ�����ͬϴ����ֵ�Ӿ�����ʾ,�ڴ���ʱ,Ӿ��2��������Һ������Ŀ������,˵��LmGSTS2������Ni����ȫ���;����ͬŨ������Һϴ�Ѻ�,�ӵ�����Ҫ��60��200 mmol��L-1����Һϴ����֣�3��8Ӿ������ϴ��,Ŀ�군�ף���ɫ��ͷ��ָλ�ã���250 mmol��L-1����Һϴ����֣�9��12Ӿ������ϴ��,������һ,�����ں������Լ�⣨ͼ1-B����
ͼ1
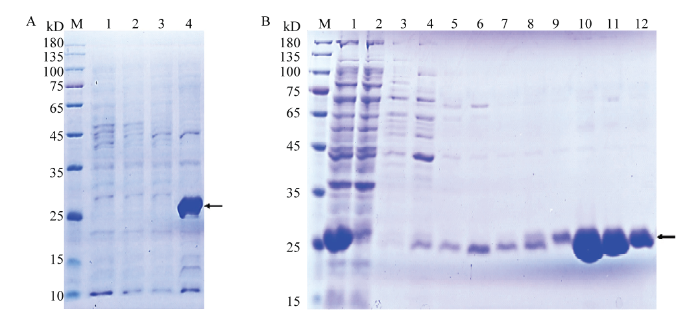 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ1LmGSTS2�ı��P����
Fig. 1Expression and purification of LmGSTS2
M��10��180 kD���������� 10-180 kD protein molecular weight standards��A��LmGSTS2�ı��� Expression of LmGSTS2��1��BL21(DE3);2��pET28a/BL21(DE3);3��δ�յ�pET28a/BL21(DE3)-LmGSTS2 pET28a/BL21(DE3)-LmGSTS2 before induction;4���յ����pET28a/BL21(DE3)-LmGSTS2 pET28a/BL21(DE3)-LmGSTS2 after induction��B��LmGSTS2�Ĵ��� Purification of LmGSTS2�� 1��δ������LmGSTS2�ܵ���Total protein of LmGSTS2 before purification;2��������Һ Flow through fraction;3��12����ͬŨ������Һϴ�����Elution components with different concentrations of imidazole buffer
2.2 LmGSTS2ø���Ե����������ⶨ
��������ĵ���ͨ�����˽����ѳ�����NaCl,��1-��-2,4-����������1-chloro-2,4-dinitrobenzene,CDNB��Ϊ�����о�LmGSTS2�ڲ�ͬ�����µ�ø���Ա仯,��OriginPro 8.0����������ϣ�����B-Spline���ӷ�ʽ��,������ͼ2����ͼ2-A��֪,LmGSTS2����pH��ΧΪ6��8,pH=7ʱ������ߡ���pH<5ʱ����û�л���,pH����5��7ʱ����pH����������������,pH>7ʱ����pH������ø�������͡���ͼ2-B��֪,��25��30�� LmGSTS2���Խϸ�, 27������ʱ�������,�¶�>30��ʱ��������,��50��ʱ������ȫɥʧ���ԡ����,LmGSTS2�����ʷ�Ӧ����ΪpH=7,27�����ҡ�ͼ2
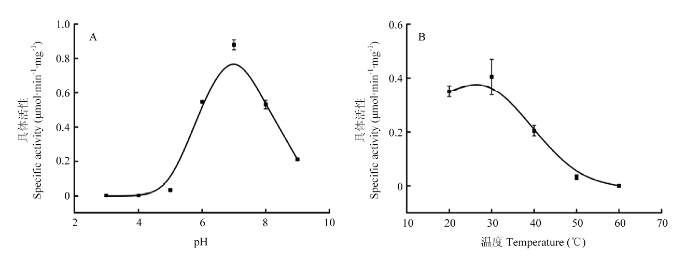 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ2��ͬ������LmGSTS2���Ա仯
Fig. 2Activity changes of LmGSTS2 under different conditions
2.3 LmGSTS2��ɱ����Ĵ�л���
��LmGSTS2����������Ӧ,ͨ��UPLC��ⲻͬ����л��������B��D���,A������������ɫ�彵�ͣ�����ʱ��2.13 min����ͼ3��,��A�������ֱ�83.6%��84.0%��84.6%,����������P<0.05������3������B��C��D 3���������������������Ա仯,˵����3 h��Ӧʱ����,LmGSTS2��GSH��ͬ����,�ɽ����������״�л������ɫ��ͼ��δ���ֲ���塣ͼ3
