 ,, 孟凡钢, 于德彬, 邱强, 张鸣浩, 饶德民, 丛博韬, 张伟,*, 闫晓艳,*吉林省农业科学院大豆研究所 / 大豆国家工程研究中心, 吉林长春 130033
,, 孟凡钢, 于德彬, 邱强, 张鸣浩, 饶德民, 丛博韬, 张伟,*, 闫晓艳,*吉林省农业科学院大豆研究所 / 大豆国家工程研究中心, 吉林长春 130033Response of agronomic traits and P/Fe utilization efficiency to P application with different P efficiency in soybean
ZHAO Jing,, MENG Fan-Gang, YU De-Bin, QIU Qiang, ZHANG Ming-Hao, RAO De-Min, CONG Bo-Tao, ZHANG Wei,*, YAN Xiao-Yan,*National Engineering Research Center of Soybean / Soybean Research Institute, Jilin Academy of Agricultural Sciences, Changchun 130033, Jilin, China通讯作者: * 张伟, E-mail:zw.0431@163.com;闫晓艳, E-mail:yanxy8548@126.com
收稿日期:2020-09-18接受日期:2021-01-21网络出版日期:2021-03-01
| 基金资助: |
Corresponding authors: * E-mail:zw.0431@163.com;E-mail:yanxy8548@126.com
Received:2020-09-18Accepted:2021-01-21Online:2021-03-01
| Fund supported: |
作者简介 About authors
E-mail:zhao114434260@163.com

摘要
土壤中磷含量与植物铁的吸收密切相关, 为研究供铁充足条件下不同P∶Fe配比对不同磷效率基因型大豆农艺性状和磷/铁利用率的影响, 本文以前期筛选到的磷高效和磷低效大豆品种为试验材料, 采用沙培方式和裂区设计研究不同P∶Fe比对大豆生物学性状的影响及基因型差异, 并利用因子得分综合评价磷高效和磷低效基因型对不同P∶Fe处理的响应, 以解析农艺性状与大豆体内磷/铁利用率的关系, 从而为磷、铁肥合理施用提供理论依据。结果表明: (1) R5期, 磷高效基因型在各处理下的株高、茎粗、根干重和地上部干重增长相对较快, 且均大于磷低效基因型; 磷高效和磷低效基因型在P∶Fe比为100∶100处理下的R5期单株根干重较低, 而百粒重较大。此外, 磷高效和磷低效基因型的籽粒磷利用率在P∶Fe比为1000∶100时降至最低。(2) 典型相关分析表明, 磷高效基因型R5期的茎粗与叶片铁利用率呈正相关关系, 而磷低效基因型R5期的地上部干重与叶片磷利用率呈负相关关系。(3) R8期单株地上部干重和R3期叶片磷利用率的增加有助于磷高效基因型单株粒重的增加, 而R3期单株根重的增加反而会导致磷高效基因型单株粒重的下降。R3期株高、R3期和R8期地上部干重的增加都有助于磷低效基因型单株粒重的增加, 而R3期、R5期和R8期的茎粗以及R5期叶片铁利用率的增加反而导致磷低效基因型单株粒重下降。而且, R8期单株地上部干重对磷高效和磷低效基因型的直接正向贡献均最大。(4) 利用因子得分综合评价发现, P∶Fe≤100∶100时, 磷高效和磷低效基因型在P∶Fe比为100∶100处理下的综合表现最好; 当P∶Fe≥500∶100时, 磷高效和磷低效基因型在P∶Fe比为1000∶100处理下的综合表现最好。综上, 鼓粒初期可以作为筛选不同磷效率基因型的一个重要时期。在铁供应充足情况下, 应考虑到土壤磷素累积和植酸对磷素效率影响问题, 无论是磷高效还是磷低效基因型施P∶Fe比达到1∶1时整体表现最好。
关键词:
Abstract
The phosphorus content in soil is closely related to the iron absorption in plant. In this study, 15 agronomic traits were analyzed by principal component analysis in sand culture and split blot design, with P-efficient and P-inefficient soybean varieties screened in the early stage as the experimental materials. To analyze the relationship between agronomic traits and P/Fe utilization efficiency, and to provide the theoretical basis for the rational application of P and Fe fertilizer in soybean, the effects of different P:Fe ratios on biological traits and genotypic differences were studied in response to P-efficient and P-inefficient genotypes under different P:Fe treatments by factor scores in soybean. The results were as follows: (1) At R5 stage, plant height, stem diameter, root dry weight, and shoot dry weight of P-efficient genotypes were increased relatively rapidly under each treatment, and all of them were higher than those of P-inefficient genotypes. When P:Fe ratio was 100:100, root dry weight per plant at R5 stage was lower, while 100-seed weight was higher. In addition, when P:Fe ratio was 100:100, P utilization efficiency of the two soybean genotypes were the lowest. (2) Canonical correlation analysis revealed that stem diameter at R5 stage of P-efficient genotypes was positively correlated with Fe utilization efficiency in leaves, while shoot dry weight of P-inefficient genotypes was negatively correlated with P utilization efficiency in leaves. (3) The increase of P utilization efficiency of leaves at R3 stage and shoot dry weight at R8 stage were beneficial to the increase of seed weight per plant of P-efficient genotypes, while the increase of P-efficient genotypes at R3 stage would lead to the decrease of seed weight per plant of P-efficient genotypes. The increase of plant height at R3 stage, shoot dry weight at R3 and R8 stages contributed to the increase of seed weight per plant of P-inefficient genotypes, while the increase of stem diameter at R3, R5, and R8 stages, and Fe utilization efficiency of leaves at R5 stage resulted in the decrease of seed weight per plant of P-inefficient genotypes. Furthermore, shoot dry weight at R8 stage had the largest direct positive contribution to both P-efficient and P-inefficient genotypes. (4) Comprehensive evaluation by factor score showed that when P:Fe ≤ 100:100, the comprehensive performance of P-efficient genotypes and P- inefficient genotypes were the best when P:Fe ratio was 100:100. When P:Fe ≥ 500:100, the comprehensive performance of P-efficient genotypes and P-inefficient genotypes were the best when P:Fe ratio was 1000:100. In conclusion, the early stage of seed filling can be an important stage for screening soybean genotypes with different P efficiency. P:Fe ratio at 1:1 was better for both P-efficient and P-inefficient genotypes under sufficient Fe supply, considering the accumulation of phosphate fertilizer in soil and effect of phytates in seed on P efficiency.
