 ,1,*
,1,*Effect of phosphorus (P) on nitrogen (N) uptake and utilization in peanut
YU Tian-Yi1,*, LI Xiao-Liang2,*, LU Ya3, SUN Xue-Wu1, ZHENG Yong-Mei1, WU Zheng-Feng1, SHEN Pu1, WANG Cai-Bin,1,*通讯作者:
收稿日期:2018-08-4接受日期:2019-01-12网络出版日期:2019-02-15
| 基金资助: |
Received:2018-08-4Accepted:2019-01-12Online:2019-02-15
| Fund supported: |
作者简介 About authors
E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (391KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
于天一, 李晓亮, 路亚, 孙学武, 郑永美, 吴正锋, 沈浦, 王才斌. 磷对花生氮素吸收和利用的影响[J]. 作物学报, 2019, 45(6): 912-921. doi:10.3724/SP.J.1006.2019.84107
YU Tian-Yi, LI Xiao-Liang, LU Ya, SUN Xue-Wu, ZHENG Yong-Mei, WU Zheng-Feng, SHEN Pu, WANG Cai-Bin.
花生是我国重要的经济和油料作物。氮是花生生长发育必需营养元素之一, 与花生植株生理代谢、产量及品质密切相关[1,2]。长期以来, 为追求高产, 花生生产中过于依赖化学氮肥, 氮肥投入量远大于花生需氮量, 造成根瘤固氮能力降低、植株徒长、早衰, 产量下降, 肥料利用率降低, 并带来一系列环境问题[3,4]。花生氮素营养来源除肥料氮之外, 还包括土壤氮和根瘤固氮[4,5]。因此, 通过提高花生3种氮源中的土壤氮和根瘤固氮的供应来降低花生对肥料氮的依赖是解决上述问题的重要措施之一。
磷与作物体内多种重要有机化合物(如脂肪、蛋白质、核蛋白)的合成有关, 是参与能量传递的高能磷酸化合物磷酸基团的组成成分[6,7]。豆科作物根瘤中的磷素积累约占整株磷积累的20%左右, 磷对根瘤的形成、氮的固定、氨的转化以及氨基酸的形成具有重要作用[8,9]。目前磷对豆科作物根瘤固氮及整株氮吸收、利用影响的研究主要集中在大豆、羽扇豆等作物。有人认为磷可以直接影响豆科作物结瘤及固氮[7,10]。缺磷会减少氧气供给, 增加氧化胁迫从而降低根瘤固氮效率[11,12,13]。磷缺乏引起根瘤中苹果酸合成, 但是苹果酸积累过度会降低天门冬氨酸转氨酶、合成酶、谷氨酰胺合成酶活性, 最终抑制根瘤固氮及整株氮积累[14,15]。另有人认为, 磷可以刺激寄主作物的生长, 间接影响根瘤菌的存活或生长及固氮功能的发挥等[16]。缺磷影响豆科作物细胞叶绿体中丙糖磷向液泡中转移, 降低RuBP羧化酶活性和叶片光合速率, 进而影响叶面积、叶片数量、分枝数, 限制碳素向根瘤中转移, 降低固氮酶活性及根瘤重量[17,18]。施磷可降低外源高氮对根瘤的抑制作用, 降低豆科作物对肥料氮和土壤氮依赖[19]。Anna等[20]在红色苜蓿上的研究表明, 高氮高磷处理的根瘤相关指标(如: 根瘤数量、根瘤干重、固氮酶等)比高氮低磷处理高6倍以上。严君等[21]报道, 我国东北地区, 大豆高产的氮磷肥推荐施肥量分别为50 kg hm-2和40 kg hm-2, 若要充分发挥根瘤菌的固氮能力, 应增加磷肥的施用比例, 此时的推荐量为纯氮 20 kg hm-2和40 kg hm-2。目前花生施磷效应相关研究多体现在产量、品质、根系形态及叶片生理特性等方面[22,23,24], 磷对花生不同氮源吸收、利用的影响鲜见报到。本文探讨了磷肥对不同花生品种氮素吸收、利用的影响, 以期为花生合理施磷提供依据。
1 材料与方法
1.1 试验地点及试验设计
试验于2015年和2016年在山东省花生研究所莱西试验基地进行(36°48′N, 120°29′E)。该地年均气温11.7℃, 年均降雨量635.8 mm (主要集中在5月至9月), 年日照时数2656.3 h。常年采用冬小麦-夏玉米-春花生两年三作种植模式。试验用直径38 cm、高度50 cm的PVC无底圆桶, 田间先起垄, 垄距80 cm, 垄面宽50 cm, 在垄上按圆桶尺寸将土挖出, 把圆桶埋入土内, 圆桶上边露出地表5 cm, 然后按0~25 cm和25~45 cm原土层顺序装入桶内, 两桶(边缘)相距20 cm, 垄上两桶之间种1穴(2粒种子)与桶内相同的品种作保护行。每桶播2粒种子, 齐苗后间苗, 每桶留1棵长势均匀一致的植株。试验设5个磷(P2O5)水平, 分别为0、45、90、135和180 kg hm-2, 根据桶的面积折算桶内施肥量, 即各处理每桶施磷量分别为0、0.51、1.02、1.53和2.04 g。供试花生品种为山东主推品种花育22号(大花生)和花育20号(小花生)。每处理氮、钾肥用量相同。其中氮(N)用量为0.85 g 桶-1(75 kg hm-2), 肥料用15N标记尿素(购买于上海化工研究院), 丰度为10.3%。钾(K2O)用量为2.04 g 桶-1(180 kg hm-2), 肥料用磷酸二氢钾和硫酸钾混合施用。各处理肥料用量见表1。播种前将肥料均匀施入0~20 cm土层中。重复6次。另以相同的处理方式种植不结瘤品种NN-1, 以计算土壤氮积累量和根瘤固氮积累量的比例。试验地为棕壤土, 容重1.54 g cm-3, pH 6.5, 含有机质1.24%、碱解氮56.8 mg kg-1、速效磷12.9 mg kg-1、速效钾79.3 mg kg-1。Table 1
表1
表1不同处理肥料用量
Table 1
| 肥料种类 Fertilizer type | P0 | P45 | P90 | P135 | P180 |
|---|---|---|---|---|---|
