 ,*, 蔡一林,*西南大学玉米研究所 / 南方山地农业教育部工程研究中心, 重庆400715
,*, 蔡一林,*西南大学玉米研究所 / 南方山地农业教育部工程研究中心, 重庆400715Genetic analysis and molecular characterization of tasselseed mutant ts12 in maize
QIN Li-Ping**, DONG Er-Fei**, BAI Yang, ZHOU Lian, REN Lan-Yang, ZHANG Ren-Feng, LIU Chao-Xian,*, CAI Yi-Lin,*Maize Research Institute, Southwest University / Engineering Research Center of South Upland Agriculture, Ministry of Education, Chongqing 400715, China通讯作者:
第一联系人:
收稿日期:2019-09-16接受日期:2020-01-15网络出版日期:2020-01-23
Received:2019-09-16Accepted:2020-01-15Online:2020-01-23
作者简介 About authors
秦利萍,E-mail:919494709@qq.com。

摘要
关键词:
Abstract
Keywords:
PDF (3390KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
秦利萍, 董二飞, 白洋, 周练, 任岚扬, 张任凤, 刘朝显, 蔡一林. 玉米tasselseed突变体ts12的遗传分析与分子鉴定[J]. 作物学报, 2020, 46(5): 690-699. doi:10.3724/SP.J.1006.2020.93051
QIN Li-Ping, DONG Er-Fei, BAI Yang, ZHOU Lian, REN Lan-Yang, ZHANG Ren-Feng, LIU Chao-Xian, CAI Yi-Lin.
玉米(Zea mays L.)是典型的雌雄同株、雌雄异花植物, 是研究性别决定的重要模式植物之一[1]。玉米花序发育过程中产生了4种类型的侧生分生组织, 在发育中按产生先后顺序分为侧支分生组织、小穗成对分生组织、小穗分生组织和小花分生组织[2]。玉米小花分生组织继续分化形成包括3个雄蕊和3个心皮的两性花, 随后雄穗中的雌蕊原基和雌穗中的雄蕊原基分别发生选择败育, 最终形成了单性花, 这一性别决定过程是一个复杂的生物学过程, 受到植物激素、遗传因子和环境因素的共同调控[3]。
大量研究表明植物激素茉莉酸(jasmonic acid, JA)、赤霉素(GA)和油菜素内酯(BR)等参与了玉米性别决定过程。前人研究表明Ts1、Ts2、ts5、Sk1、OPR7和OPR8与JA信号通路有关, 该通路调控了玉米雄蕊和雌蕊的发育。Ts1编码一个脂氧合酶[4], 是JA合成途径中的一个关键酶; 其功能的缺失导致脂氧合酶活性消失, 玉米内源JA浓度下降, 最终导致雄穗性别反转, 小穗雌性化, 产生大量花丝。Ts2基因编码一个短链乙醇脱氢酶[5], 该酶可能会产生一个促进凋亡的信号或者降解细胞存活所需要的底物[6,7]; Ts2基因功能的丧失同样会导致玉米雄穗产生与ts1突变体类似的tasselseed表型。不同于Ts1和Ts2基因功能丧失后表现为隐性突变, Ts5为显性突变体, 除了雄穗小穗性别发生反转, 产生雌性化的小穗外, 雌穗下位花不能败育, 导致果穗籽粒排列紊乱; ts5编码一个创伤诱导表达的蛋白酶ZmCYP94B1, 该酶可使有生物活性的JA和异亮氨酸偶联物JA-Ile失活, 影响了JA的代谢, 致使植株内源JA含量降低, 最终影响性别决定[8]。对ts1、ts2和Ts5突变体施加外源JA, 均可恢复小穗雄蕊发育, 产生有功能的花粉[4,8], 该结果进一步证实了这3个基因与JA代谢和信号转导过程密切相关。在玉米雄穗发育过程中, Ts1和Ts2基因调控了雄穗和雌穗下位花雌蕊的败育, 与此相反, Sk1基因促进雌蕊的发育[9]。Sk1基因突变后, 导致雌穗不能正常产生花丝; Sk1基因过表达导致T0代转基因植株雄穗完全雌性化, 雌穗上位花和下位花雌蕊均可发育。Sk1基因编码一个UDP-糖基转移酶, 该酶可能使过氧化物酶体中的JA或者JA前体失活, 从而破坏了JA信号通路, 使Ts1和Ts2调控的雌蕊败育过程受阻[10]; Sk1发挥功能的区域因受到Rmr6基因的限制, 而仅限于雌穗上位花, 从而保证了雄穗中的雌蕊和雌穗下位花雌蕊的败育[11]。与拟南芥OPR3直系同源的玉米基因OPR7和OPR8, 在单个基因功能丧失情况下, JA合成没有受到明显影响, 雄穗和雌穗均能正常发育; 然而在opr7opr8双突变体中, JA的含量显著下降, 雄穗明显表现出类似ts1和ts2的雌性化特征, 施加外源JA则可使雄穗发育恢复正常[12,13]。
除了JA, 赤霉素在玉米性别决定中也扮演了重要角色。矮秆突变体d1、d2、d3、d5和an1均表现为雄蕊不能正常败育[14], 这是因为赤霉素正常合成途径受到抑制造成的[15,16], 施加外源赤霉素则可恢复正常表型[17]。同时用外源赤霉素对性别分化前正常植株持续处理, 会导致部分雄穗小穗雄蕊完全转变成雌蕊[18], 这充分说明了赤霉素在玉米雌蕊发育中发挥重要作用。亦有研究表明油菜素内酯等激素参与了玉米性别决定过程。通过对经典矮秆突变体na1的鉴定, 发现na1植株在BR生物合成途径中携带一个功能缺失突变的DET2同源基因[19], 导致DET2特异性底物累积, 下游BR代谢物减少。用BR生物合成抑制剂处理野生型玉米植株完全模拟了na1突变体的矮化和tasselseed表型, 揭示了BR对玉米性别的控制作用。此外, Ts4和ts6分子机制的阐释, 揭示了其在转录调控水平上, 而不是在激素的生物合成上发挥的重要作用。ts6编码一个花同源异型基因APETALA2-like转录因子, 在小穗分生组织的确定性方面具重要功能; Ts4编码一个microRNA zma-MIR172e, 其调控了ts6基因的表达[20]。
我们利用EMS诱变B73花粉, 获得了一个tasselseed突变体ts12。本研究对ts12的表型进行了鉴定, 利用F2作图群体开展了目标基因的分子定位, 并与ts2突变体进行了等位性测验。该研究为进一步深入解析玉米性别决定分子机制提供了有价值的参考。
1 材料与方法
1.1 实验材料
ts2突变体(ts2-N2409)来自Maize Genetics Cooperation Stock Center (http://maizecoop.cropsci.uiuc. edu/)。2016年3月于重庆市北碚区西南大学歇马实验基地种植自交系B73和Mo17。参照Neuffer等[21]的方法, 用EMS和液体石蜡配制体积比6.7×10-4:1的工作液对B73花粉进行诱变处理, 将诱变后的花粉用毛笔涂抹于Mo17花丝; 同年9月于云南玉溪播种获得的M0代籽粒并自交。2017年6月在诱变群体中鉴定出3个雄穗结实突变体, 依次命名为ts10、ts11和ts12; 同时用ts12与Mo17建构F2分离群体, 用于ts12的分子定位。2018年6月, 在ts2植株雌性化雄穗未抽出前套袋, 待雌性化雄穗抽出后, 把ts12花粉授予ts2植株雌性化雄穗, 得到F1代杂交籽粒; 同年9月把杂交后代种植于云南玉溪, 观察F1代植株雄穗发育形态, 且对能产生花粉的植株进行自交。1.2 Simple sequence repeat (SSR) 标记开发及连锁标记筛选