Keywords:
PDF (585KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
赵婧, 孟凡钢, 于德彬, 邱强, 张鸣浩, 饶德民, 丛博韬, 张伟, 闫晓艳. 不同磷效率大豆农艺性状与磷/铁利用率对磷素的响应. 作物学报[J], 2021, 47(9): 1824-1833 DOI:10.3724/SP.J.1006.2021.04212
ZHAO Jing, MENG Fan-Gang, YU De-Bin, QIU Qiang, ZHANG Ming-Hao, RAO De-Min, CONG Bo-Tao, ZHANG Wei, YAN Xiao-Yan.
磷(P)是作物必需的营养元素之一[1], 但土壤中能被作物吸收利用的有效磷仅占土壤全磷的1% [2], 土壤有效磷不足, 已成为限制作物产量和品质提高的主要因素之一[3,4]。当供磷水平发生变化时, 植物会在光合作用、活性氧代谢、有机酸代谢等过程产生一系列生理生化方面的适应性变化, 但目前关于磷缺乏后, 植物的响应机制尚不明确, 如磷酸盐转运蛋白与植物低磷胁迫的关系等问题[5]。
土壤中磷含量与植物铁吸收密切相关, 尽管地壳中铁的丰度很高, 平均为3.2%, 但土壤溶液pH值、磷含量以及铁的存在状态(Fe3+)等因素影响铁的有效性, 进而影响植物对铁的吸收[6]。Sánchez-Rodríguez等[7,8]发现, 高水平磷加重鹰嘴豆、花生、羽扇豆和高粱的铁黄化症状。通过同位素示踪和 X-射线能谱测定等技术证明, 磷水平能够改变铁在植物体内的分布[9]。Ward等[10]发现, 缺磷情况下抑制主根伸长的直接因素是铁毒害效应。原因是只要将培养基中铁含量从100 µmol L -1降低至10 µmol L -1时, 拟南芥主根伸长就不受缺磷抑制, 而在不改变磷浓度条件下, 在培养基中加入过量的铁能够得到类似缺磷的拟南芥短根表型, 进一步证明了缺磷抑制拟南芥主根伸长是由于高铁毒害造成的。
一些分子试验研究证明, 磷对于铁相关基因的调控是通过影响铁的有效性而间接引起的。如低磷条件下, 植物体内与铁营养平衡和储存相关基因(FERRITIN1, AtFER1)表达量提高[11,12]。Bournier等[13]研究表明, 磷饥饿响应基因PHR1能够通过不完全回文序列与铁蛋白Ferritin1基因启动子结合, 这是第一个通过分子关系证明磷铁之间存在动态平衡的报道。大豆体内存在复杂的磷铁互作关系。王贤等[14]研究发现, 铁素营养对高铁品种铁积累影响程度大, 而磷素营养对低铁品种铁积累影响程度大。在正常铁素营养水平下, 降低磷素营养水平, 能在提高种子铁含量的同时降低植酸含量, 从而提高铁/植酸的比值。此外, 由于磷素营养水平对大豆根、荚和种子中铁含量影响较大, 表明铁从根向荚及种子运输过程中可能还存在磷素营养调节上的差异。赵婧等[15]研究表明, 高磷并不能使铁低效品种地上部干物质积累量提高, 在高磷条件下, 铁高效品种根系干物质积累能力强于铁低效品种。
综上所述, 磷铁2个营养元素间存在着拮抗效应。而且磷是不可再生资源, 50~400年内将会被耗尽[16,17]。探索植物体内磷铁平衡规律已成为植物营养研究领域备受关注的热点。因此, 本研究以不同磷效率大豆品种为试验材料, 研究不同磷/铁配比对大豆农艺性状和磷、铁利用率的影响, 同时为避免主观因素的干扰, 采取了主成分分析进行综合评价, 以期为土壤中磷铁肥的合理施用提供理论证据。
1 材料与方法
1.1 试验材料
以课题组近年筛选的6个磷高效品种(长农15、吉育69、九农36、吉农23、吉育95和抗线6号)和6个磷低效品种(合丰25、吉农18、欧科豆25、九农27、吉农21和绥农22)为试验材料。1.2 植株培养
选择大小均匀、健康饱满的种子, 播种于长312 cm×宽117 cm×高29 cm装满沙子的矩形种植箱中。每箱种植6个基因型, 每个基因型种植2行, 行距24 cm, 行长117 cm, 株距10 cm。在第1个三出复叶(V1)期之前, 用自来水浇灌植物。从V1期进行处理, 浇灌相应处理液, 除KH2PO4外, 营养液为改良的Hoagland溶液, 其组成为(mmol L-1): 2.0 Ca(NO3)2、0.75 K2SO4、0.65 MgSO4、0.1 Fe-EDTA、10×10-3 H3BO3、1×10-3 MnSO4、0.5×10-3 CuSO4、0.5×10-3 ZnSO4、0.05×10-3 (NH4)6Mo7O24。溶液pH按要求用NaOH或HCl调节至5.8±0.1。种植箱每隔1 d浇水, 并使用土壤测量仪(TDR 350)测量土壤水分, 田间持水量保持在60%。供试沙子的全磷含量为0.12 g kg-1、全铁含量为0.007%。1.3 试验设计
