 ,2,3,4,*
,2,3,4,*Effects of irrigating at different growth stages on kernel number of spring maize in the North China Plain
GAO Zhen1,2, LIANG Xiao-Gui2, ZHANG Li2, ZHAO Xue2, DU Xiong1, CUI Yan-Hong1, ZHOU Shun-Li,2,3,4,*通讯作者:
收稿日期:2020-07-24接受日期:2020-12-1网络出版日期:2020-12-29
| 基金资助: |
Received:2020-07-24Accepted:2020-12-1Online:2020-12-29
| Fund supported: |
作者简介 About authors
E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (784KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
高震, 梁效贵, 张莉, 赵雪, 杜雄, 崔彦宏, 周顺利. 不同时期灌溉对华北平原春玉米穗粒数的影响[J]. 作物学报, 2020, 47(7): 1324-1331. doi:10.3724/SP.J.1006.2021.03045
GAO Zhen, LIANG Xiao-Gui, ZHANG Li, ZHAO Xue, DU Xiong, CUI Yan-Hong, ZHOU Shun-Li.
水资源匮乏是限制华北平原农业生产的关键因素[1,2,3,4,5,6,7], 将冬小麦-夏玉米一年两熟制改为春玉米一年一熟制可以大幅降低灌溉用水, 同时保证相对较高的产量水平[1,7]。然而, 春玉米生育期间会遭遇干旱、高温、阴雨寡照等多种非生物逆境胁迫, 严重限制春玉米产量优势的发挥, 其中干旱胁迫是影响春玉米产量形成的重要因子[8]。华北地区降水主要集中在7月至8月份, 且年际间变化较大, 春玉米任何生育时期都可能遭遇干旱胁迫。不同生育阶段干旱胁迫对玉米产量的影响程度不同, 苗期干旱对玉米产量影响较小[9], 适当的干旱胁迫会增强其抗旱性[10]; 而大口期-吐丝期玉米植株快速生长, 且对水分需求量大, 干旱胁迫导致营养生长受阻, 玉米株高和叶面积明显降低, 最终穗粒数和产量显著降低[11]; 吐丝及吐丝后2周是玉米对水分亏缺最敏感时期, 干旱胁迫会造成玉米吐丝延迟和籽粒败育, 大幅降低玉米产量, 且干旱胁迫时间越长, 强度越大, 产量降低幅度越大[12,13]。灌溉是保证玉米产量的重要措施[14], 全生育期灌溉可以获得较高产量, 但会降低水分利用效率[15]; 单次灌溉较雨养条件可以增产3%~35%, 且花期灌溉可以达到全生育期灌溉产量的83%~89%, 水分利用效率提高27%~40%[16]。因此, 在有限水灌溉条件下, 在玉米的关键生育阶段进行灌溉, 对提高其水分高效利用和实现产量稳定至关重要[17]。
玉米产量构成因素为单位面积穗数、穗粒数和粒重。一般认为穗粒数对环境变化最敏感[18], 同一品种粒重变化较小, 在年际间表现较为稳定[17]。在确保足够穗数的基础上, 增加产量取决于穗粒数和穗粒重, 而增加穗粒数是提高穗粒重的关键[19]。干旱胁迫特别是花期干旱胁迫会显著降低春玉米穗粒数[17], 其主要原因是: (1)干旱胁迫会明显抑制花丝生长, 造成开花吐丝间隔期(anthesis silking interval, ASI)延长, 顶部籽粒不能授粉最终败育[20]; (2)籽粒的库活性和碳水化合物供应能力下降, 籽粒发生败育[21]。开花期灌水可以明显降低玉米籽粒败育率, 提高产量和水分利用效率[9]。本团队研究结果同样表明, 尽管春玉米营养生长阶段受到干旱胁迫抑制, 但花期灌水保证了吐丝-授粉阶段的水分供应, 进而降低籽粒败育率, 实现产量和水分利用效率的协同提升[17]。前人围绕不同时期干旱胁迫影响玉米籽粒结实及灌溉效应进行了大量研究[11,12,13,14,15,16,17], 但不同时期灌水对花期土壤水分、叶片光合速率及穗粒数的影响及相互关系仍不明确。本研究利用3年大田试验, 明确在不同降水年型下各生育时期灌水处理对土壤水分变化动态的影响, 阐明花期土壤水分含量与春玉米叶片光合速率及穗粒数的关系, 以期为华北地区春玉米合理灌溉策略制订提供理论支撑。