从Gramene (http://www.gramene.org/)数据库下载玉米基因组序列, 用SSR Hunter[22]搜索构成重复元件核苷酸数最多是4个, 重复次数最少为4次的SSR序列; 然后在NCBI网站 (https://blast.ncbi. nlm.nih.gov/Blast.cgi)进行Blast比对, 以单拷贝SSR序列为模板用Primer-BLAST (https://www.ncbi.nlm. nih.gov/tools/primer-blast/index.cgi)设计引物。正向引物位于模板1~150 bp之间, 反向引物位于170~308 bp之间, 扩增片段最低120 bp, 正、反向引物长22 bp, 最短20 bp, 最长不超过25 bp, GC含量在40%~60%之间, 正、反向引物退火温度差异不超过1℃, 其他参数为默认设置。然后利用开发的SSR标记对B73和Mo17基因组DNA进行扩增, PCR产物经6%聚丙烯酰胺电泳及银染后判断其是否具有多态性[23]。在F2分离群体中, 根据表型分别取10株正常植株和突变植株叶片提取DNA, 等量混合构建野生型DNA池和突变体DNA池; 用在B73和Mo17之间具有多态性的432对SSR标记[24]对2个DNA池扩增, 鉴定出与ts12连锁的分子标记。用筛选出的及新开发的SSR标记(附表1)对所有ts12突变体个体的基因型进行分析, 鉴定出发生染色体片段交换的突变植株, 对ts12基因精细定位。
Supplementary table 1
附表1
附表1ts12基因定位、候选基因克隆及表达分析引物序列
Supplementary table 1
| 引物名称 Primer name | 正向引物序列 Forward primer sequence (5′-3′) | 反向引物序列 Reverse primer sequence (5′-3′) | 标记类型 Genetic marker types |
|---|---|---|---|
| LM1 | CACAGCAAGAGTACAGCATCAG | TGATCCCAGTTCTCGATGTAGC | SSR |
| LM2 | GGAAATGTTTGACGTGACCTGC | CACTTGGCAAACCAATCCAACG | SSR |
| LM3 | AATAGCGTGCTCCCGTGTTATC | CTGTCGTTGCTTGCTTCTTCCA | SSR |
| LM4 | AATGGGATAAAGACCGGATTCT | ATAAGCGTTGTAGGACAGGAGC | InDel |
| RM1 | TCGGAGAGGACACGGTTTAGTA | CATGAACATTGGCGAAGCTACC | SSR |
| RM2 | GCTCCTGTGTGCAACTTGTTAG | GCATATGAATCGGTGTTCGGTG | SSR |
| RM3 | CTGTTCCTTCGCTTTCACAGAA | AGTAGCTCTCTAGTTGTGTCCC | SSR |
| RM4 | GCTGATTTAAGATATAAAGTGCCTCCC | CCTGCTGGACCGGAGTAAAG | InDel |
| RM5 | AGAACGGTGATTTTTGTCTGGT | GTTTTGCATGCTCATGTAGACG | InDel |
| Ts2-JD | ACAGCAGAGTAGAGTAGCACAC | GCCATGGGAATGGGATATTTGG | |
| Ts2-qPCR | GTGGAGAAGATGGAGGAGGTGGT | ATTGATTCACAAGCCGATGAGGTT | |
| Actin | TCACCCTGTGCTGCTGACCG | GAACCGTGTGGCTCACACCA |
新窗口打开|下载CSV
1.3 ts12形态学观察
将F2分离群体播于大田, 20 d后取幼苗叶片, 提取DNA, 进行个体基因型鉴定。筛选出带型与ts12突变体、杂合带型植株及与Mo17一致的纯合带型植株(野生型)。待玉米生长至V7时期, 剥离不同发育阶段ts12和野生型植株新鲜未成熟雄穗, 直接在扫描电镜(Hitachi SU3500, Tokyo, Japan)下观察其形态结构变化。1.4 外源JA处理
将JA溶于无水乙醇, 配成1 mmol L-1的工作液, 保存于4℃; 以0.05%乙醇为对照。待F2群体中ts12植株及野生型植株生长至35 d, 每天下午5:00左右用移液器吸取1 mL 1 mmol L-1 JA溶液和0.05%乙醇注入玉米心叶, 持续处理14 d。统计JA、乙醇处理及无任何处理的ts12雄穗结实数。1.5 精细定位区间基因注释
在Gramene网站下载ts12精细定位区间内基因及其编码蛋白序列, 然后以蛋白序列为种子序列Blast比对NCBI蛋白质无冗余数据库(Non-redundant protein sequences), 搜索目标蛋白在其他物种中的注释及功能, 从而对玉米中的同源基因进行功能注释。1.6 ts12突变体中ts2基因的克隆和序列分析
根据ts2基因(Zm00001d028806)序列, 利用Primer-BLAST设计扩增引物(附表1)扩增B73和ts12雄穗cDNA。PCR扩增体系含模板cDNA 1 μL、2 × KOD buffer 10 μL、KOD FX NEO (1.0 U μL-1) 0.5 μL、引物 (10 μmol L-1) 1.5 μL、dNTPs (10 mmol L-1) 0.2 μL, 添加ddH2O至20 μL。PCR扩增条件为94℃预变性5 min, 94℃变性10 s, 62℃退火30 s, 68℃延伸1 min, 扩增35个循环, 最后68℃延伸5 min。用1%琼脂糖凝胶电泳检测, 胶回收PCR产物后连接到PEASY-Blunt载体上, 连接产物转化大肠杆菌感受态细胞, 经过PCR鉴定后, 选取含有目的条带的5个菌株样本送上海英骏生物技术有限公司测序。使用DNAMAN (https://www.lynnon.com/)比对分析测序结果。1.7 RNA提取及基因表达量分析
对ts12 F2分离群体植株基因型鉴定后, 取V7时期ts12及野生型植株的根、茎、叶、雄穗(1~2 cm), V9时期的雌穗(1~2 cm), 及未抽出苞叶的花丝和授粉后20 d的胚、胚乳, 速冻于液氮, 于-80℃保存。在150℃烘箱中对研钵处理4 h后, 将各组织研磨。用RNA提取试剂盒(RNAprep Pure Plant Kit)和RNA反转录试剂盒(RevertAid RT Reverse Transcription Kit)提取RNA及合成cDNA第1链。以cDNA为模板, 用定量引物Ts2-qPCR (附表1), 以actin为内参, 在实时定量PCR仪(Analytik Jena qTOWER, Jena, Germany)上定量分析。PCR扩增体系含cDNA 1 μL、引物0.75 μL、SYBR 6.25 μL, 补超纯水至13 μL。PCR扩增条件为95℃预变性3 min; 95℃变性15 s, 62℃退火15 s, 循环数为40。使用公式2-ΔΔCT计算基因相对表达量。2 结果与分析
2.1 ts12突变体表型观察及遗传方式分析
ts12突变体营养生长阶段发育正常, 与野生型植株没有明显差异; 然而抽雄后成熟雄穗产生大量花丝(图1-B), 授粉后能正常结实; 同时发现雄穗主穗及侧枝顶端部分小穗仍能正常发育、散粉(图1-C), 自交后可获得纯合突变体。在扫描电镜下观察V7阶段未成熟雄穗可见, 野生型未成熟雄穗长约12 mm时, 小穗分生组织分化产生雄蕊和雌蕊(图1-D), 但未见雌蕊产生心皮组织; 相反, 在ts12未成熟雄穗长约13 mm时, 则明显可见雌蕊产生胚珠原基和心皮结构(图1-E)。表明ts12突变体雄穗长约13 mm时小穗发育异常, 表现出了雌性化的特征。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1ts12表型观察
A和B: 野生型和ts12成熟雄穗; C: ts12雄穗上能正常发育产生花粉的小穗; D和E: 未成熟野生型雄穗(12 mm)和ts12雄穗(13 mm)扫描电镜观察。WT: 野生型; G: 雌蕊; ST: 雄蕊; C: 心皮。
Fig. 1Phenotype of ts12
A and B: mature tassel of the wild type and ts12; C: the normal spikelets that could produce pollens on ts12 tassels. D and E: immature tassel of the wild type (12 mm) and ts12 (13 mm) under scanning electron microscope. WT: wild type; G: gynoecium; ST: stamen; C: carpel.