| 磷酸二氢钾 KH2PO4 | 0 | 0.98 | 1.96 | 2.94 | 3.92 |
| 硫酸钾 K2SO4 | 3.78 | 3.15 | 2.52 | 1.89 | 1.26 |
| 尿素 CO(NH2)2 | 1.85 | 1.85 | 1.85 | 1.85 | 1.85 |
新窗口打开|下载CSV
1.2 取样及测定方法
于花生成熟期, 将植株从桶内挖出, 并捡拾残根落叶, 用流水冲洗, 其下放一个100目的筛子以防脱落的根系及叶子被冲走。洗净后将花生分成3部分(荚果、果针和根茎叶)于烘箱105℃杀青1 h, 80℃烘干至恒重, 称重, 然后研磨、过筛。粉末经硫酸-双氧水消煮后定容、过滤, 用凯氏定氮仪(Kjeltec 2300)测定植株中的全氮含量, 用质谱仪(MAT-271)测定15N丰度。2016年, 取3桶花生植株将根系洗净, 吸干表面水分, 用医用解剖刀将根瘤取下, 统计数目, 称取鲜重。1.3 氮素利用相关指标的计算
采用Wang等[4]和郑永美等[25]的方法计算全氮积累量、肥料氮积累量及比例、土壤氮积累量及比例、根瘤固氮积累量及比例、氮肥利用率及氮素荚果生产效率等指标; 用赵亚丽等[26]的方法计算磷肥农学效率。1.4 统计分析
用Microsoft Excel 2007软件整理数据及作图, 用SPSS 13.0统计软件进行方差分析(LSD)及相关性分析。2 结果与分析
2.1 磷对3种氮源氮积累量的影响
花生植株全氮积累量随施磷量的增加而增加, 不同年份及品种表现基本一致。2016年花育22号各施磷处理的植株全氮积累量较对照增加16.11%~ 30.01%, 差异显著, P180显著高于P45和P90, P135与各施磷处理间差异不显著。从不同氮源氮积累量看, 施磷能够提高花生对肥料氮、土壤氮及根瘤固氮3种氮源的吸收。2016年花育22号各施磷处理的肥料氮、土壤氮及根瘤固氮积累量较对照分别增加11.72%~13.34%、11.95%~15.57%及19.23%~ 42.6%, 且随施磷量增加, 肥料氮和土壤氮积累量呈先增后减趋势或增幅降低, 各施磷处理间两指标差异不显著。但根瘤固氮积累量随施磷量增加呈增加趋势, 并未出现下降现象(图1)。说明施磷对根瘤固氮积累量的提高作用大于土壤氮和肥料氮, 且随施磷量增加这种现象更加明显。图1
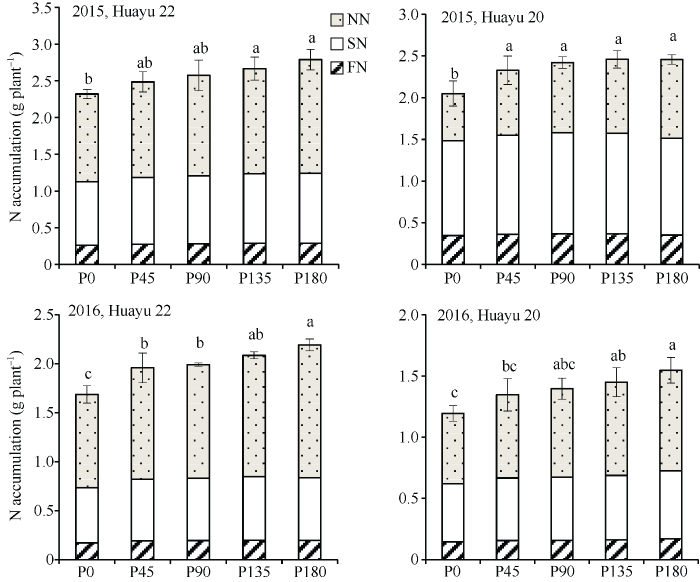 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1不同品种及磷水平条件下花生3种氮源氮积累量
标以不同小写字母的柱值于处理间0.05水平上差异显著。NN: 根瘤固氮; SN: 土壤氮; FN: 肥料氮; P0: 施磷量0 kg hm-2; P45: 施磷量45 kg hm-2; P90: 施磷量90 kg hm-2; P135: 施磷量135 kg hm-2; P180: 施磷量180 kg hm-2。
Fig. 1N accumulated amounts of three sources for peanut under different varieties and P rates
Bars superscripted by different letters are significantly different at the 0.05 probability level among treatments. NN: nitrogen fixed by root nodule; SN: nitrogen from soil; FN: nitrogen from fertilizer; P0: 0 kg hm-2 of P fertilizer; P45: 45 kg hm-2 of P fertilizer; P90: 90 kg hm-2 of P fertilizer; P135: 135 kg hm-2 of P fertilizer; P180: 180 kg hm-2 of P fertilizer.
2.2 磷对3种氮源氮积累比例的影响
由表2可知, 年份和品种对肥料氮、土壤氮和根瘤固氮3种氮源氮积累比例的影响均达到极显著水平。施磷对土壤氮和根瘤固氮积累比例的影响达极显著水平, 对肥料氮积累比例的影响不显著。年份与品种的交互效应对3种氮源氮积累比例的影响均达到极显著水平。施磷水平与年份和品种的两两交互效应对3种氮源氮积累比例的影响均未达到显著水平。除2015年花育20号之外, 3种氮源氮积累比例表现为根瘤固氮>土壤氮>肥料氮。随施磷量增加, 肥料氮和土壤氮积累比例呈降低趋势, 根瘤固氮积累比例呈增加趋势。2016年花育22号各施磷处理肥料氮和土壤氮积累比例较对照分别降低0.36~1.33和0.95~4.13个百分点, 根瘤固氮积累比例较对照增加1.78~5.40个百分点。其中P180处理肥料氮积累比例显著低于对照, 而根瘤固氮积累比例显著高于对照, 其他施磷处理与对照差异不显著; 各施磷处理的土壤氮积累比例与对照差异均未达显著水平。Table 2
表2
表2不同品种及磷水平条件下花生3种氮源氮积累比例
Table 2
| 年份 Year | 施磷量 P rate | 肥料氮 N from fertilizer | 土壤氮 N from soil | 根瘤固氮 N fixed by root nodule | |||||
|---|---|---|---|---|---|---|---|---|---|
| 花育22号 Huayu 22 | 花育20号 Huayu 20 | 花育22号 Huayu 22 | 花育20号 Huayu 20 | 花育22号 Huayu 22 | 花育20号 Huayu 20 | ||||
| 2015 | 0 | 11.34±0.51 a | 17.01±1.77 a | 37.19±1.81 a | 55.79±3.45 a | 51.53±0.42 b | 27.21±4.98 b | ||