1 材料与方法
1.1 试验地概况
试验于2014—2016年在中国农业大学吴桥实验站(37°41′N, 116°36′E)进行。试验地为轻壤土, 2 m深度土体平均土壤容重为1.51 g cm-3, 平均田间持水量为27.6%。地表0~40 cm土壤有机质含量为1.17%, 全氮含量为0.95 g kg-1, 有效钾含量104.4 mg kg-1, 有效磷含量29.2 mg kg-1。该地区过去30年春玉米生育期平均降水量为438 mm, 2014—2016年春玉米生长季降雨分别为243、410和584 mm。2014年大口期有60 mm (连续2 d降水量)降水, 该时期灌溉处理被取消, 在灌浆期有2次有效降水保证了籽粒生长, 但降水过低, 将其定为干旱年型; 2015年从V10到吐丝后15 d无有效降雨, 春玉米在籽粒形成的关键时期遭遇明显的干旱胁迫, 而之后有较多降水, 但春玉米较高的空秆率和较低的结实率导致后期降水不能被有效利用, 因此将其定为关键时期干旱年份; 2016年在营养生长早期降水较少, 从V9时期至收获, 有充足降水保证春玉米的水分需求, 但阴雨寡照天气增多(图1), 较平均降雨量多1/3, 将其定为多雨年份。
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图12014-2016年春玉米生育期降水及温度分布
图中箭头指示吐丝期; V6、V12、R1、R6分别为拔节期、大口期、吐丝期、成熟期。
Fig. 1Rainfall and temperature distribution during spring maize growth stages from 2014 to 2016
The arrows in this figure indicate silking dates of spring maize; V6 and V12 indicate the 6th, 12th leaf collar is visible, respectively; R1 and R6 indicate silking and maturity stage, respectively.
1.2 试验设计
试验设置5个灌水处理: 拔节期灌水(IV6), 大口期灌水(IV12), 抽雄期灌水(IS), 吐丝后15 d灌水(IS15)和不灌水对照(CK)。玉米品种为郑单958, 种植密度为72,000株 hm-2, 播前统一灌底墒水, 每年4月20日播种; 灌水处理采用大水漫灌, 每次灌水75 mm, 利用水表进行计量。2014年大口期有60 mm (连续2 d降水量)降水, 取消了当年IV12处理。试验采取随机区组设计, 3次重复, 小区面积为8 m×10 m。每年播种时每公顷沟施72 kg纯氮(尿素), 105 kg五氧化二磷(磷酸二铵), 120 kg氧化钾(硫酸钾), 大口期每公顷追施108 kg纯氮(尿素), 于9月初收获。其他病虫草害管理参考当地管理模式。1.3 测定项目与方法
1.3.1 土壤水分 分别在播种期、拔节期、大口期、吐丝期和成熟期, 取2 m土体土样, 每20 cm为一层, 测定土壤水分。成熟期利用环刀法测定土壤田间持水量和容重。1.3.2 生物量 于拔节期, 在每小区连续测量5 m 4行玉米的株高和茎粗, 按平均值标记取样植株和后期测量植株, 在大口期(V12)、吐丝期(R1)和成熟期(R6), 每小区取地上部植株2株, 75°C烘干至恒重, 测定植株生物量。
1.3.3 光合速率 于吐丝当天上午10:00—12:00, 利用LI-6400光合仪测定向阳穗位叶片净光合速率, 每小区选择2株, 光辐射强度设置为1200 μmol m-2 s-1, 设定叶室温度比测定时气温低1°C。
1.3.4 穗粒数 在成熟期于每小区中间测产区收获5 m两行的所有果穗, 记录收获穗数, 称量总鲜重, 按平均重量挑选20穗进行考种, 统计穗行数和行粒数, 计算出穗粒数。
1.4 数据分析
利用SPSS 18.0进行方差分析和逐步回归分析, 利用SigmaPlot 14.0进行绘图。2 结果与分析
2.1 不同时期灌水对土壤含水量的影响