用ts12纯合突变体与Mo17杂交产生的F1代雄穗发育为正常植株, 然后自交构建了F2分离群体。在F2代群体150个植株中, 突变植株35株, 正常植株115株(表1)。经过卡平方检验, 野生型和突变体符合3:1的分离比例(χ2 = 0.22 < χ20.05 = 3.84), 说明ts12突变体是由隐性单基因控制。
Table 1
表1
表1ts12显隐性的分离比检验
Table 1
| 群体组配 Population construction | F1植株雄穗表型 Phenotype of F1 plants | F2分离群体 F2 segregation population | F2植株总数 Total number of F2 plants | χ2 | |
|---|---|---|---|---|---|
| 野生型 WT | ts12 | ||||
| ts12 × Mo17 | 正常 Normal | 115 | 35 | 150 | 0.22# |
新窗口打开|下载CSV
2.2 ts12精细定位
用在B73和Mo17之间有多态性且能覆盖整个玉米基因组的432对SSR标记, 对构建的ts12突变体DNA池和野生型植株DNA池进行连锁标记的筛选。结果显示第1染色体1.04 bin的SSR标记umc1169的扩增产物在两个DNA混合池之间具有多态性(图2-A); 用该标记对15个ts12植株基因型鉴定发现, 除了交换单株基因型为杂合外(图2-B), 其他植株基因型均与表型一致, 推测umc1169与ts12呈连锁关系。在umc1169附近开发了6个SSR标记(附表1), 并分别对F2群体中501株ts12突变体的基因型进行鉴定。在umc1169位点鉴定交换单株86个, 且在这86株中分别鉴定出24、12、6个植株分别在RM1、RM2和RM3位点发生了交换; 在LM1位点鉴定交换单株28个, 且在28个重组植株中, 有10株在LM2位点发生了交换, 从而把目标基因ts12初步定位于SSR标记LM2和RM3之间, 物理距离约为1.6 Mb的区间内(图3-A)。然后继续开发SSR和InDel标记, 并分别在LM3、LM4位点鉴定4个和2个交换单株, 在RM4和RM5位点分别鉴定交换单株2个和1个, 最终把目标基因定位于InDel标记LM4和RM5之间约290 kb的物理区间内(图3-B)。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2ts12连锁标记umc1169的筛选及验证
MT: ts12 DNA混合池; WT: 野生型DNA混合池。
Fig. 2Screening and confirmation of SSR markers umc1169 linked with ts12
MT: DNA pool of ts12; WT: DNA pool of wild-type plants.
图3
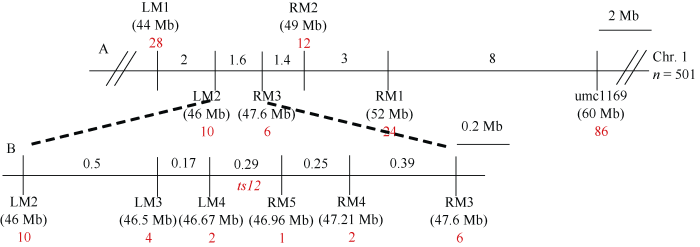 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3 ts12精细定位
黑色实线上方相邻标记之间的数字代表物理距离, 括号内数字代表标记在染色体上的确切位置, 标记下方的红色数字代表交换单株数目。
Fig. 3Fine mapping of ts12
The numbers above the solid back line between SSR markers represent physical distance. The numbers in parenthesis represent the exact position of the marker on chromosome. The red numbers under SSR markers represent recombination events.
2.3 ts12候选基因及突变位点分析
ts12定位区间290 kb内共含有9个注释基因(表2), 其中包括已经克隆的玉米性别决定基因Ts2 (Zm00001d028806), 推测ts12突变表型可能是Ts2基因突变造成的。提取ts12纯合突变体雄穗总RNA, 反转录合成cDNA。设计Ts2-JD引物(附表1)扩增Ts2基因序列, 扩增片段序列与B73 Ts2 CDS比对发现第196个碱基鸟嘌呤G被替换为腺嘌呤A (图4), 导致该位点编码氨基酸甘氨酸被替换为精氨酸; 该突变位点位于Ts2蛋白乙醇脱氢酶保守结构域A中。截取Ts2突变位点附近DNA序列与B104、EP1、F7、Mo17、PH207和W22 自交系中Ts2基因相应位置序列比对发现, 该位点在其他自交系中非常保守, 没有SNP变异(图5)。由此推测ts2基因中单核苷酸的转换可能是导致tasselseed表型的原因。Table 2
表2
表2ts12定位区间内基因功能注释
Table 2
| 基因 ID Gene ID | 基因注释 Gene annotation |
|---|---|
| Zm00001d028796 | Meristematic receptor-like kinase |
| Zm00001d028797 | Laccase-13 |
| Zm00001d028798 | Hypothetical protein |
| Zm00001d028799 | CASP-like protein 4B1 |
| Zm00001d028801 | 26S proteasome regulatory subunit 4 homolog A |
| Zm00001d028802 | Hypothetical protein |
| Zm00001d028803 | Protein SCAR2 |
| Zm00001d028804 | U2 small nuclear ribonucleoprotein B |
| Zm00001d028806 | Sex determination protein Tasselseed2 |
新窗口打开|下载CSV
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4B73和ts12中Ts2基因CDS序列比对
红色下画线部分代表Ts2保守结构域A, 绿色方框代表Ts2中氨基酸发生替换的位点
Fig. 4CDS alignment of Ts2 genes from B73 and ts12 mutant。。
The parts underlined in red color represent the conserved domain A of Ts2. The green rectangle indicates the mutation site of the amino acid substitution in Ts2. 。。
图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5ts12突变体和6个自交系中的Ts2基因突变位点序列比对。。
蓝色方框代表Ts2基因的单核苷酸突变位点
Fig. 5Sequence alignment of Ts2 partial fragment from ts12 and six inbred lines 。。
The blue rectangle indicates the single nucleotide mutation site of the Ts2. 。。
2.4 ts12和ts2突变体等位性测验
ts12突变体雌性化雄穗主穗及侧枝顶部个别小穗可正常发育产生有功能的花粉, 而ts2突变体雄穗小穗完全雌性化, 不能产生有功能的花粉(图6-A), 因此取ts12植株花粉对ts2雌性化雄穗花丝授粉, 3 d后重复授粉一次。将获得的F1代植株播于大田, 待抽雄后观察雄穗表型。通过对56个F1植株表型的观察, 发现F1植株雄穗均发生雌性化(图6-B), 可产一样能正常发育产生少量有功能的花粉, 可产生自交后代F2; 进一步观察113株F2雄穗表型, 发现所有植株均产生与F1植株一样的雌性化雄穗。由此推断ts12是ts2的等位突变体, 即ts12突变表型是由编码乙醇脱氢酶基因Ts2的突变引起的。图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6ts12和ts2突变体等位测验
A: ts2植株雌性化雄穗; B: ts12和ts2等位测验后代表型。
Fig. 6Allelism test of ts12 with ts2
A: the feminized tassel of ts2 mutant; B: the mutant phenotype in the offspring of allelism test of ts12 and ts2.
2.5 外源植物激素JA处理
JA在玉米性别决定调控中发挥重要作用, 已知ts2在外源JA处理后雌性化雄穗可恢复正常[4]。用1 mmol L-1 JA溶液、0.05%乙醇分别处理23株和16株生长35 d的ts12突变体, 雄穗发育成熟后, 分别观察小穗性别恢复情况, 并统计雄穗结实数(图7)。未实施任何处理的15株和用0.05%乙醇处理的16株ts12突变体平均结实数分别为133粒和144粒, 雄穗结实数在统计学上并无明显差异(图7-A, B, D); 相反用JA处理的植株雄穗性别得到了恢复(图7-C), 花丝数量明显减少, 平均雄穗结实29粒, 与未实施任何处理和用0.05%乙醇处理植株的雄穗结实数有极显著差异。同时在田间发现JA处理植株性别恢复程度并不完全一致, 部分植株雌性化小穗恢复程度较低, 但也有部分植株99%雌性化小穗得到恢复。该结果证实ts12基因参与JA合成途径, 从而影响了玉米性别决定过程。图7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7茉莉酸处理后ts12的表型及雄穗结实数
A: 未处理的ts12雄穗表型; B: 0.05%乙醇处理的ts12雄穗表型; C: JA处理的ts12雄穗表型; D: 雄穗结实数统计; 柱形图上方数字代表标准差; **代表JA处理植株雄穗结实数量与未处理植株和0.05%乙醇处理植株雄穗结实数在统计学上有极显著差异(P < 0.01)。
Fig. 7Phenotype of ts12 treated by JA and tassel seeds statistics
A: untreated ts12 tassel phenotype; B: 0.05% ethanol-treated ts12 tassel phenotype; C: JA-treated ts12 tassel phenotype; D: number of tassel seeds per ts12 mutant. The numbers on top of the column represent the standard deviation. ** indicates extremely significant difference of tassel seeds number between JA-treated ts12 and untreated and 0.05% ethanol-treated ts12 plants (P < 0.01).