试验于2017—2018年在吉林省农业科学院公主岭院区进行。试验采用裂区设计, 3次重复, 1个种植箱为1次重复。主区为磷/铁比处理, 设4个水平磷浓度, 使P:Fe配比如表1所示, 裂区为基因型, 设磷高效基因型和磷低效基因型。Table 1
表1
表1主区试验设计
Table 1
| 处理 Treatment | 磷浓度 P concentration (μmol L-1) | 磷:铁 P:Fe |
|---|---|---|
| CK | 0 | 0:100 |
| P1 | 100 | 100:100 |
| P2 | 500 | 500:100 |
| P3 | 1000 | 1000:100 |
新窗口打开|下载CSV
1.4 测定项目与方法
1.4.1 农艺性状 于2017年始荚期(R3)、鼓粒期(R5)和成熟期(R8)取样, 从子叶节处剪断大豆植株, 将根连同附着于根上的沙子用水浸泡, 轻轻振荡, 然后用自来水冲洗干净, 105℃杀青1 h, 然后85℃烘干, 测定其干重。每处理连续取有代表性的4株, 3次重复, 于R3和R5期测定株高、茎粗、单株根系干重、单株地上部干重; 于R8期测定株高、茎粗、单株根系干重、单株地上部干重、单株粒重、百粒重。1.4.2 磷、铁利用率 于2018年始荚期(R3)和鼓粒初期(R5)取各处理叶片, 以及成熟期(R8)各处理籽粒, 采用电感耦合等离子体发射光谱法(ICP-OES)测定样品中的磷、铁浓度, 3次重复。磷利用率用单位磷素(P; mg)所产生的干物质量(dry weight, DW; g)表示; 铁利用率用单位铁素(P; mg)所产生的干物质量(DW; g)表示。
1.5 计算方法
式中, Ci为综合因子得分, CE为综合评价值, n为主成分个数。
式中, ACE为综合评价均值, n为主成分个数。
1.6 数据分析
1.6.1 典型相关性分析 将R3期株高(X1)、R3期茎粗(X2)、R3期单株根干重(X3)、R3期单株地上部干重(X4)、R5期株高(X5)、R5期茎粗(X6)、R5期单株根干重(X7)、R5期单株地上部干重(X8)、R8期株高(X9)、R8期茎粗(X10)、R8期单株根干重(X11)、R8期单株地上部干重(X12)、R8期百粒重(X13)归为一组作为农艺性状指标; 将R3期叶片铁利用率(Y1)、R5期叶片铁利用率(Y2)、R8期籽粒铁利用率(Y3)、R3期叶片磷利用率(Y4)、R5期叶片磷利用率(Y5)和R8期籽粒磷利用率(Y6)归为一组作为磷、铁利用率指标, 做典型相关分析。1.6.2 建立逐步回归方程 以X1~X13和Y1~Y6共19个性状为自变量, 分别以磷高效基因型和磷低效基因型单株粒重为因变量, 建立逐步回归方程。
1.6.3 通径分析 以X1~X10和Y1~Y9共19个性状为自变量, 分别以磷高效基因型和磷低效基因型单株粒重为因变量进行通径分析。
1.6.4 数据处理 采用DPS v18.0软件分析数据。
2 结果与分析
2.1 P:Fe比例对大豆农艺性状的影响及基因型差异
2.1.1 株高 从图1-A可以看出, 磷高效基因型在R3期株高(X1)随着P:Fe比的增大而逐渐增高, 但到了R5期和R8期这种趋势逐渐减弱, 从R5期到R8期之间, 磷高效基因型的株高没有较大增长。磷低效基因型的R3期株高随着P:Fe比的增大而呈先降后增的趋势, 同样, 到了R5期和R8期这种趋势也逐渐减弱, 但从R5期到R8期之间, 磷低效基因型的株高增长较大。图1
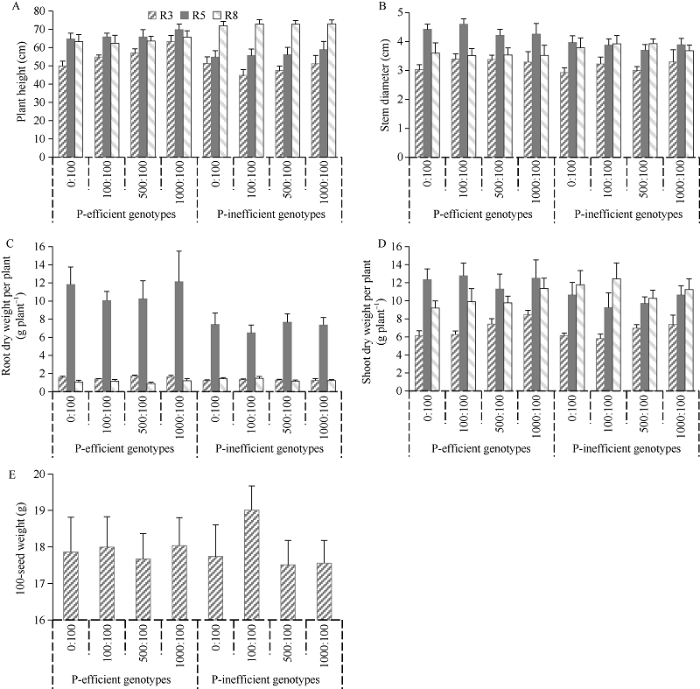 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1磷:铁比对不同大豆基因型农艺性状的影响
R3: 始荚期; R5: 鼓粒期; R8: 成熟期。
Fig. 1Effects of P:Fe ratios on agronomic traits of different soybean genotypes
R3: initial podding stage; R5: seed filling stage; R8: maturity stage.