2014年春玉米生育期总降水量较少, 但V12时期和花粒期有效的降水保证了籽粒建成及灌浆, 特别是IS处理, 明显提高了籽粒授粉和籽粒建成阶段土壤水分含量(图2-A~D)。2015年降水主要集中在在吐丝后15 d至成熟阶段, 败育籽粒已经形成, 后期降水较多也不能改变籽粒败育率。CK和IS15处理从大口期至籽粒建成期土壤含水明显低于其他处理, 而IV12和IS处理可以缓解春玉米水分临界期的干旱胁迫, 特别是吐丝期灌水, 保证了开花阶段的土壤水分供应(图2-E~I)。2016年春玉米生育期内降水较多(584 mm), 各个处理在生育期内都保持较高的土壤含水量(图2-J~N)。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同时期灌水处理对春玉米各个时期土壤含水量(g g-1)的影响
VS、V6、V12、R1、R6分别代表播种期、拔节期、大口期、吐丝期和成熟期; A~D为2014年, E~I为2015年, J~N为2016年; A、E、J: CK处理; B、F、K: IV6处理; G、L: IV12处理; C、H、M: IS处理; D、I、N: IS15处理。
Fig. 2Effects of irrigating at different growth stages on soil water content (g g-1) at each stage of spring maize
VS, V6, V12 indicate sowing date, the 6th, the 12th leaf collar is visible, respectively; R1 and R6 indicate silking and maturity stage, respectively. A-D, E-I, and J-N indicate in 2014, 2015, and 2016, respectively; A, E, J: the control treatment; B, F, K: IV6 treatment; G, L: IV12 treatment; C, H, M: IS treatment; D, I, N: IS15 treatment.
2.2 不同时期灌水对春玉米穗粒数的影响
由图3可知, 不同灌溉处理年际间穗粒数变化差异明显, 2014年大口期有明显降水, 保证了籽粒形成关键时期的水分供应, 穗粒数较高。其中IS处理穗粒数最高, 分别较CK、IV6、IS15提高了3.3%、6.4%、1.4%。2015年从V10时期到花后15d都未出现有效降水, 严重的干旱胁迫明显提高了空秆率和败育率, 其中CK和IS15处理受到的干旱胁迫最为严重。本研究中对CK处理进行了人工授粉, 降低了该处理的败育率。2015年度穗粒数最高的处理仍为IS, 分别较CK、IV6、IV12和IS15提高了25.2%、13.5%、12.7%和97.0%。2016年度降水较多, 保证了春玉米关键生育时期的水分需求, 但同时也增加了阴雨寡照等不利因素的影响, 各个处理间穗粒数差异不显著且变异度较大。图3
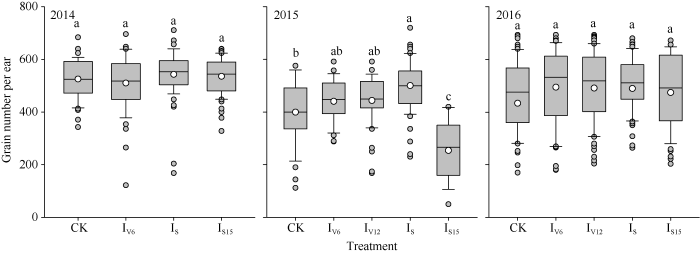 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同时期灌水处理对春玉米穗粒数的影响
图中白色圆点为平均值; 不同字母代表处理间差异显著(P < 0.05); CK、IV6、IV12、IS、IS15分别代表不灌水对照、拔节期、大口期、吐丝期和吐丝后15 d灌水处理。
Fig. 3Effects of irrigating at different growth stages on spring maize kernel number
The white circles are mean values; different letters above the boxes are significantly different among the different treatments at the 0.05 probability level; CK, IV6, IV12, IS, and IS15 indicate the control without irrigation, irrigating when the 6th and 12th leaf collar is visible, silking stage and the 15 day after silking stage, respectively.