2.6 ts2基因表达分析
基因表达水平是一个基因在特定组织中功能重要与否的重要评判依据。提取F2群体中ts12突变体和野生型植株各组织总RNA, 对Ts2基因表达进行定量分析(图8)。发现Ts2基因在玉米雄穗中表达量最高, 雌穗和叶片中次之, 而在根、茎、花丝中的表达水平较低, 在授粉后20 d的胚乳和胚中没有检测到表达, 由此可知Ts2基因主要在玉米雄穗中发挥重要作用。同时在ts12突变体中, Ts2基因突变后, 其表达在未成熟雄穗和雌穗中极显著降低, 从而可能导致雄穗中的雌蕊不能正常败育, 最终产生tasselseed的表型。图8
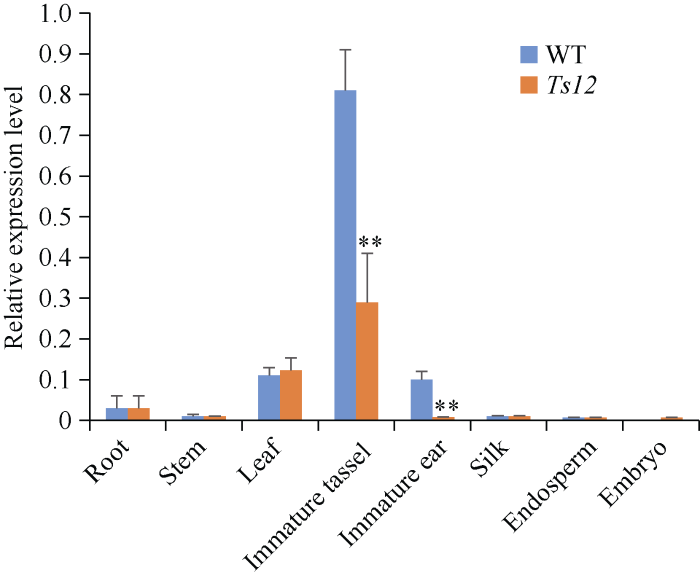 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8Ts2基因表达分析
Root: 根; Stem: 茎; Leaf: 叶; Silk: 花丝; Immature ear: 雌穗; Immature tassel: 雄穗; Embryo: 授粉后20 d胚; Endosperm: 授粉后20 d胚乳。**代表在特定组织中Ts2基因表达与野生型相比在统计学上有极显著差异(P < 0.01)。
Fig. 8Expression analysis of Ts2 gene
** indicates extremely significant difference of expression in specific tissues between Ts2 and WT (P < 0.01).
3 讨论
玉米雄穗和雌穗发育与玉米产量密切相关。玉米雄穗小花中的雌蕊、雌穗小花中的雄蕊的选择性败育产生了单性花, 这一过程是多个基因表达调控网络及遗传位点控制的。目前, 已有多个控制玉米性别的遗传位点被鉴定[25,26]。我们通过化学诱变获得了一个tasselseed表型突变体ts12, 利用图位克隆的方法分离目标基因, 发现已知基因Ts2可能就是导致ts12突变表型的原因。克隆分析Ts2基因发现, 其编码蛋白保守结构域中发生了氨基酸的替换。等位性测验结果表明ts12突变表型是已报道的性别决定基因Ts2的突变引起的(图7-A)。外源施加茉莉酸处理ts12后, 性别明显得到恢复; 杨松青等[27]研究也揭示了Ts2表达受到JA的诱导, 进一步证实了Ts2是一个典型的依赖于JA的性别决定基因。Ts2基因编码一个短链乙醇脱氢酶。短链乙醇脱氢酶超家族蛋白是一类与系统发育相关的酶, 催化作用于多种底物, 如糖、类固醇、前列腺素、芳香烃、抗生素和参与氮代谢的化合物[28]。短链乙醇脱氢酶超家族蛋白具有一个保守的结构域A, 它可以与NAD(P)辅助因子结合[29]。通过比较高粱、小米、水稻、二穗短柄草、拟南芥的Ts2直系同源蛋白及小鼠[30]、人类[31]、黄杆菌[32]、假单胞菌[33]和大肠肝菌[34]短链乙醇脱氢酶发现, 在其保守结构域A中具有典型的短链脱氢酶基序特征“TGxxxGhG”(附图1), 该基序在与辅酶结合过程中具有重要作用[35,36]。在ts12突变体中, Ts2蛋白的第66个氨基酸, 也即基序TGxxxGhG中的第2个G (甘氨酸)被替换为R (精氨酸), 可能导致Ts2无法与辅酶稳定结合, Ts2蛋白复合体催化效率降低。ts12突变体雄穗主穗及侧枝顶端部分雄蕊可正常发育, 产生有功能的花粉, 可自交获得纯合的突变体后代。然而ts2突变表型为雄穗小穗完全雌性化, 仅能产生雌性的单性花, 而且雌性化的小穗均能产生正常功能的雌配子[5]。由于ts2突变体是由Ac转座子插入引起的, 导致基因功能完全丧失; 而ts12突变体则是EMS诱变导致Ts2基因CDS序列发生单核苷酸突变, 除了该基因编码蛋白第66个氨基酸发生了替换外, 其他氨基酸序列与野生型一致。因此, 我们推测ts12突变体中, 单个氨基酸的替换虽然导致Ts2蛋白不能与辅酶稳定结合, 催化效率降低, 但仍能发挥部分功能, 致使ts12部分小穗可正常散粉。ts2与ts12突变体测验后代与ts12一样可产生有功能的花粉, 进一步印证了这一论断。但同时我们也注意到在ts12突变体中, Ts2基因在雄穗和雌穗中的表达水平与正常植株相比, 显著降低(图8), 这一因素可能也是造成性别决定发生反转的原因之一。通过对Ts2基因转录起始位点上游包括启动子区域的3 kb序列进行测序分析, 没有发现碱基的突变, 因此推测Ts2表达水平的改变可能是因单位点突变后造成mRNA不稳定, 也可能是基因表达调控网络反馈抑制的结果。
附图1
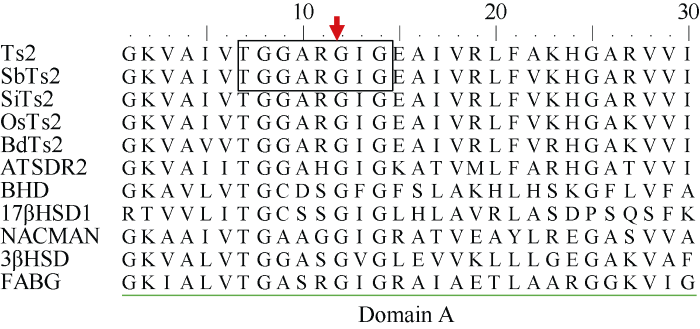 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT附图1短链乙醇脱氢酶同源蛋白保守结构域A序列分析
SbTs2: 高粱Ts2; SiTs2: 小米Ts2; OsTs2: 水稻Ts2; BdTs2: 二穗短柄草Ts2; ATSDR2: 拟南芥Ts2直系同源蛋白; BHD: 来自小鼠的短链乙醇脱氢酶; 17βHSD1: 来自人的短链乙醇脱氢酶; NACMAN: 来自黄杆菌的短链乙醇脱氢酶; 3βHSD: 来自假单胞菌的短链乙醇脱氢酶; FABG: 来自大肠杆菌的短链乙醇脱氢酶。
Supplementary fig. 1Sequence analysis of conservative domain A of short-chain alcohol dehydrogenases
SbTs2: Ts2 ortholog from sorghum; SiTs2: Ts2 ortholog from Setaria italic; OsTs2: Ts2 ortholog from rice; BdTs2: Ts2 ortholog from Brachypodium distachyon; ATSDR2: Ts2 ortholog from Arabidopsis thaliana; BHD: a short-chain alcohol dehydrogenase from rat; 17βHSD1: a short-chain alcohol dehydrogenase from human; NACMAN: a short-chain alcohol dehydrogenase from Flavobacterium; 3βHSD: a short-chain alcohol dehydrogenase from Pseudomonas testosteroni; FABG: a short-chain alcohol dehydrogenase from Escherichia coli.