2.1.2 茎粗 磷高效和磷低效基因型在P:Fe比为0:100时R3期茎粗(X2)最小, 到了R5期明显增粗, 尤其是磷高效基因型(图1-B)。但到了R8期, 磷高效基因型的茎粗有所下降, 而磷低效基因型的茎粗与R5期相比没有较大变化。
2.1.3 根干重 从3个时期的单株根重来看, 磷高效和磷低效基因型的根系生长在R5期达到峰值, 且磷高效基因型要大于磷低效基因型(图1-C)。磷高效和磷低效基因型在P:Fe比为100:100时的单株根干重均小于其余处理。
2.1.4 地上部干重 磷高效和磷低效基因型在R3期的地上部干重(X4)随着P:Fe比的增大而逐渐增高, 但到了R5期和R8期这种趋势逐渐减弱(图1-D)。到了R5期, 磷高效基因型在各处理下的地上部干重均大于磷低效基因型。
2.1.5 百粒重 磷高效基因型各处理间的百粒重没有明显差异, 而磷低效基因型在P:Fe比为100:100时百粒重要大于其余处理(图1-E)。
2.2 P:Fe比例对大豆磷、铁利用率的影响及基因型差异
2.2.1 铁利用率 从3个生育期来看, R8期时籽粒铁利用率(Y3)最高(图2-A)。磷高效和磷低效基因型在R5和R8期的铁利用率整体上都随着P:Fe比的增大而呈先增后降的趋势。磷高效基因型R8期籽粒铁利用率在P:Fe比为100:100时最高, 磷低效基因型则在P:Fe比为500:100时最高。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2磷:铁比对不同大豆基因型磷/铁利用率的影响
R3: 始荚期; R5: 鼓粒期; R8: 成熟期。
Fig. 2Effects of P:Fe ratios on P/Fe utilization efficiency of different soybean genotypes
R3: initial podding stage; R5: seed filling stage; R8: maturity stage.
2.2.2 磷利用率 从不同生育时期来看(图2-B), R5期叶片磷利用率最高, R8期籽粒磷利用率(Y6)最低。磷高效基因型的R3期叶片磷利用率(Y4)在不同P:Fe比处理间差异不明显, 但随着P:Fe比的增大, 有微弱的增加趋势, 而磷低效基因型与之相反。到了R5期, 叶片磷利用率明显上升, 随着P:Fe比的增大, 2种基因型总体上都呈先增后减的趋势。到了R8期, 2种基因型的籽粒磷利用率在P:Fe比为1000:100时降到最低。
2.3 农艺性状与磷、铁利用率的典型相关性分析
由表2可知, 通过对农艺性状与磷、铁利用率进行典型相关分析, 磷高效和磷低效基因型各得出1组典型相关变量, 达显著或极显著水平。从磷高效基因型典型变量系数可以看出, 这组主要反应了X6 (R5期单株地上部干重)和Y2 (R5期叶片铁利用率)的正相关关系, 说明磷高效基因型鼓粒初期的茎粗与叶片铁利用率呈正相关关系, 即鼓粒初期叶片铁利用率有助于增加茎粗。从磷低效基因型典型变量系数可以看出, 这组主要反应了X8 (R5期茎粗)和Y5 (R5期叶片磷利用率)的负相关关系, 说明磷低效基因型鼓粒初期的地上部干重与叶片磷利用率呈负相关关系, 也就是说鼓粒初期叶片磷利用率的提升不利于地上部干重的累积。Table 2
表2
表2不同磷:铁比条件下大豆光合叶绿素荧光参数与磷、铁性状的典型相关性分析
Table 2
| 基因型 Genotype | 典型相关系数 Canonical correlation coefficient | P值P-value | 典型向量 Canonical variables |
|---|---|---|---|
| 磷高效基因型 P-efficient genotypes | 0.92 | 0.0145 | UE = - 0.1774X1 + 0.3527X2 - 0.5376X3 + 0.6918X4 - 0.8259X5 + 1.6345X6 + 0.3873X7 - 0.5418X8 - 0.3599X9 + 0.0221X10 - 0.7607X11 + 0.8133X12 - 0.0339X13 |
| VE = 0.4051Y1 + 0.6116Y2 - 0.4141Y3 + 0.2499Y4 - 0.3933Y5 + 0.3567Y6 | |||
| 磷低效基因型 P-inefficient genotypes | 0.98 | 0.0059 | UIE = 0.7518X1 - 0.2609X2 - 0.2263X3 - 0.4150X4 - 0.5712X5 + 0.4853X6 - 0.4017X7 + 0.8477X8 + 0.3437X9 + 0.2264X10 - 0.1210X11 - 0.2134X12 + 0.0590X13 |
| VIE = 0.5590Y1 - 0.6724Y2 + 0.0917Y3 - 0.6286Y4 - 0.7682Y5 + 0.6182Y6 |
新窗口打开|下载CSV
2.4 逐步回归方程的建立
由表3可知, 磷高效和磷低效基因型的逐步回归方程均具有显著意义。从磷高效基因型的逐步回归方程可以看出, WE与X12 (R8期单株地上部干重)和Y4 (R3期叶片磷利用率)呈正相关, 与X3 (R3期单株根重)呈负相关。这表示, R8期单株地上部干重和R3期叶片磷利用率的增加有助于磷高效基因型单株粒重的增加, 而R3期单株根重的增加反而会导致磷高效基因型单株粒重的下降。Table 3
表3
表3单株粒重的逐步回归方程
Table 3
| 基因型 Genotype | 回归方程 Regression equation | 决定系数 R2 | P值 P-value | 剩余标准差 Residual standard deviation | 杜宾-沃森检验 Durbin-Watson test |
|---|---|---|---|---|---|
| 磷高效基因型 P-efficient genotypes | WE= 1.34 - 0.60X3 + 0.25X12 + 1.02Y4 | 0.89 | 1.00E-07 | 0.0118 | 2.20 |
| 磷低效基因型 P-inefficient genotypes | WIE = 6.84 + 0.027X1- 0.67X2+ 0.22X4 - 0.87X6 - 0.74X10 + 0.24X12- 0.22Y2 | 0.87 | 5.41E-06 | 0.0196 | 1.62 |
新窗口打开|下载CSV
磷低效基因型WIE与X1 (R3期株高)、X4 (R3期单株地上部干重)和X12 (R8期单株地上部干重)呈正相关, 与X2 (R3期茎粗)、X6 (R5期茎粗)、X10 (R8期茎粗)和Y2 (R5期叶片铁利用率)呈负相关。这表明, R3期株高、R3期和R8期的地上部干重的增加都有助于磷低效基因型单株粒重的增加, 而R3、R5和R8期的茎粗以及R5期叶片铁利用率的增加反而导致磷低效基因型单株粒重下降。