2.3 不同时期灌水对生物量、叶片光合速率和ASI的影响
由图4可知, IV6处理明显促进了营养生长, 其花期生物量显著高于其他处理。IV12处理同样可以促进营养生长, 花期生物量同样高于其他处理, 但低于IV6处理。2015年从V10至吐丝遇到干旱胁迫, 花期生物量明显受到影响。IV6和IV12处理增加了大口期和吐丝期的干物质积累, 但成熟期的干物质积累与结实率有关, IS处理在成熟期有较高的生物量。图4
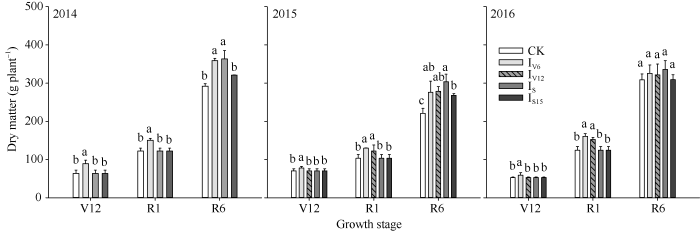 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同时期灌水处理对春玉米生物量的影响
图中不同字母代表处理间差异显著(P < 0.05); V12、R1、R6分别代表大口期、吐丝期、成熟期; CK、IV6、IV12、IS、IS15分别代表不灌水对照、拔节期、大口期、吐丝期和吐丝后15 d灌水处理。
Fig. 4Effects of irrigating at different growth stages on biomass in spring maize
The different letters above the bars are significantly different among the different treatments at the 0.05 probability level; V12, R1, R6 indicate the 12th leaf collar is visible, silking and maturity stage, respectively; CK, IV6, IV12, IS, IS15 indicate the control without irrigation, irrigating at V6, V12, R1, and R6, respectively.
不同时期灌水处理对吐丝期穗位叶光合速率有明显影响。2014年IS处理光合速率最高, CK、IV6和IS15处理之间没有显著差异。2015年春玉米在吐丝期前后遭遇严重的干旱胁迫, IV12和IS处理光合速率最高, CK和IS15处理最低, 但各个处理均低于2014年。2016年各处理均未受到明显的干旱胁迫, 其吐丝期光合速率没有显著差异, 但受阴雨寡照等因素影响, 光合速率较2014年仍有下降(图5-A)。2014年和2016年春玉米开花期未遭遇严重的干旱胁迫, 各个处理的开花吐丝间隔期(ASI)无明显差异, 2015年春玉米关键生育阶段遭遇严重的干旱胁迫, CK和IS15处理的ASI明显延长, 而开花期保持充足的水分供应可以保证玉米花丝正常生长, 避免卡脖旱, 从而缩短ASI提高穗粒数(图5-B)。
图5
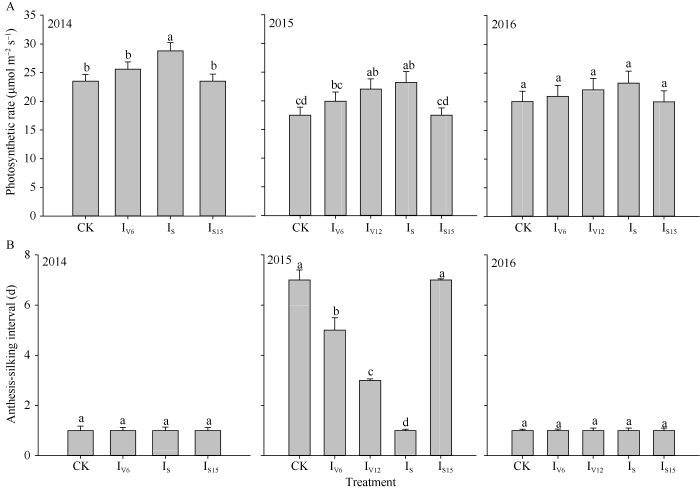 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5不同时期灌水处理对吐丝期春玉米穗位叶光合速率(A)和开花吐丝间隔期(B)的影响
图中不同字母代表处理间差异显著(P < 0.05); CK、IV6、IV12、IS、IS15分别代表不灌水对照、拔节期、大口期、吐丝期和吐丝后15 d灌水处理。
Fig. 5Effects of irrigating at different growth stages on ear leaf photosynthesis at silking (A) and ASI (B) in spring maize
The different letters above the bars are significantly different among the different treatments at the 0.05 probability level; CK, IV6, IV12, IS, IS15 indicate the control without irrigation, irrigating at when the 6th and 12th leaf collar is visible, silking and 15 day after silking stage, respectively.