玉米性别决定过程中, 雌蕊的败育依赖于植物激素茉莉酸调节的基因表达调控网络。外源施加茉莉酸可恢复ts1、ts2和Ts5突变体表型[4,8], 因此这3个基因均参与了茉莉酸的合成和代谢。ts1、ts5双突变体和ts2、ts5双突变体表型分析表明ts1和ts2与Ts5均呈上位性效应[37], 而ts2 mRNA在雌蕊中的积累依赖于ts1的生物学功能[6], 因此ts2位于ts1茉莉酸代谢合成基因表达调控网络的下游节点。Wu等[7]在大肠杆菌中制备了TS2重组蛋白, 发现TS2以四聚体形式发挥功能, 起3β-或17β-羟基类固醇脱氢酶或羰基/醌还原酶的作用。然而其在玉米性别决定过程中发挥重要功能的具体作用机制依然不是十分清楚。我们新发现的ts2等位突变体ts12丰富了与玉米性别决定研究材料, 为进一步解析Ts2基因在玉米性别决定过程中的分子机制奠定了基础。
4 结论
通过EMS诱变, 新发现了一个与玉米性别决定相关的突变体ts12。ts12被定位于玉米1号染色体1.04 bin 290 kb的物理区间。Ts2基因CDS中的第196个碱基鸟嘌呤被替换为腺嘌呤, 导致该位点编码的氨基酸甘氨酸被替换为精氨酸。ts12是ts2的等位突变体。ts12突变体为深入探究Ts2基因的功能提供了新的研究材料。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
DOI:10.3732/ajb.94.11.1745URLPMID:21636370 [本文引用: 1]

Axillary meristems play a fundamental role in inflorescence architecture. Maize (Zea mays) inflorescences are highly branched panicles because of the production of multiple types of axillary meristems. We used auxin transport inhibitors to show that auxin transport is required for axillary meristem initiation in the maize inflorescence. The phenotype of plants treated with auxin transport inhibitors is very similar to that of barren inflorescence2 (bif2) and barren stalk1 (ba1) mutants, suggesting that these genes function in the same auxin transport pathway. To dissect this pathway, we performed RNA in situ hybridization on plants treated with auxin transport inhibitors. We determined that bif2 is expressed upstream and that ba1 is expressed downstream of auxin transport, enabling us to integrate the genetic and hormonal control of axillary meristem initiation. In addition, treatment of maize inflorescences with auxin transport inhibitors later in development results in the production of single instead of paired spikelets. Paired spikelets are a key feature of the Andropogoneae, a group of over 1000 grasses that includes maize, sorghum, and sugarcane. Because all other grasses bear spikelets singly, these results implicate auxin transport in the evolution of inflorescence architecture. Furthermore, our results provide insight into mechanisms of inflorescence branching that are relevant to all plants.
DOI:10.1126/science.7985019URLPMID:7985019 [本文引用: 1]
Maize partitions the sexes into different flowers on the plant, a condition called monoecy, which facilitates outcrossing. Sex determination in maize is a complex process involving an interplay between genetic determinants, the environment, and hormones. Unisexuality of flowers is achieved by the process of selective arrest and abortion of the inappropriate organ primordia within a bisexual floral meristem. Floral organ abortion is associated with the degeneration of cells within an immature primordia. Masculinizing genes are required for gynoecial abortion, feminizing genes arrest stamen development, and both types also control secondary sexual traits involving morphological characteristics of floral tissues. Gibberellins, steroid-like plant hormones, appear to play a pivotal role in the stamen abortion process and the feminization of floral tissues.
DOI:10.1126/science.1164645URLPMID:19131630 [本文引用: 4]
Sex determination in maize is controlled by a developmental cascade leading to the formation of unisexual florets derived from an initially bisexual floral meristem. Abortion of pistil primordia in staminate florets is controlled by a tasselseed-mediated cell death process. We positionally cloned and characterized the function of the sex determination gene tasselseed1 (ts1). The TS1 protein encodes a plastid-targeted lipoxygenase with predicted 13-lipoxygenase specificity, which suggests that TS1 may be involved in the biosynthesis of the plant hormone jasmonic acid. In the absence of a functional ts1 gene, lipoxygenase activity was missing and endogenous jasmonic acid concentrations were reduced in developing inflorescences. Application of jasmonic acid to developing inflorescences rescued stamen development in mutant ts1 and ts2 inflorescences, revealing a role for jasmonic acid in male flower development in maize.
DOI:10.1016/0092-8674(93)90522-rURLPMID:8358795 [本文引用: 2]
Maize produces separate unisexual flowers through programmed abortion of preformed organ primordia. In the male inflorescence (tassel), stamen primordia develop to sexual maturity, while gynoecia (pistil primordia) are aborted. In tasselseed2 (ts2) mutant plants, floral structures in the tassel adopt a female developmental program. Here we report the transposon tagging and cloning of the TS2 gene, which plays a late but pivotal role in determining the sexual fate of floral meristems. Shortly before abortion of the gynoecium, Ts2 mRNA is expressed subepidermally in that primordium. Phenotypic instability of the Activator (Ac)-induced allele ts2-m1 indicates that late restoration of TS2 action in somatic tissues, which is correlated with Ac excision, reactivates the male developmental program. The predicted amino acid sequence of the Ts2 protein shows significant similarity to short-chain alcohol dehydrogenases, particularly hydroxysteroid dehydrogenases.
URLPMID:9876173 [本文引用: 2]
The formation of unisexual flowers in maize requires the selective elimination and sexual maturation of floral organs in an initially bisexual floral meristem. Elimination of pistil primordia occurs in the primary and secondary florets of the tassel spikelets, and in the secondary florets of ear spikelets. Ill-fated pistil cells undergo a cell death process associated with nuclear degeneration in a specific spatial-temporal pattern that begins in the subepidermis, eventually aborting the entire organ. The sex determination genes tasselseed1 and tasselseed2 are required for death of pistil cells. tasselseed1 is required for the accumulation of TASSELSEED2 mRNA in pistil cells. All pistil primordia express TASSELSEED2 RNA but functional pistils found in ear spikelets are protected from cell death by the action of the silkless1 gene. silkless1 blocks tasselseed-induced cell death in the pistil primordia of primary ear florets. A model is proposed for the control of pistil fate by the action of the ts1-ts2-sk1 pathway.
DOI:10.1111/j.1742-4658.2007.05642.xURLPMID:17298439 [本文引用: 2]
The development of unisexual flowers in maize and other plants proceeds through selective elimination of floral organs in an initially bisexual floral meristem. The essential character of the tasselseed 2 gene (TS2) in this cell-death pathway has been established previously. Molecular cloning of TS2 reveals membership to the evolutionarily conserved superfamily of short-chain dehydrogenases/reductases, but its substrate specificity remained unknown. Recombinant TS2 protein was produced in Escherichia coli, and purified to apparent homogeneity. Analytical ultracentrifugation and gel filtration experiments show that TS2 is a tetrameric enzyme. Thermal denaturation followed by circular dichroism spectroscopy reveals that TS2 binds NAD(H) and NAD(P)(H). Substrate screening demonstrates that TS2 converts steroids with specificities found at positions 3 and 17, and several dicarbonyl and quinone compounds, thus establishing TS2 as a plant 3beta/17beta-hydroxysteroid dehydrogenase and carbonyl/quinone reductase. Taken together, the genetic data and the substrate specificities determined suggest that TS2 converts specific plant compounds and acts as a prereceptor control mechanism, in a manner similar to that of mammalian hydroxysteroid dehydrogenases.
.
DOI:10.1038/s42003-019-0354-1URLPMID:30937397 [本文引用: 3]
Maize is monecious, with separate male and female inflorescences. Maize flowers are initially bisexual but achieve separate sexual identities through organ arrest. Loss-of-function mutants in the jasmonic acid (JA) pathway have only female flowers due to failure to abort silks in the tassel. Tasselseed5 (Ts5) shares this phenotype but is dominant. Positional cloning and transcriptomics of tassels identified an ectopically expressed gene in the CYP94B subfamily, Ts5 (ZmCYP94B1). CYP94B enzymes are wound inducible and inactivate bioactive jasmonoyl-L-isoleucine (JA-Ile). Consistent with this result, tassels and wounded leaves of Ts5 mutants displayed lower JA and JA-lle precursors and higher 12OH-JA-lle product than the wild type. Furthermore, many wounding and jasmonate pathway genes were differentially expressed in Ts5 tassels. We propose that the Ts5 phenotype results from the interruption of JA signaling during sexual differentiation via the upregulation of ZmCYP94B1 and that its proper expression maintains maize monoecy.