| 45 | 11.17±1.29 a | 15.63±1.91 ab | 36.49±0.28 ab | 51.23±2.88 ab | 53.15±1.78 ab | 33.14±4.33 ab | |||
| 90 | 11.05±1.77 a | 15.26±0.75 ab | 35.93±0.04 ab | 50.05±2.45 ab | 53.20±1.25 ab | 34.70±3.19 ab | |||
| 135 | 10.89±1.21 a | 14.96±1.09 ab | 35.56±0.01 ab | 49.08±3.58 b | 53.76±1.11 ab | 35.95±4.67 a | |||
| 180 | 10.43±0.99 a | 14.40±0.56 b | 34.18±2.91 b | 47.22±1.85 b | 55.62±2.75 a | 38.39±2.42 a | |||
| 2016 | 0 | 10.23±0.45 a | 12.17±1.12 a | 33.32±1.51 a | 39.89±0.83 a | 56.59±2.33 b | 48.19±1.89 b | ||
| 45 | 9.87±1.21 ab | 11.68±2.08 a | 32.37±3.95 a | 38.31±2.05 a | 58.37±2.45 ab | 51.03±4.29 ab | |||
| 90 | 9.75±0.25 ab | 11.32±1.29 a | 31.98±0.82 a | 37.14±4.24 a | 58.27±0.52 ab | 51.93±4.34 ab | |||
| 135 | 9.52±0.30 ab | 11.15±1.51 a | 31.21±0.99 a | 36.59±4.96 a | 59.30±1.84 ab | 53.00±1.73 a | |||
| 180 | 8.90±0.42 b | 11.01±1.58 a | 29.19±1.37 a | 36.04±0.20 a | 61.99±3.10 a | 53.30±1.92 a | |||
| 显著性Significance (F-value) | |||||||||
| 年份Year (Y) | ** | ** | ** | ||||||
| 品种Variety (V) | ** | ** | ** | ||||||
| 施磷量P rate (P) | ns | ** | ** | ||||||
| 年份×品种 Y×V | ** | ** | ** | ||||||
| 年份×施磷量Y×P | ns | ns | ns | ||||||
| 品种×施磷量V×P | ns | ns | ns | ||||||
新窗口打开|下载CSV
2.3 磷对根瘤的影响
与根瘤固氮积累量和积累比例相似, 花生根瘤数量及鲜重随施磷量增加而增加。与对照相比, 花育22号和花育20号根瘤数量分别增加15.9%~ 44.1%和21.9%~36.1%, 其中花育22号各施磷处理均显著高于对照, 花育20号P135和P180处理显著高于对照。花育22号和花育20号的根瘤鲜重较对照分别增加5.3%~98.2%和29.9%~69.3%, 其中花育22号P90、P135和P180处理显著高于对照, 花育20号P135和P180显著高于对照(图2和图3)。图2
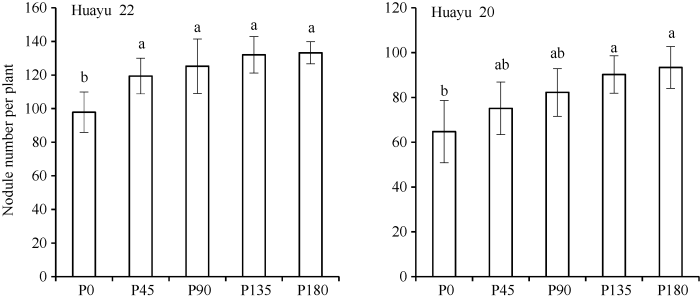 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同品种及磷水平条件下花生根瘤数量
标以不同小写字母的柱值于处理间0.05水平上差异显著。P0: 施磷量0 kg hm-2; P45: 施磷量45 kg hm-2; P90: 施磷量90 kg hm-2; P135: 施磷量135 kg hm-2; P180: 施磷量180 kg hm-2。
Fig. 2Nodule numbers of peanut under different varieties and P rates
Bars superscripted by different letters are significantly different at the 0.05 probability level among treatments. P0: 0 kg hm-2 of P fertilizer; P45: 45 kg hm-2 of P fertilizer; P90: 90 kg hm-2 of P fertilizer; P135: 135 kg hm-2 of P fertilizer; P180: 180 kg hm-2 of P fertilizer.
图3
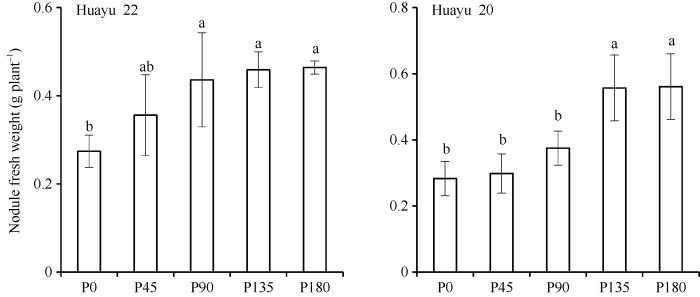 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同品种及磷水平条件下花生根瘤鲜重
标以不同小写字母的柱值于处理间0.05水平上差异显著。P0: 施磷量0 kg hm-2; P45: 施磷量45 kg hm-2; P90: 施磷量90 kg hm-2; P135: 施磷量135 kg hm-2; P180: 施磷量180 kg hm-2。
Fig. 3Nodule fresh weight of peanut under different varieties and P rates
Bars superscripted by different letters are significantly different at the 0.05 probability level among treatments. P0: 0 kg hm-2 of P fertilizer; P45: 45 kg hm-2 of P fertilizer; P90: 90 kg hm-2 of P fertilizer; P135: 135 kg hm-2 of P fertilizer; P180: 180 kg hm-2 of P fertilizer.