2.4 开花期土壤水分与穗粒数及叶片光合速率关系
吐丝期是决定玉米穗粒数的关键时期, 该时期充足的水分不仅保证花丝籽粒生长, 降低败育率, 还能保证叶片正常的光合功能, 从而提供充足的碳水化合物保证籽粒生长。逐步回归分析结果表明, 吐丝期土壤含水量越高, 穗位叶光合速率越高, 而在2016年, 过多的降水虽然提高了土壤含水量, 但光合速率降低, 即较多的阴雨寡照天气不利于叶片光合作用。土壤含水量与穗粒数间的回归分析同样表明, 提高土壤含水量有利于穗粒数的增加, 但过多降水会导致阴雨寡照天气增多(2016年), 降低春玉米结实率。另外, 吐丝期穗位叶光合速率的提高, 保证了充足的碳水化合物供应, 有利于春玉米穗粒数的提高(图6)。图6
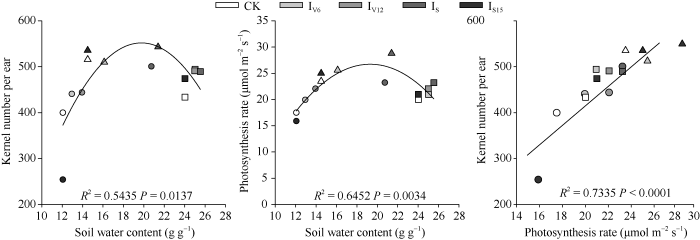 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6吐丝期土壤含水量与光合速率及穗粒数间关系
三角形、圆形和方形分别为2014年、2015年和2016年; CK、IV6、IV12、IS、IS15分别代表不灌水对照、拔节期、大口期、吐丝期和吐丝后15 d灌水处理。
Fig. 6Relationships between soil water content with photosynthesis rate and kernel number per ear at silking stage
The triangle, circle, and square indicate in 2014, 2015, and 2016, respectively; CK, IV6, IV12, IS, IS15 indicate the control without irrigation, irrigating at when the 6th and 12th leaf collar is visible, silking and 15 day after silking stage, respectively.
3 讨论
3.1 不同时期灌水对春玉米穗粒数的影响
在种植密度既定的前提下, 保证穗粒数是提高玉米产量的关键。干旱胁迫, 特别是花期干旱胁迫, 是影响玉米籽粒败育的重要因素。尽管华北平原春玉米开花期(播期4月至5月, 开花期6月下旬至7月下旬)分布在雨季, 但该地区年际间和月份间降水变异度极大, 春玉米花期仍然有较大概率遭遇干旱胁迫[17]。V6和V12时期灌溉保证了玉米的营养生长, 但开花期干旱胁迫仍会显著降低玉米穗粒数。一般认为花期干旱胁迫降低了碳水化合物供应, 从而导致籽粒中葡萄糖和淀粉含量降低, 籽粒败育[22]。Boyer团队利用茎部注射蔗糖试验, 明确了增加碳水化合物供应可以缓解干旱胁迫造成的败育损失[22,23]。还有研究认为干旱降低籽粒活性, 细胞壁转化酶活性降低, 相关基因(INCW2)表达量下调是引起籽粒败育的重要因素[24,25]。Turc团队则认为干旱胁迫影响花丝水势进而抑制花丝生长, 导致顶部花丝吐出较晚不能授粉, 这是大田生产条件下玉米籽粒败育的关键因素[20,26]。尽管前人研究对花期干旱诱导籽粒败育的机制有不同观点, 但灌溉是维持花丝水势、碳水化合物供应和籽粒活性的重要措施。本研究结果表明花期灌水提高了穗位叶光合速率(碳水化合物), 维持了正常的ASI, 尽管籽粒活性没有测定, 但最终穗粒数明显提高。前人模型研究结果同样表明花期灌水可以提高春玉米产量和水分利用效率[15]。花后15 d灌溉时穗粒数已经决定, 在穗粒数受影响较小的年份(2014年和2016年), 该处理可以保证灌浆期籽粒生长; 但在籽粒败育严重的2015年, 花后灌溉会增加玉米耗水量, 但对产量影响较小[17]。因此, 我们认为花期灌水是保证玉米关键生育时期花丝和籽粒正常生长, 提高春玉米穗粒数的关键措施。3.2 华北地区春玉米灌溉策略