2.5 通径分析
磷高效基因型单株粒重与各性状间的通径分析结果(表4)表明, X3(R3期单株根重)对磷高效基因型单株粒重具有负向作用, X12 (R8期单株地上部干重)和Y4 (R3期叶片磷利用率)对单株粒重为正向作用, 其中X12的直接正向效应最大, 达0.9028。虽然X3对单株粒重具有负向直接作用, 但X12却可以通过X3的间接正向作用促进单株粒重的提高。而X3和X12的间接负向作用也减弱了Y4对单株粒重的正向直接贡献。Table 4
表4
表4磷高效基因型单株粒重与各性状间的通径分析
Table 4
| 变量 Variable | 直接通径系数 Direct path coefficient | →X3 | →X12 | →Y4 |
|---|---|---|---|---|
| X3 | -0.2481 | -0.0755 | 0.0791 | |
| X12 | 0.9028 | 0.0208 | -0.0083 | |
| Y4 | 0.1610 | -0.1220 | -0.0465 |
新窗口打开|下载CSV
磷低效基因型单株粒重与各性状间的通径分析结果(表5)表明, 对磷低效基因型单株粒重直接效应为正的性状有3个, 大小依次为X12 (R8期单株地上部干重) > X4 (R3期单株地上部干重) > X1 (R3期株高); 而对磷低效基因型单株粒重直接效应为负的性状有4个, 分别为X2 (R3期茎粗)、X6(R5期茎粗)、X10 (R8期茎粗)和Y2 (R5期叶片铁利用率), 其中Y2的负向作用最小。而且, Y2在X1、X2、X4、X6、X10和X12中的间接作用都是正向的。X12通过X1、X2、X4、X6和Y2的正向间接作用, 促进磷高效基因型单株粒重的增加。X1、X2、X4、X6、X12和Y2正向间接作用减弱了X10对单株粒重负向直接贡献。
Table 5
表5
表5磷低效基因型单株粒重与各性状间的通径分析
Table 5
| 变量 Variable | 直接通径系数 Direct path coefficient | →X1 | →X2 | →X4 | →X6 | →X10 | →X12 | →Y2 |
|---|---|---|---|---|---|---|---|---|
| X1 | 0.2277 | 0.1770 | 0.2169 | -0.2246 | -0.0942 | 0.1118 | 0.0425 | |
| X2 | -0.4223 | 0.0954 | 0.1693 | -0.1133 | 0.0647 | -0.1678 | 0.0080 | |
| X4 | 0.3424 | 0.1442 | -0.2088 | -0.1718 | -0.1147 | 0.1520 | 0.0236 | |
| X6 | -0.4367 | 0.1171 | -0.1096 | 0.1347 | 0.0029 | -0.0101 | 0.0182 | |
| X10 | -0.4472 | 0.0479 | 0.0610 | 0.0878 | 0.0029 | 0.4820 | 0.0480 | |
| X12 | 0.7848 | 0.0324 | 0.0903 | 0.0663 | 0.0056 | -0.2746 | 0.0470 | |
| Y2 | -0.1587 | -0.0610 | 0.0214 | -0.0509 | 0.0501 | 0.1354 | -0.2326 |
新窗口打开|下载CSV
2.6 综合评价
以19个农艺性状为基础, 分别对磷高效和磷低效基因型在不同P:Fe比水平下的表现进行主成分分析, 计算主成分的综合值进行综合评价(表6和表7)。P:Fe比为0:100 (CK)处理导致磷高效和磷低效基因型的综合评价值最低, 分别为-0.265和-0.096。当P:Fe≤100:100时, 磷高效和磷低效基因型在P:Fe比为100:100 (P1)处理下的综合表现最好; 当P:Fe≥100:100时, 磷高效和磷低效基因型在P:Fe比为1000:100 (P3)处理下的综合表现最好。Table 6
表6
表6磷高效基因型在各处理下的综合评价
Table 6
| 处理Treatment | 基因型 Genotype | C1 | C2 | C3 | C4 | C5 | C6 | CE | ACE |
|---|---|---|---|---|---|---|---|---|---|
| CK | 长农15 Changnong 15 | -0.543 | 0.283 | 0.114 | -0.045 | -0.022 | 0.042 | -0.171 | -0.265 |
| 吉育69 Jiyu 69 | -0.923 | 0.392 | -0.073 | -0.184 | 0.023 | -0.154 | -0.919 | ||
| 九农36 Jiunong 36 | 1.191 | -0.432 | -0.188 | -0.218 | 0.081 | -0.122 | 0.312 | ||
| 吉农23 Jinong 23 | -1.036 | -0.340 | 0.130 | -0.112 | -0.097 | 0.056 | -1.399 | ||
| 吉育95 Jiyu 95 | -0.311 | 0.135 | 0.013 | -0.058 | -0.004 | -0.046 | -0.270 | ||
| 抗线6号Kangxian 6 | 0.736 | -0.013 | 0.410 | -0.144 | -0.069 | -0.062 | 0.857 | ||
| P1 | 长农15 Changnong 15 | -0.197 | 0.275 | 0.399 | 0.241 | 0.057 | -0.009 | 0.765 | 0.127 |
| 吉育69 Jiyu 69 | -1.117 | 0.379 | 0.077 | 0.071 | 0.088 | -0.036 | -0.538 | ||
| 九农36 Jiunong 36 | 1.149 | -0.463 | 0.318 | 0.108 | 0.035 | 0.065 | 1.212 | ||
| 吉农23 Jinong 23 | -0.742 | -0.313 | 0.232 | -0.106 | 0.034 | 0.057 | -0.838 | ||
| 吉育95 Jiyu 95 | 0.008 | -0.258 | 0.315 | 0.032 | -0.045 | -0.006 | 0.048 | ||