2.4 磷对氮、磷利用效率的影响
年份和施磷量单一效应对氮肥利用率、氮素荚果效率及磷肥农学效率的影响极显著。品种单一效应及品种和年份两因素交互效应对2个氮效率指标的影响均达极显著水平。花生氮肥利用率随施磷量增加而增加, 但施磷量超过90 kg hm-2或135 kg hm-2, 氮肥利用率呈下降或增幅降低趋势。如2016年花育22号各施磷处理的氮肥利用效率较对照增加2.43~3.16个百分点, 各施磷处理间差异不显著。随施磷量增加, 氮素荚果利用效率呈先增后减趋势, 两品种氮素荚果利用效率均在P45达到最高值, 其中2016年花育22号P45处理的氮素荚果利用效率较对照高0.2%, P90~P180处理较对照降低0.40%~ 2.09%, 其中P180处理与对照差异达显著水平。磷肥农学效率随施磷量增加而降低, 其中2016年花育22号P45处理较其他3个施磷处理分别显著高出45.21%、99.46%和184.29%。表明适量施磷能够提高氮肥利用率、氮素荚果效率及磷肥农学效率, 过量施磷三指标不升反降(表3)。Table 3
表3
表3不同品种及磷水平条件下氮肥利用率、氮肥荚果利用效率和磷肥农学效率
Table 3
| 年份 Year | 施磷量 P rate | 氮肥利用率 Nitrogen use efficiency (%) | 氮素荚果利用率 Nitrogen pod production efficiency (kg kg-1) | 磷肥农学效率 Agronomic efficiency of phosphorus fertilizer (kg kg-1) | |||||
|---|---|---|---|---|---|---|---|---|---|
| 花育22号 Huayu 22 | 花育20号 Huayu 20 | 花育22号 Huayu 22 | 花育20号 Huayu 20 | 花育22号 Huayu 22 | 花育20号 Huayu 20 | ||||
| 2015 | 0 | 30.93±0.83 b | 40.81±3.00 a | 18.64±0.61 ab | 19.06±0.35 a | — | — | ||
| 45 | 32.54±0.45 ab | 42.60±0.16 a | 19.08±0.68 a | 19.08±0.13 a | 8.00±0.85 a | 10.51±1.19 a | |||
| 90 | 33.17±2.66 ab | 43.23±1.24 a | 19.02±0.27 a | 18.72±0.14 ab | 5.57±0.74 b | 6.13±0.49 b | |||
| 135 | 34.01±2.01 a | 43.43±1.84 a | 18.56±0.37 ab | 18.70±0.23 ab | 4.01±0.45 bc | 4.53±0.92 bc | |||
| 180 | 34.10±0.58 a | 41.60±0.94 a | 17.77±0.27 b | 18.55±0.23 b | 3.08±0.61 c | 3.19±0.16 c | |||
| 2016 | 0 | 20.18±0.90 b | 17.05±0.96 b | 19.40±1.33 a | 21.34±0.95 a | — | — | ||
| 45 | 22.61±1.68 a | 18.32±1.76 ab | 19.60±0.37 a | 21.51±0.70 a | 7.42±0.68 a | 6.73±1.13 a | |||
| 90 | 22.83±0.46 a | 18.51±1.18 ab | 19.00±0.32 a | 21.40±0.60 a | 5.11±0.67 b | 3.36±1.08 b | |||
| 135 | 23.34±0.35 a | 18.90±1.53 ab | 18.36±0.30 ab | 21.29±0.18 a | 3.72±0.57 bc | 3.53±0.42 b | |||
| 180 | 22.94±0.67 a | 19.93±2.00 a | 17.31±0.53 b | 19.73±0.59 b | 2.61±0.27 c | 2.48±0.46 b | |||
| 显著性 Significance (F-value) | |||||||||
| 年份Year (Y) | ** | ** | ** | ||||||
| 品种Variety (V) | ** | ** | ns | ||||||
| 施磷量P rate (P) | ** | ** | ** | ||||||
| 年份×品种 Y×V | ** | ** | ns | ||||||
| 年份×施磷量Y×P | ns | ns | ns | ||||||
| 品种×施磷量V×P | ns | ns | ns | ||||||
新窗口打开|下载CSV
2.5 磷对生物产量和荚果产量的影响
年份、品种、施磷量三因素的单一效应及品种和年份两因素交互效应对荚果产量及生物产量的影响均达极显著水平。施磷量与年份、施磷量与品种的两两交互效应对荚果产量及生物产量的影响不显著。施磷增加了花生荚果产量, 但施磷量过高荚果产量不升反降, 除2015年花育22号P45处理外, 其他施磷处理的荚果产量均显著高于对照, 各施磷处理间差异不显著, 2016年花育22号各施磷处理的荚果产量较对照高11.49%~16.21%。生物产量随施磷量增加呈增加趋势, 2015年两品种P90~P180处理的生物产量均显著高于对照, 2016年花育22号的P135和P180两处理及花育20号的P180处理也均显著高于对照, 施磷处理之间差异不显著。说明施磷量45~90 kg hm-2时, 花生荚果产量为最佳, 过量施磷虽能提高生物产量但对荚果产量的影响较小, 2016年这种现象更为明显(表4)。Table 4
表4
表4不同品种及磷水平条件下花生荚果产量和生物产量
Table 4
| 年份 Year | 施磷量 P rate | 荚果产量 Pod yield | 生物产量 Biomass yield | |||
|---|---|---|---|---|---|---|
| 花育22号 Huayu 22 | 花育20号 Huayu 20 | 花育22号 Huayu 22 | 花育20号 Huayu 20 | |||
| 2015 | 0 | 43.25±1.62 b | 39.09±3.15 b | 84.73±2.68 b | 72.40±5.37 b | |
| 45 | 47.38±1.72 ab | 44.44±3.19 a | 92.83±3.55 ab | 80.60±6.53 ab | ||
| 90 | 48.98±4.45 a | 45.34±1.64 a | 97.13±6.90 a | 81.73±2.71 a | ||
| 135 | 49.44±2.35 a | 46.00±1.72 a | 97.63±5.63 a | 83.17±4.87 a | ||
| 180 | 49.57±3.23 a | 45.59±1.42 a | 97.80±4.94 a | 83.33±2.42 a | ||
| 2016 | 0 | 32.64±0.78 b | 25.46±1.23 b | 86.06±5.97 b | 57.45±3.59 b | |
| 45 | 36.39±2.50 a | 28.89±1.96 a | 94.56±9.24 ab | 62.14±3.64 ab | ||
| 90 | 37.81±0.84 a | 29.84±1.35 a | 93.74±3.88 ab | 65.66±5.48 ab | ||
| 135 | 38.29±1.07 a | 30.85±2.30 a | 103.33±1.37 a | 66.49±5.46 ab | ||
| 180 | 37.93±1.44 a | 30.52±2.72 a | 103.51±6.00 a | 67.75±2.85 a | ||
| 年份 Year | 施磷量 P rate | 荚果产量 Pod yield | 生物产量 Biomass yield | |||
| 花育22号 Huayu 22 | 花育20号 Huayu 20 | 花育22号 Huayu 22 | 花育20号 Huayu 20 | |||
| 显著性Significance (F-value) | ||||||
| 年份Year (Y) | ** | ** | ||||
| 品种Variety (V) | ** | ** | ||||
| 施磷量P rate (P) | ** | ** | ||||
| 年份×品种 Y×V | ** | ** | ||||
| 年份×施磷量 Y×P | ns | ns | ||||
| 品种×施磷量 V×P | ns | ns | ||||
新窗口打开|下载CSV
2.6 荚果产量与3种氮源氮积累量、积累比例及氮肥利用率的关系
由表5可知, 荚果产量与3种氮源及全氮积累量、氮肥利用率均呈极显著正相关, 与氮素荚果利用效率呈极显著负相关。根瘤固氮积累量与植株全氮积累量呈极显著正相关, 与植株土壤氮、肥料氮积累比例及氮素荚果利用效率呈极显著负相关。根瘤固氮积累比例与植株土壤氮和肥料氮积累量、积累比例及氮肥利用率呈极显著负相关。表明, 增加根瘤固氮积累量能够促进花生对氮素的吸收, 增加产量; 提高植株根瘤固氮比例能够降低花生对土壤氮和肥料氮的吸收、利用, 减少对土壤和肥料氮素的依赖。Table 5
表5
表5花生产量与氮素积累、利用相关性状的相关系数
Table 5
| X1 | X2 | X3 | X4 | X5 | X6 | X7 | X8 | X9 | |
|---|---|---|---|---|---|---|---|---|---|
| X2 | 0.988** | ||||||||
| X3 | 0.816** | 0.785** | |||||||
| X4 | 0.815** | 0.783** | 0.999** | ||||||
| X5 | 0.641** | 0.696** | 0.100 | 0.100 | |||||