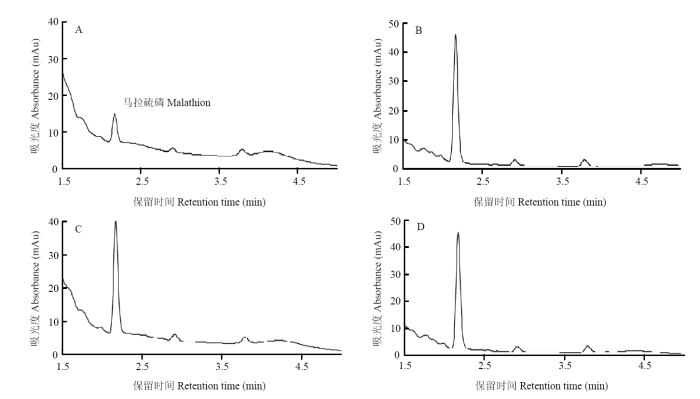 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ3LmGSTS2���������״�лɫ��ͼ
Fig. 3Chromatograms of malathion metabolized by LmGSTS2
A: Active LmGSTS2+GSH+malathion; B: GSH+malathion; C: Active LmGSTS2+ malathion; D: Inactive LmGSTS2+GSH+malathion
Table 3
��3
��3��ͬ���ɱ���ɫ�����
Table 3
| A | B | C | D | |
|---|---|---|---|---|
| ����������� Peak area of malathion | 34.98��4.20 | 213.40��31.80* | 218.47��15.19* | 226.76��19.16* |
| p,p��-DDT����� Peak area of p,p��-DDT | 177.80��14.88 | 169.34��8.95 | 193.07��7.19 | 178.57��13.63 |
�´��ڴ�|����CSV
��LmGSTS2��p,p��-DDT��Ӧ,ͨ��UPLC��ⲻͬ����л��������B��D���,A����p,p��-DDT������ʱ��3.14 min���������Բ���,��û���µIJ������֣�ͼ4��,����������������仯,����LmGSTS2����лp,p��-DDT����3����
ͼ4
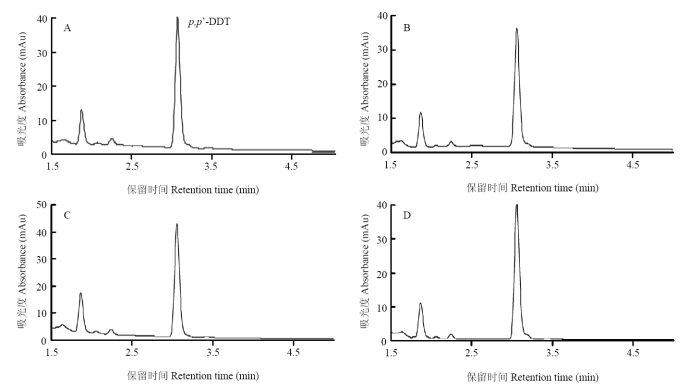 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ4LmGSTS2��p,p��-DDT��лɫ��ͼ
Fig. 4Chromatograms of p,p��-DDT metabolized by LmGSTS2
A: Active LmGSTS2+GSH+ p,p��-DDT; B: GSH+ p,p��-DDT; C: Active LmGSTS2+ p,p��-DDT; D: Inactive LmGSTS2+GSH+ p,p��-DDT
2.4 LmGSTs2�����Ĭ��ɻȶ������������жȷ���
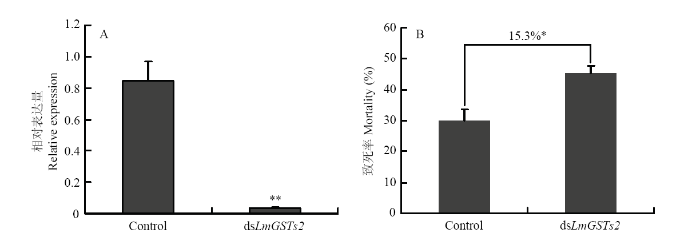
��2��ɻ�����ע��3 ��g dsLmGSTs2,24 h�������ĬЧ��,�����ʾ����������,������LmGSTs2 ��������������,Ŀ�����ij�ĬЧ�ʴ�96%��ͼ5-A����ͼ5
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ5dsLmGSTs2��ĬЧ�ʼ�LmGSTs2��Ĭ��ɻȶ������������жȷ���
Fig. 5Silencing efficiency and L. migratoria sensitivity to malathion after LmGSTs2 expression suppression
A��dsLmGSTs2��ĬЧ�� Silencing efficiency of dsLmGSTs2��**: P<0.01 (t-test);B���ɻȶ������������жȷ��� Sensitivity of L. migratoria to malathion��*: P<0.05 (t-test)