| 抗线6 Kangxian 6 | -0.057 | -0.191 | 0.307 | -0.020 | 0.023 | 0.053 | 0.114 | ||
| P2 | 长农15 Changnong 15 | -0.190 | 0.517 | 0.001 | 0.012 | 0.025 | 0.055 | 0.421 | 0.019 |
| 吉育69 Jiyu 69 | -0.538 | 0.068 | 0.244 | -0.041 | -0.080 | 0.035 | -0.312 | ||
| 九农36 Jiunong 36 | 0.266 | -0.396 | -0.346 | -0.091 | 0.175 | 0.003 | -0.390 | ||
| 吉农23 Jinong 23 | -0.732 | -0.199 | -0.340 | -0.187 | 0.076 | 0.128 | -1.254 | ||
| 吉育95 Jiyu 95 | 0.386 | 0.162 | -0.306 | 0.112 | -0.025 | 0.037 | 0.365 | ||
| 抗线6 Kangxian 6 | 0.625 | 0.047 | 0.329 | 0.195 | 0.147 | -0.058 | 1.285 | ||
| P3 | 长农15 Changnong 15 | -0.942 | 0.071 | -0.446 | 0.237 | 0.024 | -0.007 | -1.064 | 0.119 |
| 吉育69 Jiyu 69 | -0.930 | 0.194 | -0.095 | -0.039 | -0.167 | -0.060 | -1.098 | ||
| 九农36 Jiunong 36 | 1.139 | -0.671 | -0.142 | 0.053 | -0.143 | -0.035 | 0.202 | ||
| 吉农23 Jinong 23 | -0.363 | -0.156 | -0.461 | 0.016 | -0.023 | 0.015 | -0.973 | ||
| 吉育95 Jiyu 95 | 0.606 | -0.086 | -0.341 | 0.329 | -0.082 | -0.025 | 0.402 | ||
| 抗线6号Kangxian 6 | 2.517 | 0.996 | -0.151 | -0.161 | -0.032 | 0.074 | 3.243 |
新窗口打开|下载CSV
Table 7
表7
表7磷低效基因型在各处理下的综合评价
Table 7
| 处理 Treatment | 基因型 Genotype | C1 | C2 | C3 | C4 | C5 | C6 | C7 | CE | ACE |
|---|---|---|---|---|---|---|---|---|---|---|
| CK | 合丰25 Hefeng 25 | 0.040 | -0.624 | 0.120 | -0.105 | -0.031 | -0.045 | 0.017 | -0.628 | -0.096 |
| 吉农18 Jinong 18 | -0.062 | -0.891 | -0.019 | 0.052 | 0.108 | -0.041 | -0.060 | -0.913 | ||
| 欧科豆25 Oukedou 25 | 0.247 | 0.237 | 0.093 | 0.021 | 0.116 | 0.039 | -0.002 | 0.752 | ||
| 九农27 Jiunong 27 | 0.206 | 0.879 | 0.324 | -0.021 | 0.073 | -0.070 | -0.011 | 1.379 | ||
| 吉农21 Jinong 21 | -0.490 | 0.384 | 0.005 | -0.218 | 0.146 | -0.073 | -0.035 | -0.281 | ||
| 绥农22 Suinong 22 | 0.166 | -0.737 | -0.193 | -0.335 | 0.166 | 0.077 | -0.028 | -0.884 | ||
| P1 | 合丰25 Hefeng 25 | -0.927 | -0.445 | 0.229 | 0.224 | 0.006 | 0.022 | -0.003 | -0.895 | 0.106 |
| 吉农18 Jinong 18 | 0.279 | -0.237 | 0.006 | 0.159 | 0.160 | 0.056 | 0.028 | 0.451 | ||
| 欧科豆25 Oukedou 25 | -0.317 | 0.209 | -0.116 | 0.050 | 0.035 | 0.006 | -0.041 | -0.174 | ||
| 九农27 Jiunong 27 | 1.376 | -0.051 | 0.280 | 0.102 | 0.043 | -0.098 | 0.000 | 1.652 | ||
| 吉农21 Jinong 21 | 0.428 | -0.359 | -0.041 | 0.095 | 0.061 | -0.104 | 0.010 | 0.090 | ||
| 绥农22 Suinong 22 | -0.837 | 0.360 | -0.177 | 0.199 | 0.073 | -0.096 | -0.009 | -0.487 | ||
| P2 | 合丰25 Hefeng 25 | -0.395 | -0.254 | -0.006 | 0.175 | -0.121 | -0.005 | -0.001 | -0.607 | -0.075 |
| 吉农18 Jinong 18 | -0.117 | 0.071 | 0.227 | 0.047 | 0.067 | 0.253 | 0.114 | 0.662 | ||
| 处理 Treatment | 基因型 Genotype | C1 | C2 | C3 | C4 | C5 | C6 | C7 | CE | ACE |
| 欧科豆25 Oukedou 25 | -0.304 | 0.333 | 0.034 | 0.018 | -0.006 | -0.063 | 0.031 | 0.043 | ||
| 九农27 Jiunong 27 | 0.932 | -0.035 | 0.054 | -0.186 | -0.100 | -0.047 | 0.026 | 0.646 | ||
| 吉农21 Jinong 21 | -0.543 | 0.424 | -0.067 | -0.165 | 0.060 | -0.021 | -0.030 | -0.342 | ||