| X6 | 0.236 | 0.170 | 0.743** | 0.745** | -0.583** | ||||
| X7 | 0.235 | 0.170 | 0.743** | 0.745** | -0.584** | 1.000** | |||
| X8 | -0.236 | -0.170 | -0.740** | -0.745** | 0.583** | -1.000** | -1.000** | ||
| X9 | 0.818** | 0.787** | 1.000** | 1.000** | 0.110 | 0.740** | 0.740** | -0.741** | |
| X10 | -0.733** | -0.825** | -0.531** | -0.522** | -0.712** | 0.053 | 0.054 | -0.053 | -0.534** |
新窗口打开|下载CSV
3 讨论
3.1 不同氮源氮积累量及根瘤特性
前人研究表明, 适当施磷能显著提高豆科作物根瘤固氮能力及产量, 磷过量或不足均不利于结瘤及产量形成。其原因是不适宜的磷水平会抑制光合作用和固氮过程, 磷代谢的恶化会损伤光合系统和 固氮过程的正常代谢[27]。Abbasi等[28]研究认为, 与不施磷相比, 施磷能提高花生根瘤固氮能力, 但施磷(P2O5) 100 kg hm-2处理, 大豆根瘤重量、数量及产量显著低于施磷50 kg hm-2处理。姚玉波等[19]通过砂培试验研究表明, 营养液磷浓度11 mg L-1已基本满足根瘤的需求, 再提高磷水平, 对根瘤固氮无明显促进作用。本试验的结论与前人不同, 施磷(P2O5) 0~180 kg hm-2范围内, 两花生品种根瘤数量、鲜重及根瘤固氮积累量呈持续增加趋势, 这可能与土壤肥力水平和不同作物根瘤对磷素需求阈值等因素有关。另外, 施磷增加了两品种肥料氮和土壤氮积累量, 但增幅远低于根瘤固氮积累量, 其原因可能是磷能够促进花生根系生长, 进而间接提高根系对两种氮源的吸收[29]。因此, 增施磷肥是促进花生根瘤生长、提高根瘤固氮能力及植株氮素吸收的重要措施。3.2 不同氮源氮积累比例
传统观点认为, 根瘤固氮可满足花生需氮量的70%~80%, 而土壤氮和肥料氮的作用仅占20%~ 30%。近年来, 随施肥水平及地力提升, 根瘤固氮的作用逐渐降低。吴正锋等[30]研究表明, 根瘤固氮仅能提供花生植株一半的需氮量。孙虎等[31]在盆栽条件下的研究认为, 花生植株全氮积累量中根瘤固氮占41.25%~56.85%, 土壤氮占41.85%~48.63%, 肥料氮占0~13.93%。本研究结果表明, 花生根瘤固氮积累比例达27.21%~61.99%, 土壤氮略低于根瘤固氮, 肥料氮则不足20%, 施肥处理、品种及年季间均有所差异。说明根瘤固氮在花生氮素代谢中发挥着重要作用, 而氮肥作用相对较小。前人研究表明[31], 随施氮量增加, 花生根瘤固氮量降低。为追求高产, 花生生产中往往投入大量氮肥, 相当大一部分通过淋洗、氨挥发及径流等途径损失, 仅有少数能被花生吸收利用, 不仅导致氮素资源浪费, 造成环境污染, 还极大地限制了根瘤固氮。因此, 适当控制氮肥投入, 保持合理的“氮磷比”并辅以其他农艺措施, 充分发挥根瘤菌固氮潜力, 是花生“减氮增效”的重要途径。3.3 氮、磷利用效率
氮肥利用效率及氮素荚果利用率是评价花生对氮肥(氮素)吸收、利用的重要指标。施磷肥是调控氮素利用的重要措施。华伟等[32]研究表明, 在施磷量0~330 kg hm-2范围内, 玉米植株氮积累量、氮肥利用效率及氮素利用效率(同本研究中氮素荚果利用率)均呈先增后减趋势。张萌等[33]在小麦上也得到了类似的结论。本研究得到了不同的结论, 即: 适量施磷能够提高这两个氮效率指标, 施磷量过高使其不升反降, 而根瘤鲜重、数量、根瘤固氮积累量及全氮积累量却持续增加, 主要是因为过量施磷导致花生与禾本科作物氮效率降低的机制不同。施磷过多往往导致禾本科作物体内游离氨基酸和酰胺含量增加, 阻碍蛋白质的合成, 进而降低植株氮积累量和氮利用效率; 而本研究中过量施磷促进了花生根瘤固氮能力, 反而提高了植株氮素积累, 但有可能增强植株的呼吸强度, 不利于植株生长发育及产量形成, 进而造成氮素奢侈吸收、氮效率下降。磷肥农学效率表示单位施磷量所增加的作物产量。本研究表明, 施磷量在45~180 kg hm-2范围内, 磷肥农学效率随施磷量增加显著降低, 这与侯云鹏等[34]在水稻上得到的结论基本一致。而李月梅[35]研究表明, 施磷量在37.5~112.5 kg hm-2范围内, 油菜磷肥农学效率无显著差异, 这与供试土壤磷素水平及试验条件等因素有关。综合来看, 施磷量45~90 kg hm-2是兼顾花生产量和氮、磷效率的适宜施磷量。另外, 在现有的花生种质中选择或培育产量及肥料利用率双高的品种也是实现花生节本增效的有效途径之一。
3.4 氮素吸收、利用与荚果产量的关系
郑永美等[25]研究表明, 花生根瘤固氮积累量与荚果产量、全氮积累量、肥料氮积累量、土壤氮积累量及氮肥利用率呈显著或极显著正相关, 与肥料氮和土壤氮供氮比例呈极显著负相关。根瘤固氮积累比例与荚果产量、全氮积累量呈极显著正相关, 与肥料氮和土壤氮积累量呈极显著负相关。而本研究结果与之略有不同, 即根瘤固氮积累量与植株全氮积累量呈极显著正相关, 与土壤氮、肥料氮积累比例及氮素荚果利用效率呈极显著负相关。根瘤固氮积累比例与土壤氮和肥料氮积累量、积累比例及氮肥利用率呈极显著负相关。因此, 生产上充分发挥花生根瘤固氮潜力, 提高根瘤固氮积累量或积累比例, 是促进花生植株氮素吸收及减少氮肥依赖的重要途径。4 结论
施磷可提高花生肥料氮、土壤氮及根瘤固氮3种氮源氮积累量, 且根瘤固氮积累量的增幅大于土壤氮和肥料氮; 可降低土壤氮、肥料氮积累比例, 增加根瘤固氮积累比例。适量施磷可提高花生氮肥利用率、氮素荚果利用率及荚果产量, 两品种适宜的施磷量为45~90 kg hm-2。磷肥农学效率随施磷量增加而降低, 过量施磷不利于产量及氮、磷效率的进一步提高。增加根瘤固氮积累量能够促进花生对氮素的吸收, 增加产量; 提高植株根瘤固氮积累比例能够降低花生对土壤氮和肥料氮的吸收、利用, 减少对土壤和肥料氮的依赖。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/S2095-3119(15)61079-6URL [本文引用: 3]

Rational application of different forms of nitrogen (N) fertilizer for peanut (Arachis hypogaeaL.) requires tracking the N supplied sources which are commonly not available in the differences among the three sources: root nodule, soil and fertilizer. In this study, two kinds of peanut plants (nodulated variety (Huayu 22) and non-nodulated variety (NN-1)) were choosed and four kinds of N fertilizers: urea-N (CONH2-N), ammonium-N (NH4+-N), nitrate-N (NO361-N) and NH4++NO361-N labeled by15N isotope were applied in the field barrel experiment in Chengyang Experimental Station, Shandong Province, China, to determine the N supplied sources and N use efficiency over peanut growing stages. The results showed that intensities and amounts of N supply from the three sources were all higher at middle growing stages (pegging phase and podding phase). The accumulated amounts of N supply from root nodule, soil and fertilizer over the growing stages were 8.3, 5.3 and 3.8 g m612in CONH2-N treatment, which are all significantly higher than in the other three treatments. At seedling phase, soil supplied the most N for peanut growth, then root nodule controlled the N supply at pegging phase and podding phase, but soil mainly provided N again at the last stage (pod filling phase). For the whole growing stages, root nodule supplied the most N (47.8 and 43.0%) in CONH2-N and NH4+-N treatments, whereas soil supplied the most N (41.7 and 40.9%) in NH4++ NO361-N and NO361-N treatments. The N use efficiency was higher at pegging phase and podding phase, while accumulated N use efficiency over the growing stages was higher in CONH2-N treatment (42.2%) than in other three treatments (30.4% in NH4+-N treatment, 29.4% in NO361-N treatment, 29.4% in NH4++NO361-N treatment). In peanut growing field, application of CONH2-N is a better way to increase the supply of N from root nodule and improve the N use efficiency.