华北地区春玉米任何生育时期都可能遭遇干旱胁迫, 特别是早播玉米(4月和5月上旬), 在营养生长阶段有较大概率遭遇干旱胁迫(图7)。2016年尽管降水较多, 但拔节期前后同样遭遇干旱胁迫, LAI和生物量明显降低[17]; 2015年决定穗粒数的关键时期春玉米遭遇严重的干旱胁迫, LAI和生物量同样受到严重影响, 但吐丝期灌水仍然保证了较高的穗粒数[17]; 相反, 拔节期灌水明显提高了LAI, 但会增大蒸腾耗水, 尽管有较大的光合面积和花前生物量的积累, 但仍然不能保证较高的穗粒数和产量[17]。2014年春玉米生育期降水量最少, 但穗粒数和产量最高, 有限的降水恰好分布在春玉米用水的关键时期, 保证了籽粒建成和灌浆。因此, 在决定玉米穗粒数的关键生育时期(也是玉米水分需求临界期)保证水分供应是提高水分利用效率和穗粒数与产量形成的关键。而在阴雨寡照较多的年份, 灌水不是决定穗粒数和产量的关键, 维持叶片和根系功能, 防止玉米早衰至关重要。图7
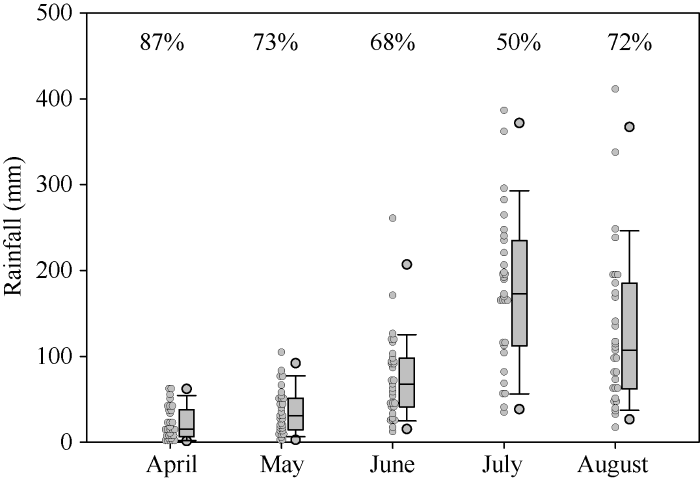 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图71986-2016年试验点各个月份降水分布图
图中百分数字代表各个月份降水量变异系数。
Fig. 7Rainfall distribution in each month at experimental site from 1986 to 2016
The data above boxes indicate the coefficient of variation in each month.
4 结论
在华北地区, 把有限的水分资源集中在决定春玉米穗粒数形成的关键时期, 即保证花期水分供应,是提高春玉米穗粒数的关键。尽管春玉米营养生长阶段会遭遇干旱胁迫, 降低叶面积指数和花前生物量积累, 但保证吐丝时的水分供应, 可以提高叶片光合速率, 增加穗粒数。相反, 保证了营养生长阶段的水分供应, 提高了春玉米的叶面积指数和生物量, 但也会增加蒸腾耗水, 花期干旱需要二次灌溉, 从而增加灌溉耗水。因此,花期灌溉是降低灌溉用水,提高春玉米穗粒数和产量的关键措施。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
DOI:10.1016/j.pbi.2008.12.006URLPMID:19179104 [本文引用: 1]

Gene expression driven by developmental and stress cues often depends on nucleosome histone post-translational modifications and sometimes on DNA methylation. A number of studies have shown that these DNA and histone modifications play a key role in gene expression and plant development under stress. Most of these stress-induced modifications are reset to the basal level once the stress is relieved, while some of the modifications may be stable, that is, may be carried forward as 'stress memory' and may be inherited across mitotic or even meiotic cell divisions. Epigenetic stress memory may help plants more effectively cope with subsequent stresses. Comparative studies on stress-responsive epigenomes and transcriptomes will enhance our understanding of stress adaptation of plants.