DOI:10.1126/sciadv.1600991URLPMID:27819048 [本文引用: 1]
Sex determination in maize involves the production of staminate and pistillate florets from an initially bisexual floral meristem. Pistil elimination in staminate florets requires jasmonic acid signaling, and functional pistils are protected by the action of the silkless 1 (sk1) gene. The sk1 gene was identified and found to encode a previously uncharacterized family 1 uridine diphosphate glycosyltransferase that localized to the plant peroxisomes. Constitutive expression of an sk1 transgene protected all pistils in the plant, causing complete feminization, a gain-of-function phenotype that operates by blocking the accumulation of jasmonates. The segregation of an sk1 transgene was used to effectively control the production of pistillate and staminate inflorescences in maize plants.
DOI:10.1007/s00425-016-2607-2URLPMID:27770199 [本文引用: 1]
The determining process of pistil fate are central to maize sex determination, mainly regulated by a genetic network in which the sex-determining genes SILKLESS 1 , TASSEL SEED 1 , TASSEL SEED 2 and the paramutagenic locus Required to maintain repression 6 play pivotal roles. Maize silks, which emerge from the?ear shoot and derived from the?pistil, are the functional stigmas of female flowers and play a pivotal role in pollination. Previous studies on sex-related mutants have revealed that sex-determining genes and phytohormones play an important role in the regulation of flower organogenesis. The processes determining pistil fate are central to flower development, where a silk identified gene SILKLESS 1 (SK1) is required to protect pistil primordia from a cell death signal produced by two commonly known genes, TASSEL SEED 1 (TS1) and TASSEL SEED 2 (TS2). In this review, maize flower developmental process is presented together with a focus on important sex-determining mutants and hormonal signaling affecting pistil development. The role of sex-determining genes, microRNAs, phytohormones, and the paramutagenic locus Required to maintain repression 6 (Rmr6), in forming a regulatory network that determines pistil fate, is discussed. Cloning SK1 and clarifying its function were crucial in understanding the regulation network of sex determination. The signaling mechanisms of phytohormones in sex determination are also an important research focus.
DOI:10.1016/j.ydbio.2007.06.004URLPMID:17612519 [本文引用: 1]
In maize (Zea mays ssp. mays), the meiotically heritable maintenance of specific transcriptionally repressed epigenetic states is facilitated by a putative RNA-dependent RNA polymerase encoded by mediator of paramutation1 (mop1) and an unknown factor encoded by the required to maintain repression6 (rmr6) locus. These so-called &quot;paramutant&quot; states occur at certain alleles of loci encoding regulators of anthocyanin pigment biosynthesis. Here we show Rmr6 acts to canalize leaf and inflorescence development by prohibiting the ectopic action of key developmental regulators. Phenotypic and genetic analyses suggest that Rmr6 ensures proper adaxial-abaxial polarity of the leaf sheath by limiting the expression domain of a putative adaxializing factor. Similar tests indicate that Rmr6 maintains maize's monoecious pattern of sex determination by restricting the function of the pistil-protecting factor, silkless1, from the apical inflorescence. Phenotypic similarities with mop1 mutant plants together with current models of heterochromatin maintenance and leaf polarity imply Rmr6 functions to maintain epigenetic repression established by non-coding small RNA molecules.
DOI:10.1105/tpc.111.094151URLPMID:22523204 [本文引用: 1]
Here, multiple functions of jasmonic acid (JA) in maize (Zea mays) are revealed by comprehensive analyses of JA-deficient mutants of the two oxo-phytodienoate reductase genes, OPR7 and OPR8. Single mutants produce wild-type levels of JA in most tissues, but the double mutant opr7 opr8 has dramatically reduced JA in all organs tested. opr7 opr8 displayed strong developmental defects, including formation of a feminized tassel, initiation of female reproductive buds at each node, and extreme elongation of ear shanks; these defects were rescued by exogenous JA. These data provide evidence that JA is required for male sex determination and suppression of female reproductive organ biogenesis. Moreover, opr7 opr8 exhibited delayed leaf senescence accompanied by reduced ethylene and abscisic acid levels and lack of anthocyanin pigmentation of brace roots. Remarkably, opr7 opr8 is nonviable in nonsterile soil and under field conditions due to extreme susceptibility to a root-rotting oomycete (Pythium spp), demonstrating that these genes are necessary for maize survival in nature. Supporting the importance of JA in insect defense, opr7 opr8 is susceptible to beet armyworm. Overall, this study provides strong genetic evidence for the global roles of JA in maize development and immunity to pathogens and insects.
DOI:10.4161/15592316.2014.970442URLPMID:25482807 [本文引用: 1]
It is well-known from the model dicotyledonous plants, Arabidopsis and tomato, that jasmonates (JAs) act as defense hormones in planta due to their potent ability to mediate defensive responses against insect/pathogen attacks or harsh environmental conditions. JA is also required for various developmental processes such as male fertility, seed maturation, root extension, and leaf senescence. In our recently published Plant Cell paper, the multiple roles of JA in the monocotyledonous agro-economically important model plant, maize, were investigated by comprehensive analysis of JA-deficient double mutant disrupted in the two oxophytodienoate reductase genes, OPR7 and OPR8. These two genes are the closest orthologs of the Arabidopsis JA-producing OPR3 and are the only maize OPRs required for JA biosynthesis. With this mutant, we previously showed that JA is essential for both male and female reproductive development, and required for the regulation of brace root pigmentation, leaf senescence, and defense against oomycete Pythium aristosporum, and beet armyworm (Spodoptera exigua). In this addendum, we expanded the investigation into the function of JA in elongation of sheaths, leaves, and roots, and its involvement in photomorphogenesis of seedlings.
DOI:10.3724/SP.J.1259.2012.00065URL [本文引用: 1]
玉米(Zea mays)属典型的雌雄异花植物, 单性花的形成经历了复杂的性别决定过程。通过雄穗小花和雌穗下位花的雌蕊原基以及雌穗小花雄蕊原基的选择性败育(或退化), 玉米最终形成正常的雌雄同株单性花。相关突变体的研究揭示, 玉米性别决定涉及选择性细胞死亡、细胞保护及信号转导等复杂的过程。其中, 植物激素信号的调控在玉米性别决定过程中处于核心地位。最近的研究表明, 赤霉素、细胞分裂素和茉莉酸类物质参与调控玉米性别决定过程。该文结合最新研究成果, 综述了植物激素在玉米性别决定中的作用及其调控途径, 同时提出了研究中存在的问题, 并对该领域未来的研究方向进行了展望。
DOI:10.3724/SP.J.1259.2012.00065URL [本文引用: 1]
玉米(Zea mays)属典型的雌雄异花植物, 单性花的形成经历了复杂的性别决定过程。通过雄穗小花和雌穗下位花的雌蕊原基以及雌穗小花雄蕊原基的选择性败育(或退化), 玉米最终形成正常的雌雄同株单性花。相关突变体的研究揭示, 玉米性别决定涉及选择性细胞死亡、细胞保护及信号转导等复杂的过程。其中, 植物激素信号的调控在玉米性别决定过程中处于核心地位。最近的研究表明, 赤霉素、细胞分裂素和茉莉酸类物质参与调控玉米性别决定过程。该文结合最新研究成果, 综述了植物激素在玉米性别决定中的作用及其调控途径, 同时提出了研究中存在的问题, 并对该领域未来的研究方向进行了展望。
DOI:10.1104/pp.88.4.1367URLPMID:16666468 [本文引用: 1]
Gibberellins A(12) (GA(12)), GA(53), GA(44), GA(19), GA(17), GA(20), GA(29), GA(1), and GA(8) have been identified from extracts of vegetative shoots of normal (wild type) maize using full scan capillary gas chromatography-mass spectrometry and Kovats retention indices. Seven of these gibberellins (GAs) have been quantified by capillary gas chromatography-selected ion monitoring using internal standards of [(14)C(4)]GA(53), [(14)C(4)]GA(44), [(2)H(2)] GA(19), [(13)C(1)]GA(20), [(13)C(1)]GA(29), [(13)C(1)]GA(1), and [(13)C(1)]GA(8). Quantitative data from extracts of normal, dwarf-1, dwarf-2, dwarf-3, and dwarf-5 seedlings support the operation of the early 13-hydroxylation pathway in vegetative shoots of Zea mays. These data support the positions in the pathway blocked by the mutants, previously assigned by bioassay data and metabolic studies. The GA levels in dwarf-2, dwarf-3, and dwarf-5 were equal to, or less than, 2.0 nanograms per 100 grams fresh weight, showing that these mutants are blocked for steps early in the pathway. In dwarf-1, the level of GA(1) was very low (0.23 nanograms per 100 grams fresh weight) and less than 2% of that in normal shoots, while GA(20) and GA(29) accumulated to levels over 10 times those in normals; these results confirm that the dwarf-1 mutant blocks the conversion of GA(20) to GA(1). Since the level of GAs beyond the blocked step for each mutant is greater than zero, each mutated gene probably codes for an altered gene product, thus leading to impaired enzyme activities.