DOI:10.1016/j.fcr.2013.11.004URL [本文引用: 1]
Worldwide, legumes are grown on approximately 250Mha and fix about 90Tg of N2 per year. Plants involved in biological nitrogen fixation (BNF) are particularly sensitive to deficiencies of phosphorus (P), potassium (K) and sulphur (S). These nutrients can affect BNF directly; this is modulating growth of rhizobia, nodule formation and functioning, or indirectly by affecting the growth of the host plant. However, several process and mechanisms remain unclear. We compiled a data set (63 studies) on the effects of P, K, and S deficiency on shoot mass, nodule mass and number, nitrogenase activity (estimated by the acetylene reduction activity test, ARA) and the concentration of N, P, K and S in shoots and nodules. Our aims were (1) to compare the relative sensitivity of these traits to nutrient deficiency and (2) to probe for nutrient-specific patterns in trait responses. Our quantitative analysis confirms that nodule growth and number are more sensitive than shoot mass in response to deficiency of P, K and S. In addition, nodule activity decreases more than both shoot and nodule mass, which indicates a reduction in nodule productivity; this is likely related with direct effects of these nutrients on physiological and metabolic processes of nodules. The conserved shoot N concentration, in comparison to concentration of P, K and S indicates a relatively greater accumulation of N that matches the proposed N-feedback mechanism down-regulating BNF in nutrient-deficient systems. Despite some nutrient-specific differences, i.e. smaller nodules and higher N/K ratio with shortage of P and K, respectively, the patterns of growth, nodule activity and nutrient concentration were similar for all three nutrients P, K and S. This indicates that a unique mechanism could be depressing BNF (N-feedback) in conjunction with direct effects of the nutrients on nodule activity. Scarcity of data related to N, K and S concentration in nodules is a major constrain for deep analysis of the deficiencies of the nutrients. Critical concentrations of P, K and S in plant and nodule tissues are also a major gap. Models are needed that integrate the direct effect of the nutrients on nodule growth and activity with the N-feedback mechanism.
[本文引用: 1]
DOI:10.1016/j.plantsci.2015.06.018URLPMID:26398789 [本文引用: 2]
Legumes have a significant role in effective management of fertilizers and improving soil health in sustainable agriculture. Because of the high phosphorus (P) requirements of N2-fixing nodule, P deficiency represents an important constraint for legume crop production, especially in tropical marginal countries. P deficiency is an important constraint for legume crop production, especially in poor soils present in many tropical degraded areas. Unlike nitrogen, mineral P sources are nonrenewable, and high-grade rock phosphates are expected to be depleted in the near future. Accordingly, developing legume cultivars with effective N2 fixation under P-limited conditions could have a profound significance for improving agricultural sustainability. Legumes have evolved strategies at both morphological and physiological levels to adapt to P deficiency. Molecular mechanisms underlying the adaptive strategies to P deficiency have been elucidated in legumes. These include maintenance of the P-homeostasis in nodules as a main adaptive strategy for rhizobia-egume symbiosis under P deficiency. The stabilization of P levels in the symbiotic tissues can be achieved through several mechanisms, including elevated P allocation to nodules, formation of a strong P sink in nodules, direct P acquisition via nodule surface and P remobilization from organic-P containing substances. The detailed biochemical, physiological and molecular understanding will be essential to the advancement of genetic and molecular approaches for enhancement of legume adaptation to P deficiency. In this review, we evaluate recent progress made to gain further and deeper insights into the physiological, biochemical and molecular reprogramming that legumes use to maintain P-homeostasis in nodules during P scarcity.