| 绥农22 Suinong 22 | -0.551 | 0.031 | -0.318 | -0.048 | -0.062 | -0.081 | 0.179 | -0.850 | ||
| P3 | 合丰25 Hefeng 25 | -0.502 | -0.469 | 0.150 | -0.073 | -0.222 | 0.058 | -0.031 | -1.090 | 0.064 |
| 吉农18 Jinong 18 | 1.439 | 0.200 | -0.542 | 0.172 | -0.037 | 0.075 | -0.022 | 1.285 | ||
| 欧科豆25 Oukedou 25 | -0.272 | 0.333 | -0.059 | 0.040 | -0.109 | 0.095 | -0.163 | -0.137 | ||
| 九农27 Jiunong 27 | 0.907 | 0.057 | 0.124 | -0.055 | -0.231 | -0.046 | 0.004 | 0.761 | ||
| 吉农21 Jinong 21 | 0.156 | 0.585 | 0.002 | -0.091 | -0.043 | 0.138 | 0.025 | 0.772 | ||
| 绥农22 Suinong 22 | -0.860 | 0.001 | -0.110 | -0.057 | -0.155 | -0.029 | 0.006 | -1.204 |
新窗口打开|下载CSV
3 讨论
铁和磷是植物生长所必需的2种元素, 这2种元素在土壤中都很丰富, 但植物对它们的可利用性很差。土壤中铁的低有效性主要是由于高碳酸氢盐浓度、高pH值、重金属等不利的土壤条件[18,19,20,21]。植物磷可用性主要受土壤中磷浓度、供给植物的固态磷量和土壤溶液磷容量3个因素影响[7]。在双子叶植物中, 缺铁或缺磷引起的一些形态和生理反应是相似的, 如根毛的形成、丛根的形成、根际的酸化以及产生和释放有机酸[22]。在整个进化过程中, 植物采取了多种适应性策略来解决铁和磷的营养胁迫[23]。随着耕地营养缺乏的加剧, Fageria等[24]认为, 在21世纪农业中, 高效营养植物将在提高作物产量方面发挥重要作用。在本研究中, 磷高效基因型在R3期株高和地上部干重均随着P:Fe比的增大而逐渐增高, 到了R5期, 磷高效基因型在各处理下的株高、茎粗、根干重和地上部干重增长相对较快, 且均大于磷低效基因型(图1)。此外, 典型相关分析表明, R5期叶片铁利用率有助于增加磷高效基因型茎粗(表2), R5期叶片磷利用率的提升不利于磷低效基因型地上部干重的累积。这些结果表明, 鼓粒初期是筛选不同磷效率基因型的一个重要时期。
缺磷胁迫下, 初生根的生长受到强烈抑制, 而侧根和根毛的发育则得到了大量促进, 最终导致根/冠比的增加。虽然在缺氮条件下也可以观察到类似的反应, 但在缺磷条件下分生组织活性逐渐丧失, 而在缺氮条件下仍保持相对活跃[25]。由于根系是磷的主要进入点, 因此根系结构的改变对于最大限度地有效吸收磷对缺磷植物存活至关重要。在本研究中磷高效和磷低效基因型在P:Fe比为100:100时的R5期单株根干重均较低(图1-C), 这可能是因为P:Fe比为100:100处理并没有引起植株根系的缺磷反应。此外, 磷高效和磷低效基因型在P:Fe比为100:100时百粒重均较大(图1-E)。而且当P:Fe≤100:100时, 磷高效和磷低效基因型在P:Fe比为100:100 (P1)处理下的综合表现最好(表6和表7)。这些结果可能表明P:Fe比为100:100是比较适合植株生长的。通过逐步回归方程和通径分析可以看出, R8期单株地上部干重对磷高效和磷低效基因型的单株粒重直接正向贡献均最大。但性状间并不是孤立存在的, 而是互相协调的。如本研究中, 虽然R3期单株根干重的增加会导致磷高效基因型单株粒重的下降, 但R8期单株地上部干重却可以通过R3期单株根重的间接正向作用促进单株粒重的提高。
提高磷的获取和利用效率对于防止磷资源短缺和减少施磷对环境的影响变得越来越重要[26]。提高磷利用效率, 可以通过对植物中磷的最佳分配和再分配来实现, 使植物最大限度地生长和生物量分配到可收获的植物部分。通过磷的再转移和减少磷向发育中籽粒的分配, 可以改善植物体内磷的分布。本研究中从R3期到R5期, 叶片磷利用率升至峰值, 这种变化将延长和提高光合作用中磷的生产性利用, 并具有营养和环境效益[27]。见图2, 2种基因型的籽粒磷利用率在P:Fe比为1000:100时降至最低。这可能是因为P:Fe比过高导致籽粒磷含量过量积累, 从而使磷利用率大大降低。Rose等[28]研究发现, 籽粒磷含量的变化与水稻籽粒大小和产量的降低均无相关性, 并认为低籽粒磷浓度可能是培育更高效磷高效水稻的一个合适标准, 以减少田间收获的磷输出。而且种子中高达80%的磷都是以植酸形式储存, 但在种子和消化过程中, 它可以使铁和锌等矿物质复杂化, 使它们无法被吸收[29]。因此, 虽然当P:Fe≥500:100时, 磷高效和磷低效基因型在P:Fe比为1000:100 (P3)处理下的综合表现最好, 过量施入磷肥可能引起品质问题和环境问题。有研究认为高磷条件会明显减少大豆植株对Fe吸收, 并降低Fe在植株中的活动性, 从而导致Fe在体内失活[30]。而高磷条件所产生的铁转运受阻, 可能是PO43-作为一种与螯合剂竞争Fe2+的配位体, 在植物体内形成FePO4的沉淀效应所致[31]。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 2]
[本文引用: 1]
PMID [本文引用: 1]

Phosphate (Pi) is an essential element for plant development and metabolism. Due to its low availability and mobility in soils, it is often a limiting nutrient for their growth. This phenomenon is reinforced by the formation of insoluble complexes in the environment with many cations, affecting the solubility of both phosphate and associated ions. This interaction is investigated here for iron, a strong phosphate chelator. Depleting the medium in phosphate clearly