DOI:10.1105/tpc.7.1.75URLPMID:7696880 [本文引用: 1]
The Anther ear1 (An1) gene product is involved in the synthesis of ent-kaurene, the first tetracyclic intermediate in the gibberellin (GA) biosynthetic pathway. Mutations causing the loss of An1 function result in a GA-responsive phenotype that includes reduced plant height, delayed maturity, and development of perfect flowers on normally pistillate ears. The an1::Mu2-891339 allele was recovered from a Mutator (Mu) F2 family. Using Mu elements as molecular probes, an An1-containing restriction fragment was identified and cloned. The identity of the cloned gene as An1 was confirmed by using a reverse genetics screen for maize families that contain a Mu element inserted into the cloned gene and then by demonstrating that the insertion causes an an1 phenotype. The predicted amino acid sequence of the An1 cDNA shares homology with plant cyclases and contains a basic N-terminal sequence that may target the An1 gene product to the chloroplast. The sequence is consistent with the predicted subcellular localization of AN1 in the chloroplast and with its biochemical role in the cyclization of geranylgeranyl pyrophosphate, a 20-carbon isoprenoid, to ent-kaurene. The semidwarfed stature of an1 mutants is in contrast with the more severely dwarfed stature of GA-responsive mutants at other loci in maize and may be caused by redundancy in this step of the GA biosynthetic pathway. DNA gel blot analysis indicated that An1 is a single-copy gene that lies entirely within the deletion of the an1-bz2-6923 mutant. However, homozygous deletion mutants accumulated ent-kaurene to 20% of the wild-type level, suggesting that the function of An1 is supplemented by an additional activity.
DOI:10.1016/S0083-6729(05)72003-3URLPMID:16492469 [本文引用: 1]
DOI:10.2135/cropsci1976.0011183X001600030013xURL [本文引用: 1]
DOI:10.1073/pnas.1108359108URLPMID:22106275 [本文引用: 1]
Brassinosteroids (BRs) are plant hormones that regulate growth and development. They share structural similarities with animal steroids, which are decisive factors of sex determination. BRs are known to regulate morphogenesis and environmental stress responses, but their involvement in sex determination in plants has been only speculative. We show that BRs control sex determination in maize revealed through characterization of the classical dwarf mutant nana plant1 (na1), which also feminizes male flowers. na1 plants carry a loss-of-function mutation in a DET2 homolog--a gene in the BR biosynthetic pathway. The mutant accumulates the DET2-specific substrate (24R)-24-methylcholest-4-en-3-one with a concomitant decrease of downstream BR metabolites. Treatment of wild-type maize plants with BR biosynthesis inhibitors completely mimicked both dwarf and tasselseed phenotypes of na1 mutants. Tissue-specific na1 expression in anthers throughout their development supports the hypothesis that BRs promote masculinity of the male inflorescence. These findings suggest that, in the monoecious plant maize, BRs have been coopted to perform a sex determination function not found in plants with bisexual flowers.
DOI:10.1038/ng.2007.20URLPMID:18026103 [本文引用: 1]
In maize (Zea mays), sex determination occurs through abortion of female carpels in the tassel and arrest of male stamens in the ear. The Tasselseed6 (Ts6) and tasselseed4 (ts4) mutations permit carpel development in the tassel while increasing meristem branching, showing that sex determination and acquisition of meristem fate share a common pathway. We show that ts4 encodes a mir172 microRNA that targets APETALA2 floral homeotic transcription factors. Three lines of evidence suggest that indeterminate spikelet1 (ids1), an APETALA2 gene required for spikelet meristem determinacy, is a key target of ts4. First, loss of ids1 suppresses the ts4 sex determination and branching defects. Second, Ts6 mutants phenocopy ts4 and possess mutations in the microRNA binding site of ids1. Finally, IDS1 protein is expressed more broadly in ts4 mutants compared to wild type. Our results demonstrate that sexual identity in maize is acquired by limiting floral growth through negative regulation of the floral homeotic pathway.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s10681-018-2132-4URL [本文引用: 1]
DOI:10.1105/tpc.5.10.1205URLPMID:12271023 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/0960-0760(94)90084-1URLPMID:7981120 [本文引用: 1]
Multiple alignment of members of the short-chain alcohol dehydrogenase (SCAD) superfamily, according to the conserved domains A-F, has revealed a number of important relationships. It can be shown that the 17 beta-hydroxysteroid dehydrogenase type 2 enzyme is more closely related to D-beta-hydroxybutyrate dehydrogenase than it is to 17 beta-hydroxysteroid dehydrogenase type 1. Carbonyl reductase, previously considered to be a member of the aldo-keto reductase superfamily, displayed high homology in the conserved domains and is clearly part of the SCAD superfamily despite the insertion of a large peptide between conserved domains. Alignment of the product of the Leishmania methotrexate resistance gene HMTX showed that an internal, highly conserved domain can be substituted by an unrelated sequence without loss of biological activity. Furthermore, comparisons of the chimeric trifunctional enzyme enoyl-CoA hydratase/3-hydroxyacyl-CoA dehydrogenase/3-hydroxyacyl-CoA epimerase with other family members suggests that the region between the conserved B and C domains is the last to diverge between closely related enzymes and that the F domain appears to evolve with a different evolutionary clock to the rest of the protein. Finally, a highly conserved pattern of serine and threonine residues in the active site of SCAD enzymes indicates that these residues may play an important role in catalysis. These observations should facilitate alignment of future members of the SCAD superfamily.
DOI:10.1128/jb.170.5.2070-2077.1988URLPMID:2834320 [本文引用: 1]
Eubacterium sp. strain VPI 12708 is an anaerobic intestinal bacterium that has inducible bile acid 7-dehydroxylation activity. At least four new polypeptides were synthesized after addition of primary bile acids to the growth medium. One of these, of molecular weight 27,000 (P-27), was shown to be involved in the 7-dehydroxylation reaction sequence. The gene coding for P-27 was cloned, and the entire DNA sequence for the protein-coding region was determined. In addition, sequence information was obtained for 294 bases upstream from the translational start codon and 329 bases downstream from the stop codon. Induction studies with a synthetic oligonucleotide probe (16-mer) revealed the presence of a cholic acid-inducible mRNA species approximately 900 bases long. A 5' terminus of this mRNA was detected by primer extension analysis, and the location of the 3' terminus of the mRNA was estimated by using S1 nuclease mapping. The 3' terminus of the mRNA contained a large element with dyad symmetry of unknown function. The open reading frame contained 249 codons, and the corresponding polypeptide had a calculated molecular weight of 26,745. The amino acid sequence of P-27 showed significant homology to several previously described alcohol-polyol dehydrogenases (&quot;nonzinc&quot; dehydrogenases), especially in the region believed to contain a pyridine nucleotide-binding domain. The implications of this homology and the possible function of P-27 in bile acid 7-dehydroxylation are discussed.