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 3]
[本文引用: 3]
[本文引用: 2]
[本文引用: 10]
DOI:10.1093/jxb/erx452URLPMID:29304259 [本文引用: 1]
Maize grain yield is highly related to the number of kernels that are established during the flowering period. Kernel number depends on the accumulation of ear biomass and the efficiency of using this biomass for kernel set. Ear biomass depends on the rate of plant biomass accumulation and the proportion of this biomass that is allocated to the ear. In contrast to other major crops, the proportion of plant biomass that is allocated to the ear is not constant in maize, being almost zero under stress conditions. Fortunately, there is wide native genetic variability for this trait, with major practical implications for crop management and plant breeding. Conditions that inhibit plant growth commonly delay silk appearance relative to male anthesis. Time to silking and silk extrusion, which is a tissue expansion process, is dependent on water turgor and ear biomass accumulation, and the magnitude of this delay is used as a marker to phenotype for stress susceptibility. Ear biomass accumulation can also be used for predicting the number of silks that have been extruded if genotype-specific parameters are known. Here, several mechanistic plant and canopy traits are described, together with their implications for better understanding maize yield determination under limited plant growth environments. An ideal genotype sustains growth in environments with limited water or nutrients, has uniform canopies, has increased biomass partitioning to the ear at reduced plant growth, reaches silking with minimum ear biomass, and has rapid silk extrusion for minimizing developmental delays between competing structures within the ear. All these traits help maximize kernel set and yield at limited plant growth, and most have been indirectly selected by breeders when increasing yield.
[本文引用: 1]
[本文引用: 1]
DOI:10.1104/pp.15.01130URLPMID:27208256 [本文引用: 2]
Flower or grain abortion causes large yield losses under water deficit. In maize (Zea mays), it is often attributed to a carbon limitation via the disruption of sucrose cleavage by cell wall invertases in developing ovaries. We have tested this hypothesis versus another linked to the expansive growth of ovaries and silks. We have measured, in silks and ovaries of well-watered or moderately droughted plants, the transcript abundances of genes involved in either tissue expansion or sugar metabolism, together with the concentrations and amounts of sugars, and with the activities of major enzymes of carbon metabolism. Photosynthesis and indicators of sugar export, measured during water deprivation, suggested sugar export maintained by the leaf. The first molecular changes occurred in silks rather than in ovaries and involved genes affecting expansive growth rather than sugar metabolism. Changes in the concentrations and amounts of sugars and in the activities of enzymes of sugar metabolism occurred in apical ovaries that eventually aborted, but probably after the switch to abortion of these ovaries. Hence, we propose that, under moderate water deficits corresponding to most European drought scenarios, changes in carbon metabolism during flowering time are a consequence rather than a cause of the beginning of ovary abortion. A carbon-driven ovary abortion may occur later in the cycle in the case of carbon shortage or under very severe water deficits. These findings support the view that, until the end of silking, expansive growth of reproductive organs is the primary event leading to abortion, rather than a disruption of carbon metabolism.
DOI:10.1093/jxb/ery013URLPMID:29365129 [本文引用: 1]
Selective seed abortion is a survival strategy adopted by many species that sacrifices some seeds to allow the remaining ones to set. While in evolutionary terms this is a successful approach, it causes huge losses to crop yields. A pollination time gap (PTG) has been suggested to be associated with position-related grain abortion. To test this hypothesis, we developed a novel approach to alter the natural pattern of maize (Zea mays L.) pollination and to examine the impact of PTGs on kernel growth and the underlying physiological basis. When apical and basal kernels were synchronously pollinated, the basal kernels set and matured but the apical kernels were aborted at an early stage. Delaying pollination to the basal ovaries suppressed their development and reduced invertase activity and sugar levels, which allowed the apical kernels to set and grow normally. In situ localization revealed normal cell wall invertase activity in apical and basal kernels under synchronous pollination but reduced activity in the delayed-pollinated kernels independent of their position. Starch, which was abundant in basal kernel areas, was absent in the apical kernel regions under synchronous pollination but apparent with delayed pollination. Our analyses identified PTG-related sink strength and a low level of local assimilates as the main causes of grain abortion.