��Ĭ�ɻ����ڵ�LmGSTs2��,�����������, 24 h����ɻ����жȱ仯,�����ʾ�б���� LmGSTs2��Ĭ��,�ɻ���������29.9%������45.2%��ͼ5-B����
3 ����
�ⶾø�����ⳬ������ڸ��õ��о�øѧ���ʼ����ɱ����Ĵ�л���þ�����Ҫ���塣WAN��[24]�����ҹ�꣨Spodoptera exigua���е�SeGSTe��������б���,����������SeGSTe���ؽ���Cu2+��Cd2+����,��ø���Ա�����ũҩ��������,�����Ʋ�SeGSTe���ܲ������ڵ�����Ӧ����ɱ����Ľⶾ����;LIAO��[25]������ȫצ����Panonychus citri���е�PcGSTm5���������ԭ�˱����,ȷ�����CDNB��GSH���д�л����,��ͨ��dsRNA���������һ����֤��PcGSTm5������ɱ�����ά���أ�abamectin�����Լ�������;ZHANG��[7]�Էɻ�LmGSTS2��ǰ���о���,�����LmGSTS2�ڲ�ͬ�����µĻ��Լ�����֯�ֲ�,����LmGSTs2���г������Ϲܺ�֬����Ⱦ��������ͽⶾ����֯�б��������,�Ʋ�LmGSTS2���ܲ���ɻ����ں��������ʴ�л��Ϊ��һ���������LmGSTS2��ũҩ�Ĵ�л����,���о���LmGSTS2�ɹ��ڴ˾��б���,ͨ��Ni-NTA�Ͳ�����õ�һ��LmGSTS2����,������ø�����ʷ�Ӧ����,�о���LmGSTS2����������p,p��-DDT��ɱ����Ĵ�л����,��ͨ��RNAi���������������ѧ�ⶨ,��һ���ڷɻ�������֤��LmGSTS2���������Ĵ�л�ⶾ���ԡ���IPTG�յ���,LmGSTS2�ɹ����ͨ��Ni-NTA�Ͳ���������õ�һ��Ŀ�������������ڲ�ͬ�����µĻ��Խ��м�ⷢ��,LmGSTS2��pH=7,27������ʱ������ߡ��ɻȵ���ҺpHΪ7.0����,���������¶�Ϊ28��30��,����������Ϊ����ø���������ṩ������������QIN��[20]��LmGSTs5��øѧ�����о�����,�����ʷ�ӦpHΪ6��9,�¶�Ϊ20��40��;��˧��[26]����Сʳ�ij棨Grapholita molesta����GmolGST6��pH=7.5��40��ʱ�������;HIROWATARI��[27]��б��ҹ�꣨Spodoptera litura����SlituGST7��SlituGST20�����ʷ�Ӧ��ΧΪpH 6��8,30��50�档��ͬ���ּ�GST���ʷ�Ӧ������ͬ,������������������������������֯�ֲ���ء�
Ŀǰɱ�����л��ⷽʽ��Ҫ��Һ�ࡢ���༰ɫ��-�������ü���[28,29,30,31],���о���������LmGSTS2����������ֱ�����������p,p��-DDT���з���,ͨ������ЧҺ��ɫ�Է�Ӧ����м�⡣�������,LmGSTS2�ɴ�л��������,�����ܴ�лp,p��-DDT����һ��ͨ��RNAi��ĬLmGSTs2��,�ɻȶ������������ж�����15.3%,����LmGSTS2�������������ڷɻ����ڵĴ�л�ⶾ����,�ӷɻ�������֤�������л�Ľ����GST������Դ����Դ�������ʴ�л������Ҫ��Ϊ4�֣���1��GST���Դ�GSH��ɱ����Ľ��,���ɶ��Խ�С�Ŀ���������[32,33]��NAKKA��[34]ͨ��HPLC��ⷢ�ֳ�â�ȣ�Palmer