DOI:10.1021/bi00130a009URLPMID:1567834 [本文引用: 1]
The amino acid sequence of D-beta-hydroxybutyrate dehydrogenase (BDH), a phosphatidyl-choline-dependent enzyme, has been determined for the enzyme from rat liver by a combination of nucleotide sequencing of cDNA clones and amino acid sequencing of the purified protein. This represents the first report of the primary structure of this enzyme. The largest clone contained 1435 base pairs and encoded the entire amino acid sequence of mature BDH and the leader peptide of precursor BDH. Hybridization of poly(A+) rat liver mRNA revealed two bands with estimated sizes of 3.2 and 1.7 kb. A computer-based comparison of the amino acid sequence of BDH with other reported sequences reveals a homology with the superfamily of short-chain alcohol dehydrogenases, which are distinct from the classical zinc-dependent alcohol dehydrogenases. This protein family, initially discerned from Drosophila alcohol dehydrogenase and bacterial ribitol dehydrogenase, is now known to include at least 20 enzymes catalyzing oxidations of distinct substrates.
DOI:10.1016/0014-5793(88)80548-9URLPMID:2846351 [本文引用: 1]
cDNA clones for 17 beta-hydroxysteroid dehydrogenase (17-HSD; EC 1.1.1.62) were isolated from a placental lambda gt11 expression library using polyclonal antibodies against placental 17-HSD. The largest cDNA contained 1325 nucleotides, consisting of a short 5'-noncoding segment, a coding segment of 987 nucleotides terminated by a TAA codon, and a 329 nucleotide long 3'-noncoding segment. The open reading frame encoded a polypeptide of 327 amino acid residues with a predicted Mr of 34853. The amino acid sequence of 23 N-terminal amino acids determined from purified 17-HSD agreed with the sequence deduced from cDNA. The deduced amino acid sequence also contained two peptides previously characterized from the proposed catalytic area of placental 17-HSD.
URLPMID:1854199 [本文引用: 1]
The gene coding for N-acyl-D-mannosamine dehydrogenase (NAM-DH) from Flavobacterium sp. strain 141-8 was cloned and expressed under the control of a lac promoter in Escherichia coli JM109. The DNA sequence of the gene was determined, and an open reading frame encoding a polypeptide composed of 272 amino acid residues (Mr, 27,473) was identified. The E. coli transformants which showed over 200-fold higher NAM-DH activity than did the Flavobacterium strain produced the enzyme as a protein fused with beta-galactosidase. Despite being a fusion, NAM-DH produced by E. coli transformants appeared unchanged in pH optimum, Km, and substrate specificity from Flavobacterium sp. strain 141-8. This newly recombinant enzyme may be applicable to the quantitative determination of sialic acid in serum.
DOI:10.1016/0960-0760(93)90020-wURLPMID:8382516 [本文引用: 1]
We describe the cloning, sequencing and overexpression of the (3-17)beta hydroxysteroid dehydrogenase gene of Pseudomonas testosteroni. A genomic library of Ps. testosteroni total DNA constructed from SauIIIA digests ligated to a lambda gt11 vector was probed with polyclonal antibody raised against purified enzyme. Subclones derived from a recombinant phage containing a 2661-base-pair insert were sequenced and found to contain an open reading frame of 765 base pairs that corresponds to a protein of 254 amino acid residues. A 1492-base-pair fragment was inserted into pBR322 plasmid vector and used to construct a strain of E. coli HB101 that overexpressed the steroid dehydrogenase gene.
URLPMID:1556094 [本文引用: 1]
The gene encoding Escherichia coli acyl carrier protein (ACP) has been isolated and sequenced. The ACP gene (called acpP) was located on the genetic map between fabF and fabD which encode two fatty acid biosynthetic enzymes, 3-ketoacyl-ACP synthase II and malonyl CoA-ACP transacylase, respectively. An open reading frame between acpP and fabD encodes a 26.5-kDa protein that has significant sequence identity (greater than 40%) with two acetoacetyl-CoA reductases and thus is believed to encode a 3-ketoacyl-ACP reductase. This gene (called fabG) is cotranscribed with acpP. Thus, the gene encoding ACP, the key carrier protein of fatty acid synthesis, is located within a cluster of fatty acid biosynthetic genes.
DOI:10.1021/bi961803vURLPMID:8993315 [本文引用: 1]
Mutagenetic replacements of conserved residues within the active site of the short-chain dehydrogenase/reductase (SDR) superfamily were studied using prokaryotic 3 beta/17 beta-hydroxysteroid dehydrogenase (3 beta/17 beta-HSD) from Comamonas testosteroni as a model system. The results provide novel data to establish Ser 138 as a member of a catalytically important &quot;triad&quot; of residues also involving Tyr151 and Lys155. A Ser--&gt;Ala exchange at position 138 results in an almost complete (&gt; 99.9%) loss of enzymatic activity, which is not observed with a Ser--&gt;Thr replacement. This indicates that an essential factor for catalysis is the ability of side chain 138 to form hydrogen bond interactions. Mutations in the NAD(H) binding region, in strands beta A, beta D, and adjacent turns, reveal two additional residues, Thr12 and Asn87, which are important for correct binding of the coenzyme and with a differential effect on the reactions catalyzed. Thus, mutation of Thr12 to Ala results in a complete loss of the 3 beta-dehydrogenase activity, whereas the 3-oxoreductase activity remains unchanged. On the other hand, a T12S substitution yields a protein with unaltered catalytic constants for both reactions, revealing that a specific hydrogen bond is critical for the dehydrogenase activity. Our interpretation of the available crystal structure of 3 alpha/20 beta-HSD from Streptomyces hydrogenans suggests a hydrogen bond in that enzyme between the Thr12 side chain and the backbone NH of Asn87 rather than the coenzyme, indicating that this hydrogen bond to the beta D strand might determine a crucial difference between the reductive and the oxidative reaction types. Similarly, mutation of Asn87 to Ala results in an 80% reduction of kcat/Km in the dehydrogenase direction but also unchanged 3-oxoreductase properties. It appears that the binding of NAD+ to the protein is influenced by local structural changes involving strand beta D and turn beta A to alpha B.
DOI:10.1016/s0014-5793(99)00130-1URLPMID:10094468 [本文引用: 1]
Short-chain dehydrogenases/reductases (SDR) and medium-chain dehydrogenases/reductases (MDR) are protein families originally distinguished from characterisations of alcohol dehydrogenase of these two types. Screening of completed genome sequences now reveals that both these families are large, wide-spread and complex. In Escherichia coli alone, there are no fewer than 17 MDR forms, identified as open reading frames, considerably extending previously known MDR relationships in prokaryotes and including ethanol-active alcohol dehydrogenase. In entire databanks, 1056 SDR and 537 MDR forms are currently known, extending the multiplicity further. Complexity is also large, with several enzyme activity types, subgroups and evolutionary patterns. Repeated duplications can be traced for the alcohol dehydrogenases, with independent enzymogenesis of ethanol activity, showing a general importance of this enzyme activity.
DOI:10.1016/s0736-5748(97)00001-4URLPMID:9178034 [本文引用: 1]
We studied the effects of chronic K(+)-induced membrane depolarization and treatment with N-methyl-D-aspartate (NMDA) on cerebellar granule cells (CGCs) from weaver mutant mice and non-weaver litter-mates. The weaver mutation is a Gly-to-Ser substitution in a conserved region of the Girk2 G protein-coupled inward rectifying potassium channel [Patil N., Cox D. R., Bhat D., Faham M., Myers R. M. and Peterson A. S. (1995) Nature Genet. 11, 126-129] which induces early death of CGCs. The biochemical differentiation of CGCs was estimated as the rate of 2-deoxy-D-glucose accumulation and the expression of neural cell adhesion molecule (NCAM). High (25 mM) K+ ion concentration or treatment with NMDA greatly promoted the biochemical differentiation of both weaver mutant and non-weaver litter-mate mouse CGCs. In contrast to the marked effect on biochemical differentiation in both weaver and non-weaver mice CGSs, chronic high K+ treatment only had limited effect on survival. The survival of weaver mutant mouse CGCs in medium containing 5 mM K+ ions was very low, only 20% of the plated cells surviving at 7 days after plating, as opposed to the 50% for non-weaver CGCs. Chronic high K+ treatment improved the relative survival of weaver mutant mouse CGCs 1.6 2.2-fold and that of non-weaver CGCs 1.2-1.4-fold; the same number of CGCs (about 20% of the plated cells) were rescued by high K+ in both types of culture. The findings indicate that, in culture weaver mutant mouse, CGCs have a normal response to membrane depolarization and that the normal function of the Girk2 potassium channel is not critical for the survival of differentiated CGCs.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}