resulted in an increase of iron content in Arabidopsis. These modifications triggered molecular responses linked with iron status (transport, homeostasis and accumulation). Interestingly, physiological modifications affecting iron storage were also observed. The accumulation of phosphate/iron complexes in the vacuoles of plants grown in Pi-rich medium disappeared in Pi-depleted medium in favor of accumulation of iron inside the chloroplasts, likely associated with ferritin.
DOIURL [本文引用: 1]
[本文引用: 1]
PMID [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIPMID [本文引用: 1]
To cope with nutrient deficiencies, plants develop both morphological and physiological responses. The regulation of these responses is not totally understood, but some hormones and signaling substances have been implicated. It was suggested several years ago that ethylene participates in the regulation of responses to iron and phosphorous deficiency. More recently, its role has been extended to other deficiencies, such as potassium, sulfur, and others. The role of ethylene in so many deficiencies suggests that, to confer specificity to the different responses, it should act through different transduction pathways and/or in conjunction with other signals. In this update, the data supporting a role for ethylene in the regulation of responses to different nutrient deficiencies will be reviewed. In addition, the results suggesting the action of ethylene through different transduction pathways and its interaction with other hormones and signaling substances will be discussed. © 2015 American Society of Plant Biologists. All Rights Reserved.
[本文引用: 1]
DOIPMID [本文引用: 1]
Phosphorus is an essential macronutrient for plant growth and development. Phosphate (Pi), the major form of phosphorus that plants take up through roots, however, is limited in most soils. To cope with Pi deficiency, plants activate an array of adaptive responses to reprioritize internal Pi use and enhance external Pi acquisition. These responses are modulated by sophisticated regulatory networks through both local and systemic signaling, but the signaling mechanisms are poorly understood. Early studies suggested that the phytohormone ethylene plays a key role in Pi deficiency-induced remodeling of root system architecture. Recently, ethylene was also shown to be involved in the regulation of other signature responses of plants to Pi deficiency. In this article, we review how researchers have used pharmacological and genetic approaches to dissect the roles of ethylene in regulating Pi deficiency-induced developmental and physiological changes. The interactions between ethylene and other signaling molecules, such as sucrose, auxin, and microRNA399, in the control of plant Pi responses are also examined. Finally, we provide a perspective for the future research in this field.
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}