amaranth���е�GST�ɴ�GSH�밢���ؽ�����������,�Ӷ��Գ��ݼ����ֳ�����;��2��GSH��Ϊ�������ӿ������Եش�DDTֱ�Ӵ�лΪ��DDE,�Ӷ���ɶ�DDT�Ľⶾ[35,36],WANG��[35]���ָԱ��ǰ��ã�Anopheles gambiae���е�agGSTe2����DDT��л����,agGSTe2���GSH���DDT�����˹��������лΪDDE�Ӷ���������;��3��GST���Լ�����ɱ����������������Ӧ�����������ж���������,������ֳ���������ø����[37]��LUMJUAN��[38]���ְ������ã�Aedes aegypti��������GSTE2-2ø���˲���ɱ����ⶾ����֮��,���ɲ���֬�ʹ������ղ���Ĵ�л,���й�������ø����;��4��GST��ͨ����ϲ�����ɱ���,�Ӷ�ʹ���������ҩ��[11,18]�����о���,LmGSTS2���������״�л������δ����������,������Ŀǰ���õ�ɫ���������ʺ��������״�л����ķ���,�������һ����ɫ�����������Ż������������GONZALEZ��[39]�Ժڸ���Ӭ��Drosophila melanogaster����GSTD2���е��ᾧ���ṹ����,ͨ�����ӶԽ�ģ���������Ȼɱ���������������isothiocyanates,ITC����GSTD2��ά�ṹ�������;LOW��[40]ͨ��������ṹ����Ԥ���˺ڸ���Ӭ��GSTD1��GSH��DDT������λ��,��ͨ����ά�˴Ź���1H��15N�Խ��ǰ������н���,�Ӷ�����GSTD1��GSH��DDT�����÷�ʽ�����,�ڽ����о���,��ͨ��������ṹ����,��һ������LmGSTS2���������Ĵ�л�ⶾ���ơ�
4 ����
��LmGSTS2��������б��﴿��,����CDNBΪ�������ø���ʷ�Ӧ����ΪpH=7,27������,��������ɻ����ڻ�����Ϊһ��,�����ɻ����ڻ���Ϊø���������ṩ����������;ͨ������ЧҺ��ɫ��������LmGSTS2�ɴ�л��������,��һ��RNAi��ɱ�����������������ʾLmGSTs2�����Ĭ��ɻȶ������������ж�����,�����ڷɻ�����LmGSTS2������������һ���Ĵ�л���á��ݴ˷���,LmGSTS2���������Ĵ�л���Կ����Ƿɻȶ��������ײ�����ҩ�Ե�ԭ��֮һ���о������Ϊ�ɻȿ��������ͺ���ʩ��ɱ����ṩ��Ҫ���ݡ��ο����� ԭ��˳��
������ȵ���
������������
�����ڿ�Ӱ������
[��������: 1]
[��������: 1]
DOI:10.1007/s00436-015-4898-9URL [��������: 1]
[��������: 1]
[��������: 2]
[��������: 2]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 4]
[D].
[��������: 1]
[D].
[��������: 1]
[D].
[��������: 1]
[D].