DOI:10.1104/pp.121.1.25URLPMID:10482657 [本文引用: 2]
After reproduction is initiated in plants, subsequent reproductive development is sometimes interrupted, which decreases the final number of seeds and fruits. We subjected maize (Zea mays L.) to low water potentials (psi(w)) that frequently cause this kind of failure. We observed metabolite pools and enzyme activities in the developing ovaries while we manipulated the sugar stream by feeding sucrose (Suc) to the stems. Low psi(w) imposed for 5 d around pollination allowed embryos to form, but abortion occurred and kernel number decreased markedly. The ovary contained starch that nearly disappeared during this abortion. Analyses showed that all of the intermediates in starch synthesis were depleted. However, when labeled Suc was fed to the stems, label arrived at the ovaries. Solute accumulated and caused osmotic adjustment. Suc accumulated, but other intermediates did not, showing that a partial block in starch synthesis occurred at the first step in Suc utilization. This step was mediated by invertase, which had low activity. Because of the block, Suc feeding only partially prevented starch disappearance and abortion. These results indicate that young embryos abort when the sugar stream is interrupted sufficiently to deplete starch during early ovary development, and this abortion results in a loss of mature seeds and fruits. At low psi(w), maintaining the sugar stream partially prevented the abortion, but invertase regulated the synthesis of ovary starch and partially prevented full recovery.
[本文引用: 1]
DOI:10.1093/jxb/erl177URLPMID:17105969 [本文引用: 1]
In many situations, organisms respond to stimuli by altering the activity of large numbers of genes. Among these, certain ones are likely to control the phenotype while others play a secondary role or are passively altered without directly affecting the phenotype. Identifying the controlling genes has proven difficult. However, in a few instances, it has been possible to reverse the phenotype by physiological or biochemical means without altering the genetics of the organism. During this functional reversion, only a few genes may respond, thus identifying those likely to be controlling the phenotype. Floral abortion during a water shortage in maize is an example because the response is inherently multigenic, and the phenotype can be reversed by physiological/biochemical means. A recent analysis used this reversal to reveal that only a few genes are likely to control the abortion phenotype. In maize, these genes coded for a cell wall invertase (Incw2), a soluble invertase (Ivr2), a ribosome-inactivating protein (RIP2), and phospholipase D (PLD1). The invertases appeared to control the normal sugar uptake by the ovaries. Their down-regulation depleted ovary sugar pools and resulted in an up-regulation of the genes for ribosome-inactivating protein and for phospholipase. The latter changes appeared to initiate senescence that degraded cell membranes, thus causing irreversible abortion. With these findings, these genes have become targets for preventing abortion. This approach might have value in other contexts with some additional methods.
DOI:10.1016/j.tplants.2012.06.005URLPMID:22776090 [本文引用: 1]
Seed and fruit set are established during and soon after fertilization and determine seed and fruit number, their final size and, hence, yield potential. These processes are highly sensitive to biotic and abiotic stresses, which often lead to seed and fruit abortion. Here, we review the regulation of assimilate partitioning, including the potential roles of recently identified sucrose efflux transporters in seed and fruit set and examine the similarities of sucrose import and hydrolysis for both pollen and ovary sinks, and similar causes of abortion. We also discuss the molecular origins of parthenocarpy and the central roles of auxins and gibberellins in fruit set. The recently completed strawberry (Fragaria vesca) and tomato (Solanum lycopersicum) genomes have added to the existing crop databases, and new models are starting to be used in fruit and seed set studies.
DOI:10.1104/pp.15.00268URLPMID:26598464 [本文引用: 1]
Grain abortion allows the production of at least a few viable seeds under water deficit but causes major yield loss. It is maximum for water deficits occurring during flowering in maize (Zea mays). We have tested the hypothesis that abortion is linked to the differential development of ovary cohorts along the ear and to the timing of silk emergence. Ovary volume and silk growth were followed over 25 to 30 d under four levels of water deficit and in four hybrids in two experiments. A position-time model allowed characterizing the development of ovary cohorts and their silk emergence. Silk growth rate decreased in water deficit and stopped 2 to 3 d after first silk emergence, simultaneously for all ovary cohorts, versus 7 to 8 d in well-watered plants. Abortion rate in different treatments and positions on the ear was not associated with ovary growth rate. It was accounted for by the superposition of (1) the sequential emergence of silks originating from ovaries of different cohorts along the ear with (2) one event occurring on a single day, the simultaneous silk growth arrest. Abortion occurred in the youngest ovaries whose silks did not emerge 2 d before silk arrest. This mechanism accounted for more than 90% of drought-related abortion in our experiments. It resembles the control of abortion in a large range of species and inflorescence architectures. This finding has large consequences for breeding drought-tolerant maize and for modeling grain yields in water deficit.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}