DOI:10.1111/j.1399-3054.1997.tb05406.xURL [本文引用: 1]
The limitation of symbiotic nitrogen fixation due to P deficiency restricts the development of a sustainable agriculture, particularly in Mediterrancan and tropical soils. Common bean genotypes, APN18, BAT271, PVA846, POT51, G2633 and G12168, were grown in an aerated N-free nutrient solution at low (72 mol plant -1 week -1 ) and control P supplies (360 渭mol plant -1 week -1 ). Nitrogenase activity was estimated by in situ measurements of acetylene reduction activity (ARA) in a flow-through system. During the assays, maximum values of ARA (peak ARA) were reached between 20 and 30 min after exposure to C 2 H 2 , depending on P treatment and growth stage. Thereafter, a decline in C 2 H 4 evolution was observed. This decline was most pronounced in low-P plants and there was a significant genotypic effect. ARA per plant was decreased by P deficiency, mostly because nodulation was delayed and the number and mass of nodules were reduced. The ARA decrease during pod filling was also activated by P deficiency. ARA per g dry weight nodule was increased by P deficiency in G2633 and G12168, unchanged in APN18, BAT271 and POT51 and decreased in PVA846. Except for the climbing type IV G2633, total N at harvest for all P treatments was correlated with the cumulative value of peak ARA and with peak ARA at early pod-filling which was the highest peak ARA throughout the growth cycle of type III bushy genotypes. We conclude that if phenology and growth habit are carefully considered, peak ARA is a reliable screen of genotypes for N 2 fixation tolerance to P deficiency. Selection of lines with early nodulation under P deficiency is also advisable, and the effect of P deficiency on the nodule functioning needs to be considered.
[本文引用: 1]
DOI:10.1146/annurev-arplant-050213-035949URLPMID:24579991 [本文引用: 1]
Abstract Phosphorus is an essential nutrient that is required for all major developmental processes and reproduction in plants. It is also a major constituent of the fertilizers required to sustain high-yield agriculture. Levels of phosphate--the only form of phosphorus that can be assimilated by plants--are suboptimal in most natural and agricultural ecosystems, and when phosphate is applied as fertilizer in soils, it is rapidly immobilized owing to fixation and microbial activity. Thus, cultivated plants use only approximately 20-30% of the applied phosphate, and the rest is lost, eventually causing water eutrophication. Recent advances in the understanding of mechanisms by which wild and cultivated species adapt to low-phosphate stress and the implementation of alternative bacterial pathways for phosphorus metabolism have started to allow the design of more effective breeding and genetic engineering strategies to produce highly phosphate-efficient crops, optimize fertilizer use, and reach agricultural sustainability with a lower environmental cost. In this review, we outline the current advances in research on the complex network of plant responses to low-phosphorus stress and discuss some strategies used to manipulate genes involved in phosphate uptake, remobilization, and metabolism to develop low-phosphate-tolerant crops, which could help in designing more efficient crops.
DOI:10.1093/aob/mcf260URLPMID:12451030 [本文引用: 1]
The effects of P deficiency on growth, N60-fixation and photosynthesis in white clover (Trifolium repens L.) plants were investigated using three contrasting relative addition rates of P, or following abrupt withdrawal of the P supply. Responses to a constant below-optimum P supply rate consisted of a decline in N60-fixation per unit root weight and a small reduction in the efficiency with which electrons were allocated to the reduction of N60 in nodules. Abrupt removal of P arrested nodule growth and caused a substantial decline in nitrogenase activity per unit root weight, but not per unit nodule mass. Similarly, the rate of photosynthesis per unit leaf area was unaffected by abrupt P removal, whereas CO60 acquisition for the plant as a whole decreased due to a decline in total leaf area, leaf area per unit leaf weight and utilization of incoming radiation. These changes followed the decline in tissue P concentrations. The ratio between CO60-fixation and N60-fixation was maintained under short-term P deprivation but increased under long-term low P supply, indicating a regulatory inhibition of nodule activity following morphological and growth adjustments. It is concluded that N60-fixation did not limit the growth of clover plants experiencing P deficiency. A low P status induced changes in the relative growth of roots, nodules and shoots rather than changes in N and/or C uptake rates per unit mass or area of these organs.
.
DOI:10.1111/nph.12714URLPMID:24571730 [本文引用: 1]
The reactive oxygen species (ROS) generated by respiratory burst oxidative homologs (Rbohs) are involved in numerous plant cell signaling processes, and have critical roles in the symbiosis between legumes and nitrogen-fixing bacteria. Previously, down-regulation of RbohB in Phaseolus vulgaris was shown to suppress ROS production and abolish Rhizobium infection thread (IT) progression, but also to enhance arbuscular mycorrhizal fungal (AMF) colonization. Thus, Rbohs function both as positive and negative regulators. Here, we assessed the effect of enhancing ROS concentrations, by overexpressing PvRbohB, on the P. vulgaris090009rhizobia and P. vulgaris090009AMF symbioses.We estimated superoxide concentrations in hairy roots overexpressing PvRbohB, determined the status of early and late events of both Rhizobium and AMF interactions in symbiont-inoculated roots, and analyzed the nodule ultrastructure of transgenic plants overexpressing PvRbohB.Overexpression of PvRbohB significantly enhanced ROS production, the formation of ITs, nodule biomass, and nitrogen-fixing activity, and increased the density of symbiosomes in nodules, and the density and size of bacteroides in symbiosomes. Furthermore, PvCAT, early nodulin, PvSS1, and PvGOGAT transcript abundances were elevated in these nodules. By contrast, mycorrhizal colonization was reduced in roots that overexpressed RbohB.Overexpression of PvRbohB augmented nodule efficiency by enhancing nitrogen fixation and delaying nodule senescence, but impaired AMF colonization.
URL [本文引用: 1]
The presence of nitrogen inhibits legume nodule formation, but the mechanism of this inhibition is poorly understood. We found that 2.5 mM nitrate and above significantly inhibited nodule initiation but not root hair curling inMedicago trunatula. We analyzed protein abundance inM. truncatularoots after treatment with either 0 or 2.5 mM nitrate in the presence or absence of its symbiontSinorhizobium melilotiafter 1, 2 and 5 days following inoculation. Two-dimensional gel electrophoresis combined with mass spectrometry was used to identify 106 differentially accumulated proteins responding to nitrate addition, inoculation or time point. While flavonoid-related proteins were less abundant in the presence of nitrate, addition of Nod gene-inducing flavonoids to theSinorhizobiumculture did not rescue nodulation. Accumulation of auxin in response to rhizobia, which is also controlled by flavonoids, still occurred in the presence of nitrate, but did not localize to a nodule initiation site. Several of the changes included defense- and redox-related proteins, and visualization of reactive oxygen species indicated that their induction in root hairs followingSinorhizobiuminoculation was inhibited by nitrate. In summary, the presence of nitrate appears to inhibit nodulation via multiple pathways, including changes to flavonoid metabolism, defense responses and redox changes.