[��������: 1]
[��������: 1]
[��������: 1]
DOI:10.1111/imb.2005.14.issue-1URL [��������: 2]
[��������: 1]
[��������: 1]
Magsci [��������: 1]

������<em>S</em>-ת��ø��glutathione <em>S-</em>transferase, GST����һ��㷺�ֲ��Ķ�ܳ�����øϵ�� ����Omega����GST���������ڵ�����Ҫ�������ܡ�Ϊ̽�ַɻ�<em>Locusta migratoria</em> Omega����GST���ܣ� ����RT-PCR������¡�õ�1���ɻȹ�����S-ת��øOmega�������ȫ��cDNA�� ����ΪLmGSTo1 ��GenBank��¼�ţ� JQ750592�����û����Ķ���738 bp�� ����245�������ᡣ��ø����N-�˺�C-��2���ṹ�� N-�˽ṹ����5��β-�۵���3��α������ɣ� ����4��GSH���λ�㣻 C-�˽ṹ����8��α������ɣ� ����5��������λ�㡣Real-time PCR�������, <em>LmGSTo1</em>�ڷɻȲ�ͬ���ھ��б�� ��θä�Һ��г��������ϵͣ� ��ǰ�������Ϲܡ������֬����������ϸ�; ������������ɵ���<em>LmGSTo1</em>����ˮƽ�����½�����Щ���Ϊ��һ���о�<em>LmGSTo1</em>�������ṩ�����ݡ�
Magsci [��������: 1]
������<em>S</em>-ת��ø��glutathione <em>S-</em>transferase, GST����һ��㷺�ֲ��Ķ�ܳ�����øϵ�� ����Omega����GST���������ڵ�����Ҫ�������ܡ�Ϊ̽�ַɻ�<em>Locusta migratoria</em> Omega����GST���ܣ� ����RT-PCR������¡�õ�1���ɻȹ�����S-ת��øOmega�������ȫ��cDNA�� ����ΪLmGSTo1 ��GenBank��¼�ţ� JQ750592�����û����Ķ���738 bp�� ����245�������ᡣ��ø����N-�˺�C-��2���ṹ�� N-�˽ṹ����5��β-�۵���3��α������ɣ� ����4��GSH���λ�㣻 C-�˽ṹ����8��α������ɣ� ����5��������λ�㡣Real-time PCR�������, <em>LmGSTo1</em>�ڷɻȲ�ͬ���ھ��б�� ��θä�Һ��г��������ϵͣ� ��ǰ�������Ϲܡ������֬����������ϸ�; ������������ɵ���<em>LmGSTo1</em>����ˮƽ�����½�����Щ���Ϊ��һ���о�<em>LmGSTo1</em>�������ṩ�����ݡ�
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 2]
[��������: 1]
[��������: 2]
[��������: 1]
[D].
[��������: 1]
[D].
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
Magsci [��������: 1]
ͨ������ʵ��, ��������V(�״�)��V��(ˮ)=7��3Ϊ������, ��Ѳⶨ����Ϊ220 nm�ĸ�ЧҺ��ɫ��ͬʱ�ⶨ����������������ķ���. �������Է�Χ: ��������0.003 66~1.83 ��mol/L, ��������0.121~60.3 ��mol/L, �������ϵ����r����0.9994, ��Ա�ƫ��С��10%. ��������ֱ�Ϊ��������0.003 66 ��mol/L, ��������0.121 ��mol/L.
Magsci [��������: 1]
ͨ������ʵ��, ��������V(�״�)��V��(ˮ)=7��3Ϊ������, ��Ѳⶨ����Ϊ220 nm�ĸ�ЧҺ��ɫ��ͬʱ�ⶨ����������������ķ���. �������Է�Χ: ��������0.003 66~1.83 ��mol/L, ��������0.121~60.3 ��mol/L, �������ϵ����r����0.9994, ��Ա�ƫ��С��10%. ��������ֱ�Ϊ��������0.003 66 ��mol/L, ��������0.121 ��mol/L.
[��������: 1]
DOI:10.3390/molecules21121652URL [��������: 1]
DOI:10.1007/s00216-018-1167-7 [��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 2]
[��������: 1]
[��������: 1]
DOI:10.1016/j.ibmb.2005.03.008URL [��������: 1]
[��������: 1]
[��������: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}