DOI:10.1111/j.1469-8137.2007.02305.xURL [本文引用: 1]
090004 Nodulated lupins ( Lupinus angustifolius cv. Wonga) were hydroponically grown under conditions of low phosphate (LP) or adequate phosphate (HP) to assess the effect of phospho enol pyruvate carboxylase (PEPC)-derived organic acids on nitrogen assimilation in LP nodules. 090004 LP conditions are linked to altered organic acid metabolism, by the engagement of PEP metabolism via PEPC. In LP nodules, the enhanced organic acid synthesis may reduce the available organic carbon for nitrogen assimilation. The diversion of carbon between the organic acid- and amino acid pools was assessed through key nodular enzymes and 14 CO 2 metabolism. 090004 Under LP conditions, increased rates of organic acid synthesis via PEPC and malate dehydrogenase (MDH), coincided with reduced nitrogen assimilation via aspartate aminotransferase (AAT), aspartate synthetase (AS) and glutamine synthetase (GS)/glutamate synthase (GOGAT) activities. There was a preferential metabolism of nodular 14 CO 2 into organic acids and particularly into malate. High malate levels were associated with reduced N 2 fixation and synthesis of amino acids. 090004 These results indicate that phosphorus deficiency can enhance malate synthesis in nodules, but that excessive malate accumulation may inhibit N 2 fixation and nitrogen assimilation.
[本文引用: 1]
[本文引用: 1]
DOI:10.1074/jbc.271.13.7609URL [本文引用: 1]
Phosphorous (P) fertilization is the major mineral nutrient yield determinant among legume crops. However, legume crops vary widely in the ability to take up and use P during deficiency. The aim here was to compare P uptake and translocation, biological nitrogen fixing ability and photosynthetic rate among mashbean (Vigna aconitifolia cv. 'Mash-88'), mungbean (Vigna radiata cv. 'Moong-6601') and soybean (Glycine max L. cv. 'Tamahomare') during deficiency in hydroponics. Two treatments, the withdrawalof P from the solution (P-deprivation) and continued P at 160 mu M (P sufficient) were effected at the pod initiation stage. Plants were grown for 20 days. Short-term labeling with ~(32)P showed the uptake and distribution of P into plant parts. Withdrawal of P from the solution reduced biomass, photosynthetic activity, and nitrogen fixing ability in mungbean, and mashbean more than in soybean. P deprivation decreased P accumulation more than N accumulation. The decrease was more severe in mungbean andmashbean than soybean. More P was translocated and distributed into leaves in soybean than in mungbean and mashbean. Leaf P amount was more correlated to leaf area than to photosynthetic rate per unit leaf area among all three legume species. The results indicate that selection for increased efficiency of P utilization and leaf area may be used to improve leguminous crops.
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s10705-005-0075-3URL [本文引用: 1]
Phosphorus and calcium are the major nutrients limiting groundnut production. The objectives were to determine (a) optimum application levels of P and Ca, and (b) compare the effectiveness of calcitic lime (40% Ca, 4.5% Mg) and gypsum (22% Ca, 17% S) as sources of Ca for groundnut grown on sandy soils. Field experiments were established in smallholder farming areas using four levels of P (0, 8.5, 17 and 3402kg02ha 611 ) combined factorially with calcitic lime (0, 200, 400 and 80002kg02ha 611 ) to give 16 treatments. Similar levels of P were combined factorially with gypsum (0, 100, 200 and 40002kg02ha 611 ) to give sixteen treatments. Experiments were laid in a randomized complete block design with three replications. Phosphorus had a significant effect on groundnut yield at the majority of the experimental sites. Application of P at 8.502kg02ha 611 gave the optimum groundnut yield response. The optimum application rates for calcitic lime and gypsum were 200 and 10002kg02ha 611 . Gypsum and calcitic lime were not significantly different as sources of Ca for groundnut. Soil chemical properties were significantly improved following application of P and Ca sources.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1626/pps.11.368URL [本文引用: 1]
Two separate experiments (pot and field) were conducted to examine the response of soybean to Bradyrhizobium japonicum and phosphorus (P) fertilization. Different treatments were (i) Rhizobium strains (0, S377, S379, and the mixture of S377+S379 i.e. S0, S1, S2, S3); (ii) phosphorus (field only, 0, 50, 100 kg ha-1 i.e. T0, T1, T2) and (iii) two soils (pot only) i.e. autoclaved (A1) and non-auto...
DOI:10.1364/AO.49.001586URLPMID:2806177 [本文引用: 1]
Abstract BACKGROUND AND AIMS: White lupin is highly adapted to growth in a low-P environment. The objective of the present study was to evaluate whether white lupin grown under P-stress has adaptations in nodulation and N2 fixation that facilitate continued functioning. METHODS: Nodulated plants were grown in silica sand supplied with N-free nutrient solution containing 0 to 0.5 mm P. At 21 and 37 d after inoculation (DAI) growth, nodulation, P and N concentration, N2 fixation (15N2 uptake and H2 evolution), root/nodule net CO2 evolution and CO2 fixation (14CO2 uptake) were measured. Furthermore, at 21 DAI in-vitro activities and transcript abundance of key enzymes of the C and N metabolism in nodules were determined. Moreover, nodulation in cluster root zones was evaluated. KEY RESULTS: Treatment without P led to a lower P concentration in shoots, roots, and nodules. In both treatments, with or without P, the P concentration in nodules was greater than that in the other organs. At 21 DAI nitrogen fixation rates did not differ between treatments and the plants displayed no symptoms of P or N deficiency on their shoots. Although nodule number at 21 DAI increased in response to P-deficiency, total nodule mass remained constant. Increased nodule number in P-deficient plants was associated with cluster root formation. A higher root/nodule CO2 fixation in the treatment without P led to a lower net CO2 release per unit fixed N, although the total CO2 released per unit fixed N was higher in the treatment without P. The higher CO2 fixation was correlated with increased transcript abundance and enzyme activities of phosphoenolpyruvate carboxylase and malate dehydrogenase in nodules. Between 21 and 37 DAI, shoots of plants grown without P developed symptoms of N- and P-deficiency. By 37 DAI the P concentration had decreased in all organs of the plants treated with no P. At 37 DAI, nitrogen fixation in the treatment without P had almost ceased. CONCLUSIONS: Enhanced nodulation in cluster root zones and increased potential for organic acid production in root nodules appear to contribute to white lupin's resilience to P-deficiency.
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}